葡萄品种资源遗传关系的ISSR分析
2014-12-24赖呈纯范丽华黄贤贵甘煌灿谢鸿根余亚白
赖呈纯,范丽华,黄贤贵,甘煌灿,谢鸿根,余亚白
(福建省农业科学院农业工程技术研究所,福建福州350003)
葡萄是最古老的植物之一,经历数百万年的进化,已发展成多个品种,并在人类悠久的栽培过程中,品种不断增加,进而形成了诸多各具地区特色的品种群。目前已知全世界葡萄属植物有70余种,我国是东亚种群的集中分布区,分布约38个种[1].葡萄属植物起源广泛,在人类的长期引种和选育中,不同地域间品种交流频繁,而且葡萄分类标准不统一,导致同名异物或同物异名现象十分普遍,同时葡萄种间杂交容易产生一些中间型和过渡型杂种,也影响分类鉴定和选育种等研究[2,3].因此,开展葡萄种质资源尤其是栽培种的亲缘关系及遗传多样性研究,对葡萄选育种和种质资源的保护利用具有重要意义[4].传统的葡萄资源分类基本以形态学、生理学、农艺性状、同工酶标记等为主[5-13].由于人类定向选育的需求,许多葡萄育种亲本越来越集中在少数优良品种(系)上,以致新品种在植物学性状上较相似,使得传统的形态学标记难以区分相似性高、亲缘关系较近的品种(系)[14].
随着生物技术的发展,从基因水平上进行研究更能真实地反应物种亲缘关系[15].与传统分类鉴定方法相比,DNA分子标记技术以其快速、高效、重复性高等特点,在许多果树分类、品种鉴定、系谱分析、遗传图谱构建、基因标记、分子辅助选择育种等多方面得到广泛应用[16,17].其中,ISSR是一种基于微卫星序列的分子标记,其与RAPD相比更可靠、重复性和多态性更高,并具备RAPD的简便和易操作性;其与RFLP、AFLP相比更快捷、稳定、成本较低、DNA用量小、安全性较高;与SSR相比,不需要预先获知序列信息,程序简化因而成本更低[18].因此,ISSR分子标记在果树遗传资源鉴定中应用广泛.近年来,采用RAPD、AFLP、SSR、ISSR等分子标记技术对葡萄的遗传多样性进行分析,能够解决传统分类方法不能解决的一些问题[19-31],并为葡萄种质资源的高效利用提供依据.目前,国内基于DNA分子标记技术的葡萄种质资源遗传多样性研究中,由于样本数量较少或只进行系谱研究,未能充分反映我国葡萄种质资源尤其是栽培种的遗传多样性情况.因此,需要开展较大规模的葡萄种质资源遗传多样性研究,才能对其进行有效利用.本研究共收集95份葡萄品种(系),利用优化后的ISSR体系[32]对其进行分析,从分子水平上探讨其遗传多样性及亲缘关系,该研究结果不仅是传统葡萄分类的补充,而且可为葡萄品种鉴定、杂交亲本选配和葡萄种质资源利用提供依据.
1 材料与方法
1.1 材料
95份葡萄品种(系)为各地引入福建栽培或作为种质保存的品种资源以及福建当地资源(表1)[1,33].所用引物为British Columbia大学公布的ISSR引物序列,由上海生工生物工程有限公司合成.

表1 供试材料1)Table 1 Meterials used in the study

续表1
1.2 方法
1.2.1 基因组DNA提取 取每份供试材料的幼嫩叶片,用改良CTAB[34]法提取基因组DNA,1.0%琼脂糖凝胶电泳检测质量,紫外分光光度计检测DNA浓度,用超纯水稀释至40 ng·μL-1,于-20℃保存备用.
1.2.2 ISSR-PCR扩增 根据赖呈纯等[32]优化后的葡萄ISSR-PCR反应体系进行.扩增程序为:94℃预变性5 min,94℃变性1 min,50-52℃(不同引物退火温度有差异)退火1 min,72℃延伸90 s,设40次循环,最后72℃延伸10 min,10℃保存.PCR产物用2.0%琼脂糖凝胶于4 V·cm-1电场下电泳60 min,电泳结束后琼脂糖凝胶经EB(0.5 μg·L-1)染色,并在凝胶成像系统中拍照.
1.2.3 数据分析 根据PCR扩增产物的电泳结果,采用人工读带方式,在凝胶相同迁移位置上的DNA条带,有带记为1,无带记为0,同时剔除模糊不清的条带,由此生成由0和1组成的原始矩阵.采用POPGENE Version 1.31软件计算多态位点百分率、Nei's遗传多样性指数、Shannon信息多样性指数、观察等位基因数和有效等位基因数等参数.NTSYSpc V2.10e软件计算遗传相似系数及遗传距离,并依据遗传相似系数利用UPGMA法进行聚类分析,绘制树状聚类图.
2 结果与分析
2.1 扩增产物多态性分析
根据British Columbia大学公布的100条ISSR引物序列,以001号材料对引物进行筛选,最终选出12条引物(表2),12条引物扩增的谱带清晰,且多态性丰富(图1).12条引物共扩增出160条带,其中多态性条带147条,平均多态性位点为91.88%.单条引物扩增的条带为11-18条,平均13.33条.扩增产物大小为300-3000 bp,以500-2000 bp的扩增片段居多.扩增条带数最多的引物为UBC810(18条),扩增条带数最少的引物为UBC811、UBC823、UBC834、UBC835、UBC873,扩增出的条带均为11条.多态性最高的为 UBC811、UBC834、UBC840、UBC845、和 UBC873,达 100%,最低的为 UBC835,为 63.64%.因此,ISSR 检测葡萄遗传多样性的效率较高,也表明葡萄的遗传多样性丰富.

表2 不同引物的碱基序列及扩增结果Table 2 Base sequences and amplification results of different primers
2.2 葡萄遗传多样性分析
通过对95个葡萄品种(系)资源群体遗传参数进行分析,结果表明,95个葡萄品种(品系)平均观察等位基因数为1.8750,平均有效等位基因数为1.4326,平均Nei's基因多样性指数为0.2598,平均Shannon信息指数为0.3979.就各位点而言,有效等位基因数最大值为1.9996,最小值为1.0000;Nei’'s基因多样性指数0-0.4999;Shannon信息指数0-0.6930.这些分析数据进一步表明,95个葡萄品种(系)资源间存在丰富的遗传多样性.
根据传统的种群分类可将95份葡萄品种(系)资源分成5个种群(表3).平均Nei's基因多样性指数最高的是欧美杂种(0.2338),最低是美洲种(0.1139);平均Shannon信息指数最高的是欧亚种(0.3511),最低是美洲种(0.1663).不同种群的遗传多态性为27.50%-71.88%.这说明在葡萄种群内遗传多样性也极丰富.
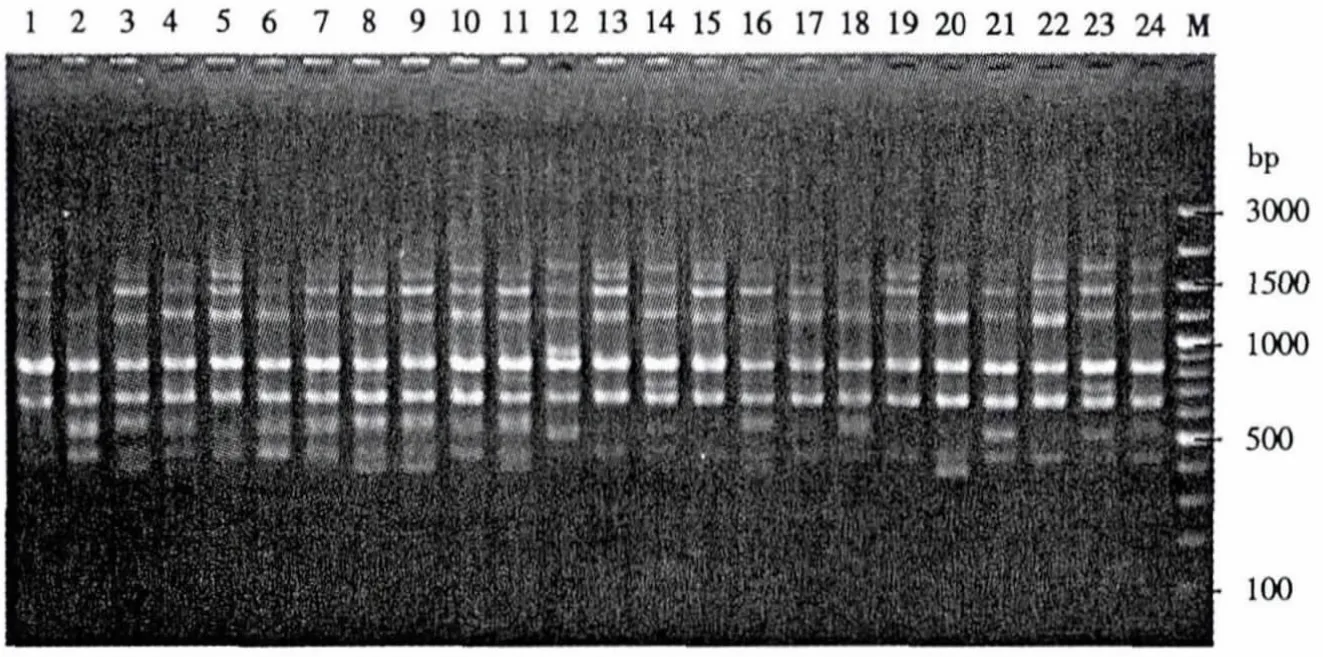
图1 引物UBC815扩增的带型Fig.1 Amplification profiles obtained with UBC primer 815

表3 95份葡萄样品遗传多样性的ISSR分析Table 3 ISSR analysis of genetic diversity of 95 grape samples
2.3 聚类分析
95份葡萄品种(系)资源的遗传相似系数0.55000-0.99375,平均遗传相似系数0.72171.其中,“野生171”和“野生172”的遗传相似系数最大(0.99375),“刺葡萄187”和“户太8号”遗传相似系数最小(0.55000).利用ISSR标记数据计算供试材料间的遗传相似性系数矩阵,采用UPGMA法构建95份葡萄品种(系)资源间的遗传关系聚类图(图2).从图中可知,当遗传相似系数为0.64时,可将东亚种群的野生种与东亚种群的刺葡萄以及其他种群的葡萄品种(系)资源区分开;当遗传相似系数为0.66时,可将东亚种群与其他种群相区分;当遗传相似系数为0.74时,可以将欧美杂种和欧亚种区分,其中欧亚种在该位置只分成1组,而欧美杂种在该位置分成4个小组.3个欧山杂种分处于不同种群,其中“桂葡1号”归类到欧亚种群中,而“北醇”则归类至欧美杂种种群内.2个美洲种内杂种在欧美杂种种群内.从聚类的结果看,3个在福建建阳当地种植年代久远的葡萄资源应为欧美杂种;而2个无相关记录表明种群归属的葡萄,“美秋”应为欧美杂种,“婴儿子”应为欧亚种.在聚类图中有4个葡萄品种与传统的分类相比较有一定的出入,“金手指”[1,33]在相关品种介绍时属于欧美杂种,而却归类到欧亚种,“矢富罗莎”、“六月紫”、“霞多丽”等[1,33]3个品种介绍为欧亚种,但聚类时却划入欧美杂种,这种情况可能是由于品种的混杂而引起,也可能与其多代回交亲本的选择有关,有待进一步研究.
3 讨论
葡萄种质资源的鉴定和评价对于育种亲本选配、丰富遗传基础以及种质资源科学利用等方面具有重要作用.研究人员利用传统的形态学、生理学、农艺性状、同工酶标记等对葡萄分类进行了大量研究[5-9,11-13],为我国葡萄品种资源鉴定起到关键作用,但基因型与环境互作使这些鉴定方法受到限制,而且人类定向选育使得利用传统方法鉴别相似性较高或亲缘关系较近的品种(系)时存在较大困难.DNA分子标记技术具备高效、准确、不受环境及主观因素影响等特点,在植物种质资源的鉴定与评价中得到了广泛的应用.本研究在筛选出12条ISSR引物的基础上,对95份葡萄种质进行遗传多样性分析,单条引物扩增的条带为11-18条,共扩增出160条带,其中多态性条带147条,平均多态性位点百分率为91.88%,表明葡萄在长期进化过程中,其基因组DNA发生了丰富变异,同时也说明ISSR分子标记可以较好地用于葡萄多样性分析.95份葡萄种质间Nei's遗传多样性指数0-0.4999,平均0.2598;遗传相似系数0.55625-0.99375,平均0.72182,这与张永辉等[35]对 81 份葡萄种质研究中的遗传相似系数(0.57-0.99)相近.
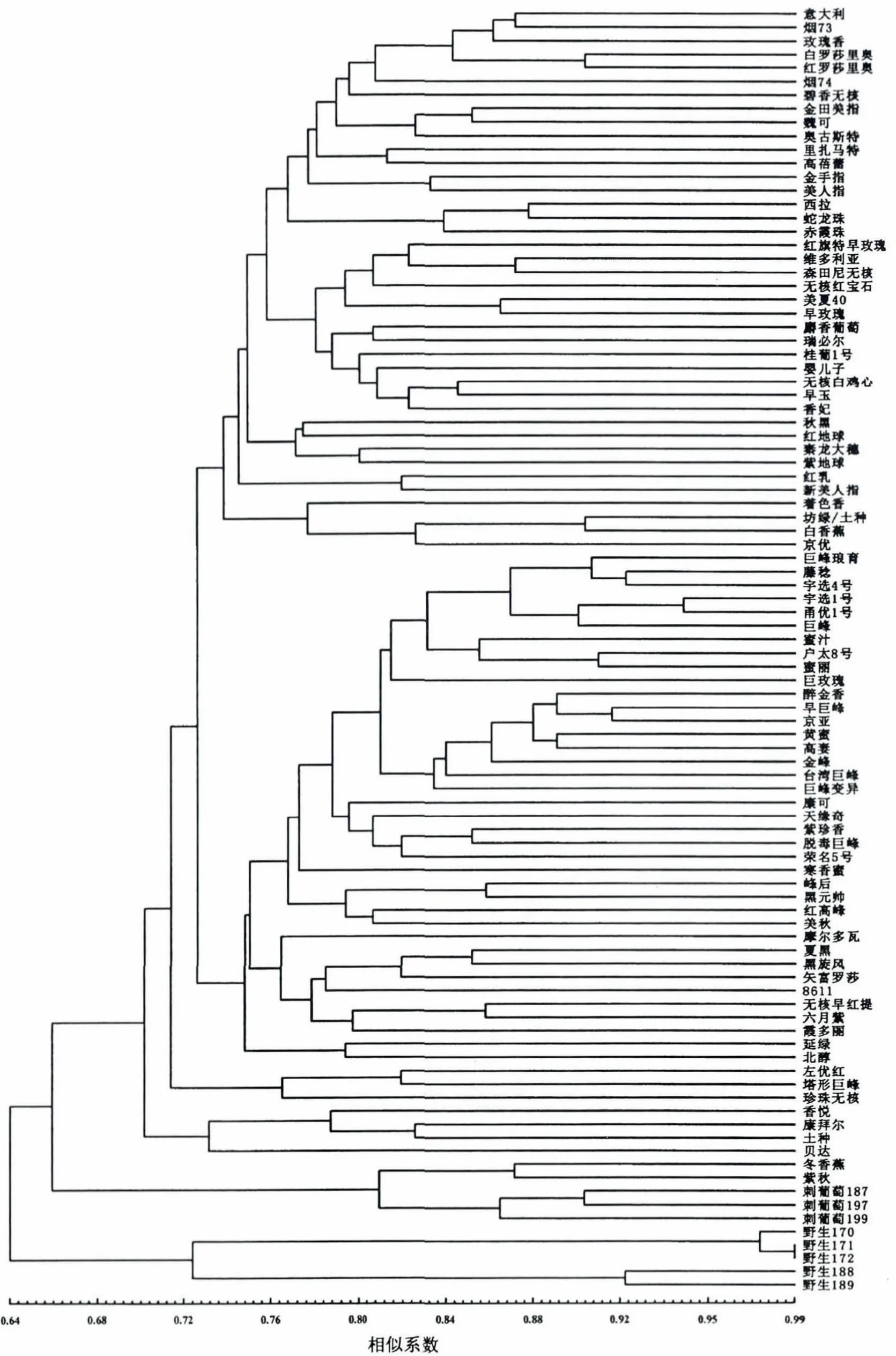
图2 95份葡萄品种资源的UPGMA聚类图Fig.2 The UPGMA phylogenetic tree of 95 grape variety resources
ISSR聚类结果与传统分类基本一致,按欧亚种、欧美杂种和东亚种群进行分组,由于美洲种收集的品种只有2个,未出现独立分支,而并入欧美杂种分支中.有36个葡萄品种资源聚类到欧亚种,在这36个品种中,有特殊玫瑰香味的葡萄品种“意大利”、“烟73”、“玫瑰香”(汉堡麝香)、“白罗莎里奥”、“红罗莎里奥”、“烟74”、“碧香无核”聚在1个分支上.相关研究资料表明,“意大利”为“比坎”ד玫瑰香”,是玫瑰香系第2代品种,“烟73”为“玫瑰香”ד紫北塞”、“白罗莎里奥”为“Rosaki”ד亚力山大红玫瑰”、“红罗莎里奥”为“Rosario Bianco”דRuby Okuyama”、“烟74”为“紫北塞”ד汉堡麝香”、“碧香无核”为“1851”ד莎巴珍珠”(“匈牙利玫瑰”ד奥托涅玫瑰”),这些品种都属麝香葡萄系的品种,在今后杂交育种中,如果育种的目的是获得有玫瑰香味的后代,根据育种的需要可以选择该分支中的品种作为亲本;“玫瑰香”与“白罗莎里奥”和“红罗莎里奥”亲缘关系较近,据《中国葡萄志》[1]记载,“玫瑰香”的原产地是英国,品种来源有3种记载:一是“黑罕”ד白玫瑰香”,二是“白玫瑰香”ד红玫瑰”(或其他黑色“Muscat”品种),三是亲本不详.但从聚类图可以看出,“玫瑰香”来源第2种的情况可能性较大.因此,通过此例可以看出,采用ISSR分子标记,可以对某些来源有争议的葡萄品种进行正本清源.
3个欧亚种酿酒葡萄品种“西拉”、“蛇龙珠”、“赤霞珠”在1个分支上,“蛇龙珠”的来源存在争议(属于“赤霞珠”或“品丽珠”的一个品系).根据聚类图可以得出,“蛇龙珠”与“赤霞珠”的亲缘关系较远(遗传相似系数0.825),而与“西拉”的亲缘关系较近(遗传相似系数0.875),说明其遗传上更接近“西拉”,由于未收集到“品丽珠”葡萄资源,对“蛇龙珠”与“品丽珠”的亲缘关系,有待进一步研究.
相关研究资料表明,“玫瑰香”和“麝香葡萄”是同一个品种,但在本研究中,聚类图显示其与“瑞必尔”亲缘关系最近,这可能与引种过程的品种混杂有关.由于相关资料未记载关于“瑞必尔”亲本的介绍,故无法进一步考究其关系.在聚类图中,相似的情况较多,如据报道,“秦龙大穗”是“里扎马特”(“可口甘”ד帕尔肯特”)芽变选育的品种,但在本研究中,“里扎马特”却与“高蓓蕾”聚在一起,而“秦龙大穗”与“秋黑”、“红地球”(C12-80×S45-48)、“紫地球”在相同分支下,由于资料有限,无法确定“里扎马特”是否与“红地球”或“秋黑”有溯源的相同亲本,但根据果实性状分析,“里扎马特”或“秦龙大穗”更接近“红地球”或“秋黑”.这种情况说明在我国葡萄品种引种过程中,品种的混杂较为普遍,同时也印证了王华等[2]的观点.在今后的葡萄杂交育种过程中,亲本选择时要避免品种混杂造成的失误,可行的方法就是收集不同地区同一品种名的葡萄品种,经ISSR分子标记分析后确认纯正的亲本.
在欧美杂种中,有4个品种“着色香”(“玫瑰露”ד罗也尔玫瑰”)、“坊绿”(建阳本地种)、“白香蕉”(品种来源和原名有不同观点,此品种亲本为“康可”ד麝香沙斯拉”或此品种应为“金玫瑰”)、“京优”[“黑奥林”(“巨峰”ד巨鲸”)实生苗]分为一组,其偏离欧美杂种而更接近于欧亚种,类似于从欧亚种到欧美杂种的过渡品种.“白香蕉”的品种来源存在争议,从ISSR聚类分析的情况看,其为“金玫瑰”的可能性更大.而且,被福建建阳当地长期种植保留的品种“坊绿”,其与“白香蕉”的亲缘关系很近,表明该品种并非当地原产品种,类似情况还有2个品种“延绿”和“土种”(从民间收集,未具品种名),前者与“北醇”亲缘关系很近,后者与“康拜尔”亲缘关系很接近,说明在很早以前,该地区民间就有零星引种葡萄的历史.
收集的欧美杂种中,“巨峰”系所占份额较大,有20多个品种(品系),从聚类图看,能比较清楚的反应其亲缘关系.在福建沙县南阳乡采集的1个葡萄品种,园主介绍其为从福安引种的“塔形巨峰”(可能是当地从巨峰系选育),据其品种名起初认为是“巨峰”系品种,但ISSR聚类分析表明其与“左优红”的亲缘关系很近,而“左优红”是由中国农业科学院特产研究所选育的“左山二”(山葡萄)与“小红玫瑰”的杂交种,是否该品种就是“左优红”,或是在引种中出现品种混杂,或是“巨峰”系品种与当地野生葡萄杂交的后代,有待进一步研究.
东亚种群有2个类群,分别为“刺葡萄”和“野生葡萄”,不同来源的“刺葡萄”聚在相同分支下,而从福建不同地方收集的“野生葡萄”聚在一起.尤其是从福安收集的2个百年野生葡萄品种(“野生188”和“野生189”),在南方高温多湿的环境条件下,经历了100多年仍能旺盛生长,表明其携带优良的抗性基因,如与现有优良栽培种杂交,有望培育出适合南方发展的高优葡萄品种.
对于欧山杂种而言,ISSR聚类分析的结果比较复杂,本研究中收集到3个欧山杂种,分别为“桂葡1号”(“毛葡萄”ד欧洲酿酒葡萄”)、“左优红”(“左山二”ד小红玫瑰”)和“北醇”(“玫瑰香”ד山葡萄”),“桂葡1号”聚类时在欧亚种中,而“左优红”和“北醇”聚到欧美杂种中,这种情况表明,欧山杂种与欧亚种或欧美杂种的亲缘关系,可能与其选育时所采用的多代回交亲本的选择有关,这与温景辉等[9]的相关研究相似.
本研究中,还发现“金手指”、“矢富罗莎”、“六月紫”、“霞多丽”聚类时出现异常情况.“金手指”(亲本不详)在相关品种介绍时属于欧美杂种[1],而却归类到欧亚种,从聚类图可知,“金手指”与“美人指”的亲缘关系很近,但由于缺乏“金手指”亲本的相关资料,很难进行判断.“矢富罗莎”[“潘诺尼亚”×(“莎巴珍珠”ד楼都玫瑰”)]、“六月紫”(山东早红芽变)、“霞多丽”(亲本不详)等3个品种介绍为欧亚种[1],但本研究结果为欧美杂种;“矢富罗莎”与“夏黑”亲缘关系较近,“六月紫”和“霞多丽”与“无核早红提”亲缘关系较近.这可能与品种混杂或多代回交亲本选择有关.
本研究通过ISSR分子标记绘制的葡萄品种资源聚类图,可为今后的葡萄杂交育种工作提供依据.通过ISSR可以确认大部分葡萄品种的亲缘关系,理清引种过程中因人为疏忽而造成的品种混杂情况,但个别品种还存在不确定性,需要分析引种源头和品种性状,并结合其他分子标记方法加以确认.在进行葡萄杂交育种实践前,可以通过ISSR分子标记对亲本进行前期亲缘关系确认,为葡萄杂交育种提供指导建议.
[1]孔庆山.中国葡萄志[M].北京:中国农业科学技术出版社,2004.
[2]王华,崔福君,张继澍,等.现代生物技木在葡萄种质资源、品种改良研究中的应用[J].中外葡萄与葡萄酒,2003(1):18-21.
[3]SABIR A,TANGOLAR S,BUYUKALACA S,et al.Ampelographic and molecular diversity among grapevine(Vitis spp.)cultivars[J].Czech J Genet Plant Breed,2009,45(4):160-168.
[4]YE G N,SPYLEMEZOGLU G,WEEDEN N F,et al.Analysis of the relationship between grapevine cultivars,reports and clones via DNA fingerprinting[J].Vitis,1998,37(1):33-38.
[5]何永华,李朝銮,曹亚玲.葡萄属营养器官的比较解剖学及其系统学意义[J].植物分类学报,1994,32(2):154-164.
[6]牛立新,贺普超.我国葡萄属野生种形态特性的研究[J].葡萄栽培与酿酒,1995(4):15-17.
[7]李朝銮,曹亚玲,何永华.中国葡萄属(Vitis L.)分类研究[J].应用与环境生物学报,1996,2(3):234-253.
[8]刘三军,孔庆山,顾红.我国葡萄属植物孢粉学分类研究[J].果树科学,1997,14(2):100-105.
[9]刘三军,孔庆山,顾红,等.我国野生葡萄过氧化物酶同工酶研究[J].果树学报,1998,15(4):322-326.
[10]SANTIAGO J L,BOSO S,MARTÍN J P,et al.Characterisation and identification of grapevine cultivars(Vitis vinifera L.)from northwestern Spain using microsatellite markers and ampelometric methods[J].Vitis,2005,44(2):67-72.
[11]吕秀兰,张光伦,龚荣高,等.22个葡萄品种过氧化物同工酶研究[J].四川农业大学学报,2005,23(2):182-185.
[12]王姣,刘崇怀,樊秀彩,等.葡萄种类和品种鉴定技术研究进展[J].植物遗传资源学报,2008,9(3):401-405.
[13]刘崇怀,冯建灿,姜建福.中国葡萄属植物形态学聚类分组研究[J].植物遗传资源学报,2011,12(6):847-852.
[14]王玉娟,张彦,房经贵,等.利用基于RAPD标记的MCID法快速鉴定72个葡萄品种[J].中国农业科学,2012,45(14):2913-2922.
[15]温景辉,申海林,邹利人,等.20份葡萄种质亲缘关系的SSR分析[J].果树学报,2011,28(5):782-786.
[16]王倩,王斌.DNA分子标记在果树遗传学研究上的应用[J].遗传,2000,22(5):339-344.
[17]王军.生物技术与葡萄遗传育种[J].中国农业科学,2009,42(8):2862-2874.
[18]张青林,罗正荣.ISSR 及其在果树上的应用[J].果树学报,2004,21(1):54-58.
[19]QU X P,LU J,LAMIKANRA O.Genetic diversity in muscadine and american bunch grapes based on randomly amplified polymorphic DNA(RAPD)analysis[J].J Amer Soc Hort Sci,1996,121(6):1020-1023.
[20]王跃进,LAMIKANRA O,SCHELL L,等.用RAPD分析鉴定葡萄属远缘杂种[J].西北农业大学学报,1997,25(3):16-20.
[21]张立平,林伯年,沈德绪,等.葡萄属 RAPD 分类研究[J].园艺学报,1998,25(2):191-193.
[22]CERVERA M T,CABEZAS J A,SANCHA J C,et al.Application of AFLPs to the characterization of grapevine Vitis vinifera L.genetic resources:A case study with accessions from Rioja(Spain)[J].Theoretical and Applied Genetics,1998,97(1/2):51-59.
[23]高爱农,韩振海,张开春,等.RAPD技术在葡萄种质鉴定上的应用[J].农业生物技术学报,2002,10(2):133-137.
[24]鲍露,徐昌杰,江文彬,等.葡萄AFLP技术体系建立及其在超藤与藤稔葡萄品种鉴别中的应用[J].果树学报,2005,22(4):422-425.
[25]吴子龙,方连玉,王军,等.15份葡萄种质亲缘关系的 ISSR分析[J].果树学报,2006,23(4):605-608.
[26]DHANE M,TAMHANKAR S A,PATIL S G,et al.Assessment of genetic diversity and relationships among some grape varieties using ISSR markers[J].Journal of Applied Horticulture,2006,8(1):50-52.
[27]刘闯萍,王军.SSR标记及其在葡萄上的应用[J].果树学报,2008,25(1):93-101.
[28]ALIZADEH M,SINGH S K.Molecular assessment of clonal fidelity in micropropagated grape(Vitis spp.)rootstock genotypes using RAPD and ISSR markers[J].Iranian Journal of Biotechnology,2009,7(1):37-44.
[29]ARGADE N C,TAMHANKAR S A,KARIBASAPPA G S,et al.DNA Profiling and Assessment of Genetic Relationships A-mong Important Seedless Grape(Vitis vinifera)Varieties in India Using ISSR Markers[J].J Plant Biochemistry & Biotechnology,2009,18(1):45-51.
[30]HASSAN N A,EL-HOMOSANY A,GOMMA A H,et al.Morphological and Issr Polymorphisms in Some Egyptian Grapes(Vitis vinefera L.)Collection[J].World Applied Sciences Journal,2011,15(10):1369-1375.
[31]WEN J H,SHEN H L,ZOU L R,et al.Analysis of genetic diversity of Vitis amurensis Rupr.(Vitaceae)germplasm resources[J].Agricultural Biotechnology,2012,1(5):27-29.
[32]赖呈纯,范丽华,谢鸿根,等.葡萄ISSR-PCR反应体系的建立与优化[J].生物技术通报,2012(2):159-164.
[33]杨治元.葡萄100个品种特性与栽培[M].北京:中国农业出版社,2007.
[34]王军,贺普超.山葡萄基因组DNA提取及RAPD鉴定[J].果树科学,2000,17(2):79-82.
[35]张永辉,刘崇怀,樊秀彩,等.ISSR标记在中国野生葡萄分类中的应用[J].果树学报,2011,28(3):406-412.
