假俭草生物量季节动态研究
2014-12-12徐礼英周守标柳后起
徐礼英 周守标 柳后起
(1 芜湖职业技术学院园林园艺系,安徽 芜湖 241003)(2 安徽师范大学重要生物资源保护与利用研究重点实验室,安徽 芜湖 241000)
假俭草(remochloa ophiuroides)属C4结构暖季型多年生禾本草植物,植株低矮、叶形优美,是世界三大暖季型草坪草之一[1]。其原产于中国中南部,但现已广泛分布于世界各地[2,3]。近年来国内外学者多从生理、分子及遗传变异等角度,研究假俭草的抗寒性、抗旱性等抗逆性机理,相关报道较多,而对假俭草各构件生物量动态变化进行系统的研究未见报道。植物的各部分是一个统一的整体[4],地上生长与地下生长既互相依存,又互相竞争,构成相互协调又与环境条件相适应的有机整体[5],植物种群个体和构件生物量的动态是植物与环境因素共同作用的结果,它既反映了植物种群对环境条件的适应能力和生长发育规律,也反映了环境条件对植物种群影响和饰变程度[6,7,8]。 因此系统研究假俭草生物量变化规律,为其牧草资源开发、草坪建植管理,以及改善农牧业生态环境、促进畜牧业发展和退化生态系统的恢复与重建具有重要意义。本文对假俭草草地进行定期取样测定,以揭示植株各构件生物量季节变化规律,为假俭草在草坪建植与水土保持等方面提供科学依据。
1 材料与方法
1.1 研究地的自然概况
在安徽省芜湖市神山公园取研究样地,本地处 117°57′—118°54′E,30°38′—31°34′N 的长江沿江平原,属于温带大陆性气候,年平均气温15.7~16.0℃,极端高温达 41℃,极端最低温-15℃,年降雨量1198~1413mm,主要集中在夏季,年日照时数2000 h,无霜期7~8个月[9]。研究样地在神山公园内一个未被开发利用的典型假俭草地段上,该处地势平坦,生境条件、植物群落组成、结构及利用强度一致。以假俭草为优势种,另外伴生有少量结缕草、狗牙根、知风草、鸡眼草等,周围乔木有樟树、悬铃木、刺槐等,另有白檀、野蔷薇、火棘等灌木,土壤类型为砂壤土。
1.2 研究方法与材料处理
通过定位监测,样地围建在以假俭草为主的典型地段上[10]。自2013年4月至11月,间隔一个月取样一次。
地上生物量测定:每次10个重复随机选取25cm×25cm×20cm的样方土柱,将样块放入纱网后置水中充分浸泡[11],冲洗干净。按叶片、叶鞘、匍匐茎、根、凋落物和穗等构件分开分别装入纸袋内,80℃烘至恒重后称重,计算各构件生物量占种群总生物量的比例[12]。
地下生物量测定:采用分层取样法,每次10个重复随机选取15cm×15cm样方挖沟分层,每1cm深为1层,共分10层。将根系土壤混合体放入铁筛中,反复冲洗,拣出异物,在75℃的温度下将样品烘至衡重后称取干重,计算各层根生物量占种群总根生物量的比例[13]。
1.3 数据处理
利用EXCEL和SPSS软件对观测构件质量的分配比例、季节变化规律以及构件之间的相关关系进行统计和分析。
2 结果与分析
2.1 假俭草地上、地下及总生物量的季节动态
分析图1可知,假俭草种群自4月返青后,根系开始迅速合成和储存能量,地上部分利用良好的水热、光照条件及向上转移的地下储存物质,一直到6月份,植物体迅速生长,4~6月份间地上、地下及总生物量显著增加,差异极显著(P<0.05),此后,地上部分生长趋向相对平稳,至9月份地上生物量达到最大值,随着气温降低,植物体生长趋向停止并开始出现枯萎,地上生物量开始明显下降,月份间差异显著(P<0.05),而地下生物量迅速积累于6月份达到最大值后,渐渐下降直至枯萎,由于在此期间地上部分相对生物量较大,需消耗大量营养成分,地下部分营养物质又重新向上转移,以满足地上部分生长的需要,虽然地上部分合成的生物量较多,但多数用于自身生长和消耗的需求,向下转移有限,致使地下生物量降低。
总生物量变化是地上、地下生物量变化的综合表现,因此在返青初期至6月份,生物量迅速增加,在9月份达到最大值,虽然6~9月份植物体合成的物质较多,但由于植物体的向外扩展和匍匐茎的延伸消耗了大量的储存物质,导致总生物量出现负增长,但之后,由于植株逐渐衰老、地上部分开始枯萎、地下部分的自身消耗,生物量逐渐下降。由于上述原因,假俭草地上、地下生物量的比值情况如图1所示,即4~9月呈上升趋势,9月份后一直下降。

图1 假俭草地下、地上及总生物量的季节动态
2.2 假俭草各构件生物量分配及占总生物量百分比的季节动态
综合分析图2与图3可知,在假俭草整个生长期中,匍匐茎生物量分配在种群生物量中始终占有绝对优势,保持在30%以上,而根的生物量分配比例较低,在15%左右;叶片和匍匐茎所占总生物量比例随季节变化波动较大,根和叶鞘占有相对稳定的比例。
假俭草营养生长早期,匍匐茎储存物质增加较快,生物量迅速上升,7月份开始,因地上部生长迅速,消耗营养物质大幅度增加,为了满足叶片、叶鞘快速生长的需要,地下部分营养物质开始部分向上转移,导致自身生物量及所占总生物量的比例有所下降,9月份以后,虽然地上部分生
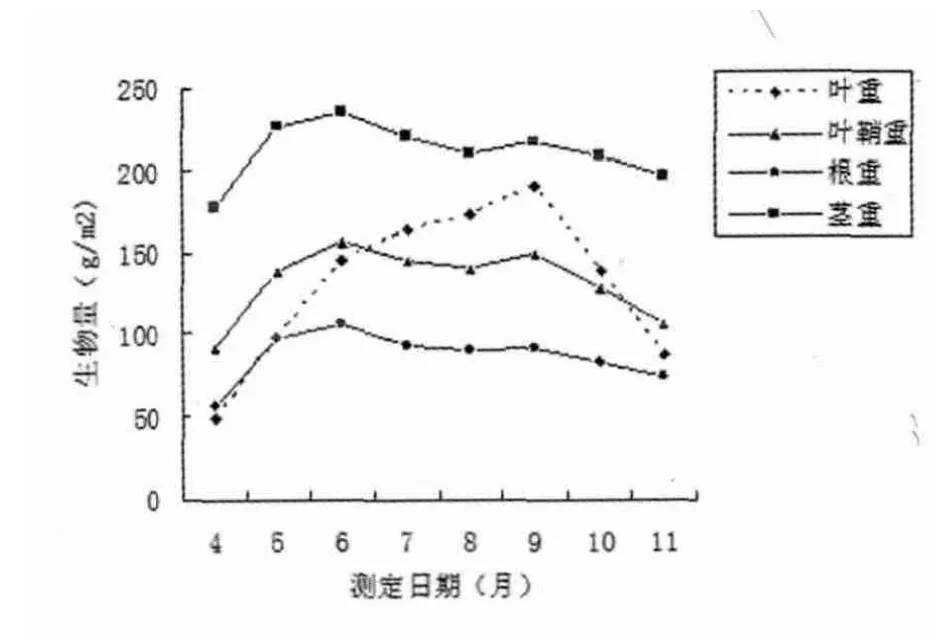
图2 假俭草各构件生物量的季节动态
2.3 假俭草地上、地下部分及总生物量的生长率
生长率是衡量生物净积累量,绝对生长率(AGR)为单位时间内单位面积生物量的净累积量,AGR=(W2-W1)/(t2-t1);相对生长率(RGR)则说明单位生物量单位时间的净积累量,RGR=(LnW2-LnW1)/(t2-t1)[6],其中 W1、W2 分别表示t1和t2时刻的生物量,LnW1和LnW2为t1和t2时刻生物量的对数,AGR和RGR的单位分别为g/m2.d 和 g/g.d[14]。
假俭草地上、地下部分及总生物量的生长率见图2,根据图2绘制季节动态图,见图4。分析图2和图4可知,在4~6月期间假俭草种群地上部分生长迅速,地下储存营养物质的部分向上转移,植物生长迅速,地上、地上及总生物量的绝对生长速率和相对生长率为正值,但呈下降趋势;长趋向停止,消耗营养物质开始变少,但此时植物体将部分能量用于生殖生长,导致匍匐茎生物量仍呈下降趋势,此时因总生物量下降较快,所以占总生物量比例呈上升状态,方差检验表明,假俭草匍匐茎的生物量及占总生物量比例在月份之间存在显著差异(P﹤0.05);叶片生物量及占总生物量分配自返青后保持上升趋势,到9月份达到最大,随着气温降低,叶片开始枯萎,生物量逐渐下降,叶片生物量及占总生物量比例及在月份之间呈现极显著的差异(P﹤0.05);叶鞘与根的生物量变化趋势相似,4~6月份上升,再平稳变化,9月份后呈下降趋势,生物量在月份间差异极显著(P﹤0.05),但占总生物量的百分比相对比较稳定,且月份间都无明显差异。在6~8月期间,地下部分及总生物量的绝对生长速率和相对生长率呈现负值,但呈增加趋势,并在8月份变为正增长,而地上生物量的绝对生长速率和相对生长率在6~8月期间一直保持正增长呈增加趋势;9月份后各生长速率转为负增长并逐渐下降。

图3 假俭草生物量分配的季节动态
因此9月份地上部分生物量与总生物量达到了最大值,此后呈负增长趋势,它们不断下降。地下生物量在4~6月期间呈增长趋势,但变化幅度小于地上部分,自7月开始,由于地上部分生长迅速消耗了大量的营养物质,地下部分生物量有所减少,但此时因地上部分同时合成了大量的营养物质并部分向下转移,地下生物量相对率和绝对生长率接近零,变化幅度较小,9月以后,地下生物量无显著变化,绝对生长率趋向于零,相对增长速率呈负值并呈下降趋势。
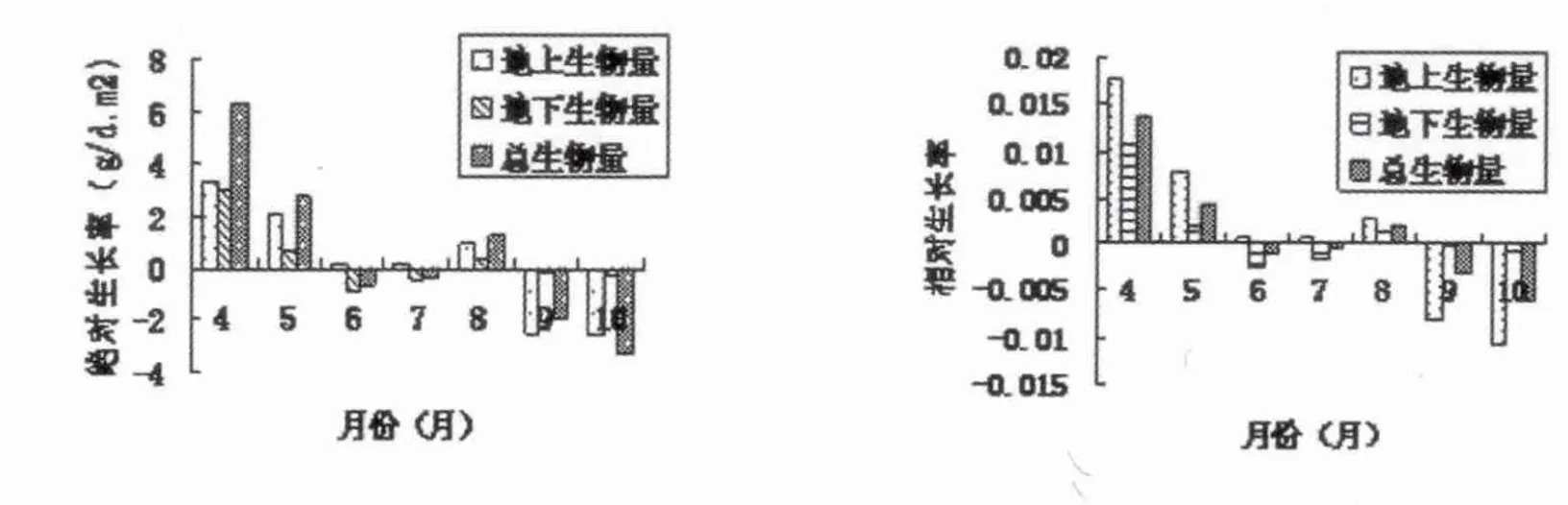
图4 假俭草地上、地下及总生物量生长率的季节动态
2.4 假俭草根生物量的垂直分布及根冠比的季节动态
2.4.1 假俭草根生物量的垂直分布
假俭草种群的须根系主要分布在地表以下的浅层土壤区域,由浅到深呈“T”分布(如图5所示),其中根生物量的50%分布在地表以下2厘米的范围内,地表至地表下5厘米的范围内根的生物量达到了总体生物量的90%以上。
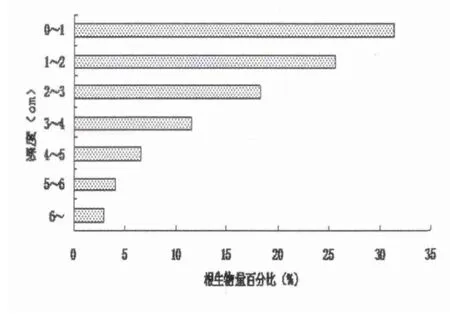
图5 假俭草根生物量的垂直分布
2.4.2 假俭草根冠比的季节动态
根冠比反映了植物地下部分与地上部分的生长相关性,在假俭草返青后的生长前期,根系生物量在较短时间内迅速增加达到了较高水平,同时地上部分生物量刚刚开始生长积累,相对较少,故根冠比较大;5~9月期间,根系趋向稳定并有所下降,此时假俭草地上部分生长迅速,合成及储存生物量较大,导致根冠比明显下降,并在9月达到了最小值,此后,随着地上部开始枯萎及植物生长趋于停止,根冠比又开始上升 (如图6所示)。
3 讨论
3.1 假俭草构件生物量的季节动态

图6 假俭草根冠比的季节动态
假俭草种群自4月返青至6月期间,植物迅速生长,地上、地下及总生物量都迅速增加,而在6~9月期间,变化幅度不大,地上部分生物量在9月达到最大值,地下部分生物量于6月份达到最大,总生物量在6月和9月出现两个生长高峰。9月之后,随着气温的下降,各测定的生物量也开始下降。
在6~8月期间,地下部分及总生物量的绝对生长速率和相对生长率呈现负值,但呈增加趋势,并在8月份变为正增长,这可能在6、7两月雨水过多,对根系的生长影响很大,同时因地上部分生长迅速消耗了大量的营养物质,导致地下部分生物量有所减少,且绝对生长速率和相对生长率呈现负值,8月份雨水减少,天气炎热,大多植物地下部分恢复正生长,而地上部分自4月返青迅速生长直到9月,因此地下部分及总生物量的绝对生长速率和相对生长率在8月份变为正增长。
假俭草种群自返春后,匍匐茎生物量迅速增加到6月份达到最大值,且在整个生长期中,在种群生物量分配比中始终占有绝对优势,保持在30%以上,叶片生物量一直增加到9月达到最大值,匍匐茎和叶片的生物量及占总生物量比例在月份之间都存在显著差异(P﹤0.05),根生物量于6月份达到最大,此后下降,8月份出现最低值,且根生物量在月份间差异极显著(P﹤0.05),但占总生物量的百分比相对比较稳定,且月份间都无显著差异(P>0.05);根冠比在生长前期变化不明显,5~9月期间,根冠比明显下降,9月以后上升。
3.2 假俭草根的垂直分布
植物所需的大量营养物质和水分是植物根系从土壤中吸收所得,除自身需要外,还将所贮藏的营养物质一起运输到植物的各个部分,供整个植物的生长发育,对地上生物量的形成起着重要的作用,是草地生态系统物质循环和能量流动不可缺少的环节[15]。植物根系分布通常为倒金字塔型,即由深到浅呈“T”形分布[4,13,16]。
从以上分析可知假俭草种群的须根系主要分布在地表,由浅到深呈“T”型分布,姜海楼及董瑞音等研究了根蘖类型植物麻黄的整个根系呈“T”型分布,约80%地下生物量分布于0~60 cm土层内,即有横走的水平根,还有垂直向下的垂向根。这种庞大的根系,可保证对地上枝条水分和养分的供给,促进其生长发育,增强抗旱、抗寒能力[16]。而假俭草种群根生物量的50%分布在地表以下2厘米的范围内,地表以下5厘米范围内根的生物量达到了根总生物量的90%以上,假俭草根系的这种分布状况与生境密切联系,在未开发的研究样地上,大部分有机质和养分都集中于土壤表层,假俭草的主要根系于该层能够获得大量的物质与能量,满足植物生长需求。
综上所述假俭草种群成坪速度较快,匍匐茎发达具耐践踏,绿期长达7个月左右,是草坪建植的优良品种。在建植草坪时,建议在地表以下5厘米左右施肥,根系更好地向深层分布,发挥假俭草在草坪建植及治理水土流失的生态建设中独特的功效。
[1]Hitchcock A S.Manual of the grass of the United States[M].Washington DC:US Dept Agric,Agric Res Admin,1951.
[2]Hanna W W,Burton W G.Reproductive behavior and fertility characteristics of centipedegrass[J].CropSci.,1978,(18):835-837.
[3]Duble R L.Turfgrass management and use in the Southern Zone[M].Texas A,M University Press,1996.
[4]王代军,黄文惠,苏加楷.多年生黑麦草和白三叶人工草地生物量动态研究[J].草地学报,1995,(2):135-143.
[5]张娜,梁一民.黄土丘陵区天然草地地下/地上生物量的研究[J].草业学报,2002,(2):72-78.
[6]陈灵芝,任继凯,鲍显成,等.北京西山(卧佛寺附近)人工油松林群落学特性及生物量的研究[J].植物生态学与地植物学丛刊,1984,(3):173-181.
[7]方精云,刘国华,徐嵩龄.我国森林植被的生物量和净生产量[J].生态学报,1996,(5):497-508.
[8]殷淑燕,刘玉成.大头茶构件种群生物量及叶面积动态[J].植物生态学报,1997,(1):83-89.
[9]安徽植被协作组.安徽植被[M].合肥:安徽科学技术出版社,1981:22-154.
[10]刘永志,邢旗,姜永.科尔沁草原地上生物量动态的研究[J].内蒙古草业,1996,(Z2):36-45.
[11]许志信,曲永全,白飞.草甸草原12种牧草生长发育规律和草群地上生物量变化动态研究[J].内蒙古农业大学学报,2001,(20):28-32.
[12]王仁忠,祖元刚.羊草种群生物量和能量生殖分配的研究[J].植物研究,2001,(2):299-303.
[13]姜海楼,董瑞音,贾长友,等.麻黄生物学特性及生物量研究[J].草业学报,1997,(1):18-22.
[14]姜恕编著.草地生态学研究方法[M].北京:农业出版社,1988:66-70.
[15]胡中民,樊江文,钟华平,等.中国草地地下生物量研究进展[J].生态学杂志,2005,(9):1095-1101.
[16]王启基,周兴民,张堰清,等.高寒小嵩草草原化草甸植物群落结构特征及其生物量[J].植物生态学报,1995,(3):225-235.
