五种管口类纤毛虫的形态学研究*
2014-12-02潘宏博马洪刚许恒龙
赵 璐,潘宏博,马洪刚,许恒龙
(1.中国海洋大学海洋生物多样性与进化研究所,山东 青岛 266003;2.上海海洋大学水产与生命学院,上海 201306)
管口类纤毛虫是周丛生原生动物的重要组成部分。根据Lynn[1]的分类系统,其隶属于纤毛门(Ciliophora)、叶咽纲(Phyllopharyngea)、管口亚纲(Cyrtophoria)。除具有纤毛虫的普遍性状外,该类群具有独特的咽管状口器,纤毛集中于腹侧,且大核具有异质性[2]。近10年的分子系统学研究普遍认为该类群是一个外延清晰的单源发生性分类阶元[3-5]。目前管口亚纲已有近150种,但由于前人实验条件和方法的制约,大部分种类缺失重要的活体信息(如:伸缩泡数目及分布,咽杆数目等)和纤毛图式资料,急需详尽的重描述[2,6-7]。本工作为近年我国海洋纤毛虫多样性调查的一部分,涉及5种已知种类,其中尖绒毛虫、阿氏直毛虫和近岸类偏体虫均为南海新纪录种。
1 材料与方法
本文所涉纤毛虫的采样地和采样方法较为多样。乙状轮毛虫(Trochiliasigmoides)采于青岛栈桥风景区,在退潮后通过软毛刷轻轻刷取潮间带岩石表面积水获得样品;尖绒毛虫(Microxysmaacutum)采用悬挂载玻片法[8]于青岛小港的水体中取得;阿氏直毛虫(Orthotrochiliaagamalievi)利用直接采水法采于青岛第一海水浴场;近岸类偏体虫(Agnathodysterialittoralis)采自湛江东海岛养虾池;斯雷腹沟虫(Coeloperix sleighi)采于湛江沿海潮间带退潮后的积水。样品采集后,在实验室内室温(约20℃)培养2~3d。具体采集、分离、培养及形态观察的方法见文献[8],蛋白银染色参考文献[9],名词术语参照文献[1-2]。
2 结果与讨论
2.1 乙状轮毛虫(见图1A-B,E-J)
2.1.1 形态学描述 活体大小约(20~28)μm×(15~20)μm,阔卵圆形;左右侧扁,宽厚比约1∶2。体左侧扁平,右侧略隆起,具5~7条明显的肋纹(见图1A,F,长箭头)。体后端具矛状附着器。伸缩泡2个,位于体前后各1/3处。纤毛长约5μm。纤毛图式极为稳定:恒有4条右区动基列,其中2条腹前动基列几乎等长(见图1B,H);围口动基列(见图1B,H,长箭头)和左前动基列各2条,在口前方呈“八”字形排布(见图1B,J,短箭头)。大核椭球形,并列异质,染色后大小约为10μm×5μm。虫体运动相对缓慢,常于基质上缓慢爬行。
2.1.2 比较与讨论 轮毛虫属目前已知纤毛图式的种类仅4种,即小轮毛虫(Trochiliaminuta)、乙状轮毛虫、彼氏轮毛虫(Trochiliapetrani)和膜泡轮毛虫(Trochiliaalveolata)。它们的纤毛图式基本一致:4列右区动基列,2列围口动基列,2列左前动基列,其区别仅在于活体大小、背面肋纹和表膜泡有无[4,10]。
2.2 尖绒毛虫(见图1C-D,K-N)
2.2.1 形态学描述 活体大小约(25~45)μm×(15~25)μm,虫体卵圆形,前端钝圆,尾端较尖;左右侧扁约1∶1.5;背方明显向外隆起,腹方较平直(见图1C,K)。胞口位于腹面前端,略向外突起(见图1M,长箭头)。矛状附着器位于尾端(见图1K,短箭头)。虫体无色透明,内含多个食物泡。伸缩泡1枚,位于体前1/3处(见图1N,短箭头)。11~12条体动基列,其中腹前动基列恒为2条并延伸到体前端。最左侧3条动基列相对较长,约由6~10个毛基粒组成(见图1D,L,长箭头),其余动基列短小,仅含3~6个毛基粒。围口动基列2条,平行排布于胞口前方(见图1D)。赤道动基列片段约由2~7个毛基粒组成。大核卵圆形,染色后约为10μm×5μm。该种常以附着器附着于菌丝上绕体轴旋转。
2.2.2 比较与讨论 本种为南海新纪录种,其形态特征同Deroux[11]的原始描述完全一致,但香港种群同龚骏在2005年报道的青岛种群[10]相比缺少一枚伸缩泡(1vs.2)。通常在仅具1~2个伸缩泡的管口类中,伸缩泡的数目极为稳定。但是考虑到其他特征完全一致,且缺乏分子生物学数据,单纯依靠一个性状无法确定二者是不同种。故暂推断是种群间的差异。
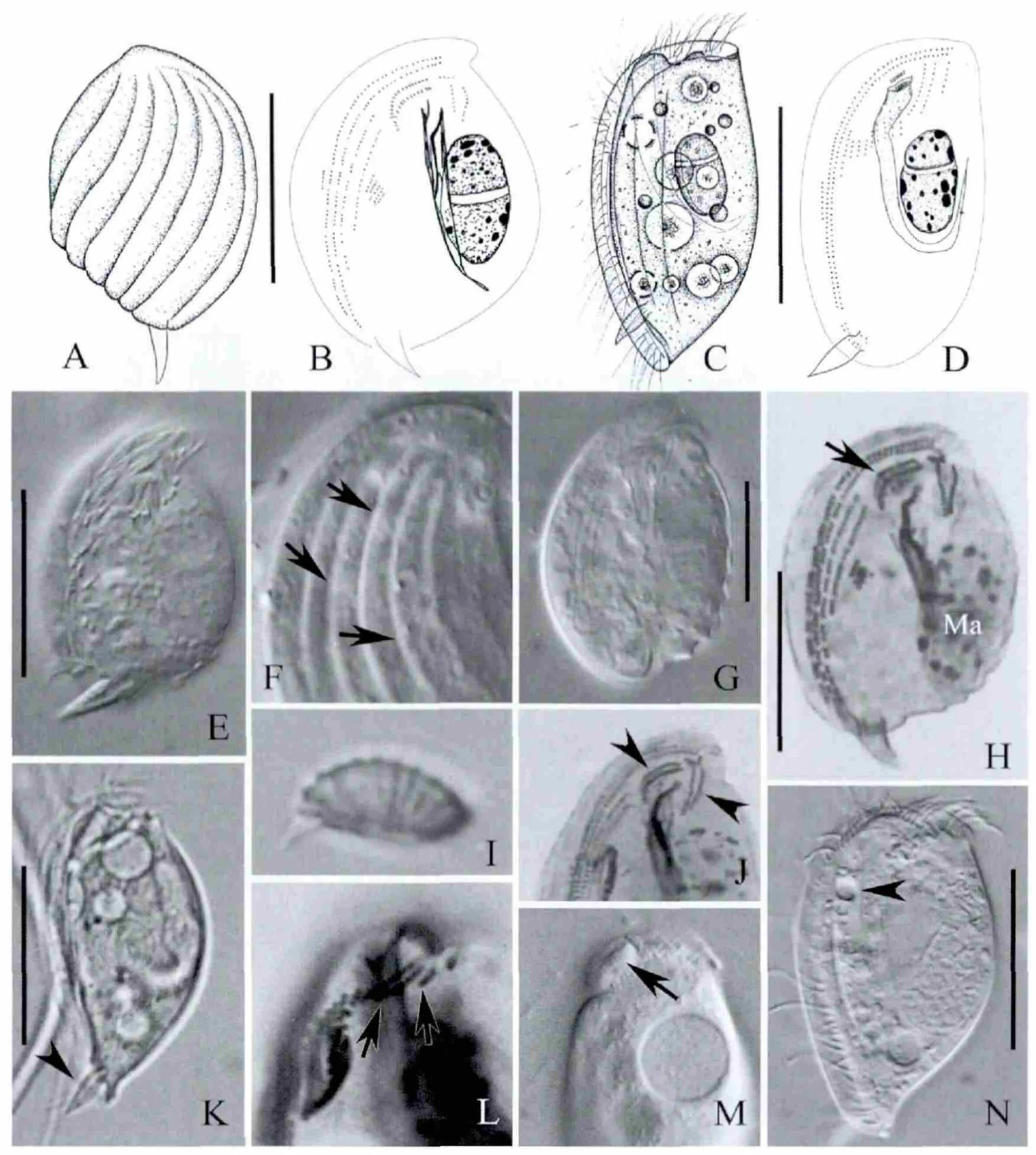
图1 乙状轮毛虫(A,B,E~J)和尖绒毛虫的活体和蛋白银染色后显微照片(C,D,K~N)Fig.1 Trochilia sigmoides(A,B,E~J)and Microxysma acutum (C,D,K~N)from life and after protargol impregnation
2.3 阿氏直毛虫(见图2A-B,E-K)

图2 阿氏直毛虫(A,B,E~H)和海滨类偏体虫(C,D,L~P)的活体及蛋白银染色后的显微照片Fig.2 Orthotrochilia agamalievi(A,B,E~H)and Agnathodysteria littoralis(C,D,L~P)from life and after protargol impregnation
2.3.1 形态学描述 活体大小约40μm×20μm,染色后约为27μm×15μm,虫体瘦卵圆形,背腹扁平,前端钝圆,后端渐窄(见图2A,E)。2个伸缩泡位于体右侧前、后1/3处(见图2G,长箭头)。虫体中央具单枚卵圆形大核(Ma),并列异质,约12μm×7μm大小(见图2F,短箭头,G)。体动基列共12~14条,在腹面连续排布。其中腹前动基列2条;其余动基列前端几乎在胞口下方同一水平。最右侧3条体动基列末端向后延伸至体缘并终止于同一水平线,其他动基列后端从右到左急剧缩短。背端动基列片段位于腹前动基列前端附近,约含5个毛基粒。围口动基列2条,位于胞口前方“八”字排布(见图2B,J,短箭头)。蛋白银染色后,附着器基部常可见毛基粒样嗜银颗粒(见图2K,短箭头)。
2.3.2 比较与讨论 该种为南海新纪录种,其同直毛虫属另一已知种环状直毛虫(Orthotrochiliapilula)[12]的区别主要在于体动基列数目(12~14列 vs.9列)和口器的咽杆数目(约40vs.6个)。
2.4 近岸类偏体虫(见图2C-D,L-P)
2.4.1 形态学描述 虫体无色透明,左右侧扁,活体大小约为(35~45)μm×(20~30)μm。腹面观(见图2M)为长椭圆形,侧面观(见图2C,E)为近半圆形。胞口圆形,位于腹前1/4处;咽杆恒为6根。附着器矛头状,位于腹面末端偏左位置,长约6~8μm(见图2L)。伸缩泡2个,位于体右侧前、后1/3处。单一大核于虫体前部。纤毛图式如图2J~L所示。稳定9条右区动基列,其后端从中间向两侧逐渐缩短,最右侧3列前伸超过口区并弯向背方,其他6列前端于胞口后方终止。附着器基部具若干嗜银颗粒(见图2K,短箭头)。2条围口动基列和1条口前动基列几乎等长,横向排于口前(见图2D,N,短箭头);左前动基列恒为3条(见图2D,O,短箭头)。
2.4.2 比较与讨论 本种为南海新纪录种,在体形、大小和纤毛图式方面同Deroux[11]的原始描述基本吻合,仅伸缩泡数目略有差别(2vs.1)。龚骏[10]于青岛也曾采到该种,但同样伸缩泡仅1枚。考虑到其余性状完全一致,伸缩泡数目在不同种群间偶尔会存在细微差异(如尖绒毛虫,见上文),故推断鉴定无误。
2.5 斯雷腹沟虫(见图3A-H)
2.5.1 形态学描述 活体大小(35~50)μm×(25~35)μm,虫体卵圆形或肾形。体周边的横纹条带于赤道处分成2段(见图3B,长箭头)。咽杆12~16条。2枚伸缩泡,对角线位(见图3A,D)。大核椭球形,位于体中央。体动基列被口区分为前后两部分:前部稳定具有4条弧状体动基列,横向排布;后部具15或16条动基列,并在体后部形成不太明显的缝合线(见图3H,短箭头)。伸缩泡2枚,对角线位,蛋白银染色后伸缩泡开口通常可见(见图3E,长箭头)。口区动基列3条,1前2后,均为双动基系,横向排布于口前。(见图3G,长箭头)。

图3 斯雷腹沟虫活体(A~D,F)及蛋白银染色后(E,G,H)照片Fig.3 Photomicrographs of Coeloperix sleighi from life(A~D,F)and after protargol impregnation(E,G,H)
2.5.2 比较与讨论 本南海种群与龚骏的青岛种群[13]形态上完全吻合。
[1]Lynn D.The Ciliated Protozoa,Characterization,Classification and Guide to the Literature.3rdEdition[M].New York:Springer,2008:348-350.
[2]龚骏,宋微波.青岛沿海的管口类纤毛虫(原生动物,纤毛门,管口目)[J].动物分类学报,2009,34(4):949-953.
[3]Gao S,Huang J,Li J,et al.Molecular phylogeny of the cyrtophorid ciliates(Protozoa,Ciliophora,Phyllopharyngea)[J].Plos One,2012,7(3):33198.
[4]Pan H,Lin X,Gong J,et al.Taxonomy of five species of cyrtophorids(Protozoa:Ciliophora)including consideration of the phylogeny of two new genera[J].Zool J-Linn Soc,2012,164:1-17.
[5]Gong J,Stoeck T,Yi Z,et al.Small subunit RNA phylogenies show that the class Nassophorea is not Monophyletic(Phylum Ciliophora)[J].J Eukaryot Microbiol,2009,56:339-347.
[6]龚骏.青岛沿海管口目纤毛虫的分类学研究及科属级阶元的系统修订[D].青岛:中国海洋大学,2005.
[7]潘宏博.中国海洋管口目和侧口目纤毛虫多样性的研究[D].青岛:中国海洋大学,2011.
[8]宋微波,徐奎栋.纤毛虫原生动物形态学研究的常用方法[J].海洋科学,1994,6:6-9.
[9]Wilbert N.Heine verbesserte technik der protargolimpragnation für ciliaten[J].Mikrokosmos,1975,64:171-179.
[10]龚骏,沃伦 A,宋微波.管口类[M].//宋微波,沃伦 A,胡晓钟.中国黄渤海的自由生纤毛虫.第一版.北京:科学出版社,2009:49-92.
[11]Deroux G.Le plan cortical des Cyrtophorida.III.Les structures différenciatrices chez les Dysteriina[J].Protistologica,1976,12:505-538.
[12]Song W.On two marine cyrtophorid ciliates from China,with description ofChlamydonelladerouxinov.spec.andOrthotrochiliapilula(Deroux,1976)nov.comb.,and reestablishment of the genusOrthotrochilianov.gen.(Protozoa,Ciliophora,Cyrtophorida)[J].Hydrobiologia,2003,499:169-177.
[13]Gong J,Song W.Re-establishment of the cyrtophorid genusCoeloperixDeroux,nov.gen.,with a description ofCoeloperix sleighinov.spec.(Protozoa,Ciliophora,Cyrtophorida)[J].Europ J Protist,2004,40:175-181.
