DNA条形码技术在黑瞎子岛鼠类鉴定中的应用*
2014-11-10刘润吉张荣波刘丽娟呼满霞曹晓梅郭天宇张晓龙
刘润吉 张荣波 李 明 刘丽娟 呼满霞 王 静 曹晓梅 郭天宇 张晓龙**
(1.安徽理工大学,安徽淮南 232000;2.中国检验检疫科学研究院,北京 100123;3.黑龙江出入境检验检疫局,哈尔滨 150001)
传统形态分类方法用于日常物种鉴定,会出现表型可塑性和遗传变异性引起的鉴定错误、忽略隐存种、受限于个体发育阶段和性别等4个明显的局限性(Hebertetal.,2003)。传统形态学分类的固有局限性、地球上多达1000万的庞大物种数量和不断缩减的分类学家队伍,预示着对一种新的分类方案的巨大需求。20世纪后半叶,遗传物质核酸登上了自然科学史的舞台,人们对基因的认识和操作水平突飞猛进,系统分类学开始了以基因或基因产物解析物种演化历史的新历程。DNA测序和PCR问世后,对基因进化速率的研究也几乎同时展开(Zinneretal.,2009; 刘海龙,2009; Wangetal.,2011)。由于线粒体没有内含子,较少受到重组影响,且为单倍型遗传模式,所以比细胞核基因更适合用作分类标准。线粒体12S和16S核糖体基因存在大量的插入和缺失现象,使序列比对陷入困境,制约了它们在分类中的应用。线粒体细胞色素c氧化酶亚基Ⅰ基因(COⅠ)比其他线粒体基因拥有更多的系统发育信号,COⅠ基因密码子的第3位核苷酸表现出很高的碱基置换率,这使得COⅠ基因在分子进化速率方面超出12S rDNA和16S rDNA两倍之多(Hebertetal.,2003)。COⅠ基因的氨基酸序列变化率要比cytb和任何其他线粒体基因更慢一些,这使得COⅠ基因能够提供更广阔的有关系统发生的视角。2003年,加拿大生物学家Hebert在总结前人工作的基础上,以加拿大37种蚊虫、北美260种鱼类、南美531种热带蝴蝶和87种蝙蝠为研究对象,提出以COⅠ基因为主要标准基因的DNA条形码技术。 (Hebertetal.,2003; Lecompteetal.,2008; Pereiraetal.,2010)。
目前DNA条形码研究的重点是界定种内种间遗传差异和物种之间的变异范围。在分析种内种间差异阈值时,往往出现样本量太少的情况,以致不能做出有效的统计分析,无法进行精确评估(陈春生等,2012;马英等,2012)。
鼠类是多种自然疫源性疾病的储存宿主。快速准确地鉴别鼠类,是防控疾病的关键。针对鼠类的传统形态学鉴定十分依赖于对鼠类的头骨、牙齿和外形的辨别,具有很强的专业性。本研究通过DNA条形码技术,对黑瞎子岛地区常见的2科3属4种41只鼠类的序列进行种内种间遗传距离的计算和聚类分析,以便积累鼠类条形码数据,建立COⅠ基因条形码数据库。
1 材料与方法
1.1 标本采集
所用实验样品为2011年在黑瞎子岛地区采集的鼠类,形态学鉴定为:棕背Clethrionomysrufocanus5只,红背C.rutilus8只,东方田鼠Microtusfortis28只,花鼠Eutamiassibiricus3只。现场鉴定后,分离肝脏和脾脏组织,保存于-80℃。从GenBank和BOLD systems下载用于分析的其他条形码序列,详见表1。

表1 实验样本信息Tab.1 Information of samples in this research
1.2 基因组DNA提取
取鼠肝脏或脾脏组织20 mg,研磨均匀。用北京天根生物公司基因组DNA提取试剂盒,提取组织DNA,并保存于-20℃。
1.3 PCR反应和序列测定
COⅠ基因扩增所使用的通用引物为COⅠ-L(5′-ACTTCTGGGTGTCCAAAGAATCA-3′)和COⅠ-H(5′-CCTACTCRGCCATTTTACCTATG-3′)(Robinsetal.,2007)。在50 μL反应体系中,Premix Taq(Takara)25 μL,上下游引物各1 μL(10 μmol/L),模板3 μL,去离子水20 μL。 PCR反应参数为:94℃预变性5 min; 94℃变性30 s,55℃退火40 s,72℃延伸1min,共35个循环;72℃延伸10 min。随后用1%的琼脂糖凝胶电泳检测PCR产物,并送北京华大基因测序。
1.4 COⅠ序列分析和系统树构建
用Chromos软件查看COⅠ基因序列峰图,判断峰图质量。用Clustal X软件进行多重序列比对。校正后的COⅠ序列在NCBI上用BLAST程序进行同源性比对。用MEGA5.0软件计算COⅠ序列的碱基组成。基于Kimura-2-parameter模型计算种内和种间遗传距离,以邻接法(Neighbour-joining,NJ)构建系统进化树。最大简约法分析使用启发式搜索,参数如下:1 000 逐步加入法随机加入序列构树方法采用二等分再连接(Tree-bisection reconnection,TBR),可靠性由1 000次自展分支检验。邻接法分析采用HKY85遗传距离,自展检验1 000次。
2 结果
2.1 41个鼠类标本COⅠ基因扩增结果
通过通用引物成功扩增出41个鼠类标本的COⅠ基因片段,大小约为750 bp。电泳结果未发现非特异性杂带。通过测序,进一步验证了扩增结果。
2.2 COⅠ基因的BLAST比对结果
将41个鼠类样本COⅠ基因序列同NCBI上鼠类条形码序列进行同源性比对,结果显示,5只红背被误判为棕背,24只莫氏田鼠被误判为东方田鼠,1只大林姬鼠被误判为东方田鼠。其余样本形态学鉴定结果和DNA条形码比对结果一致。
2.3 种内与种间遗传距离
用MEGA5.0软件,基于Kimura-2-parameter模型计算黑瞎子岛地区41个鼠类标本的平均种内和种间遗传距离,见表2。
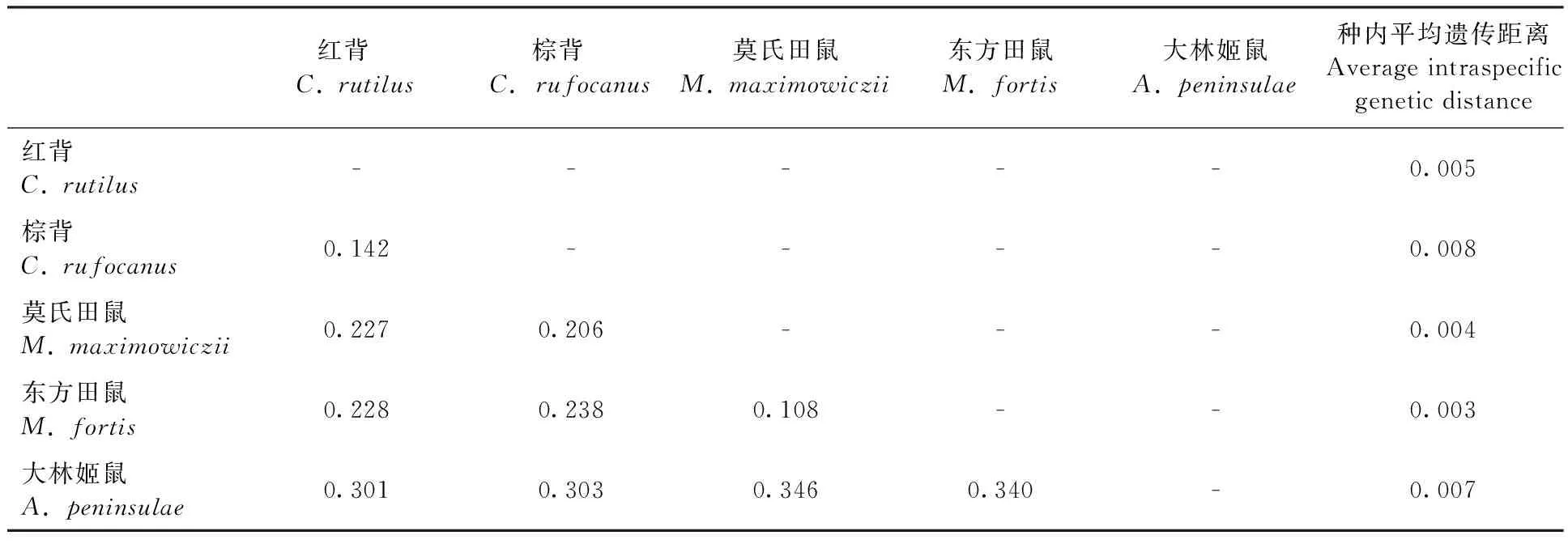
表2 黑瞎子岛地区鼠类两两比对的种间平均遗传距离和种内遗传距离Tab.2 Average intraspecific genetic distance and average interspecific genetic distance
注:棕背样本数据来自BOLD systems数据库
Note: TheC.rufocanussample date comes from BOLD systems
2.4 系统发育树构建
邻接法构建的系统发育树可以反映序列之间的真实距离。本研究利用黑瞎子岛地区4种鼠类共41个COⅠ基因序列,连同BOLD systems和GenBank中的鼠类序列,用NJ法构建系统发育树,结果如图1。聚类分析结果表明,黑瞎子岛地区常见4种鼠类的同种个体聚为单独的一支,同属不同种的个体聚为更大的一支,同科不同属的分支聚为一总支。
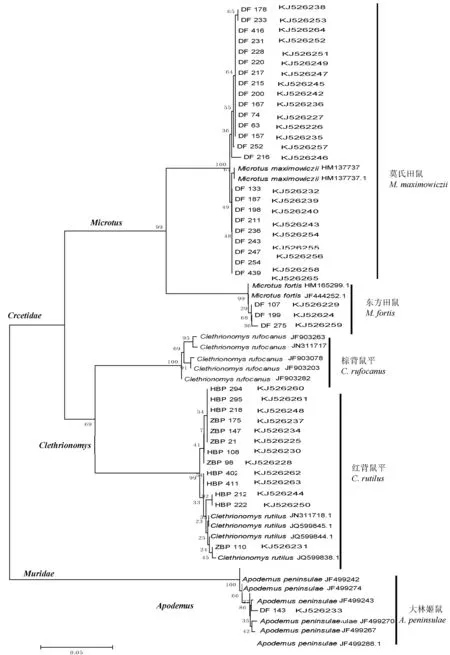
图1 NJ法构建的黑瞎子岛地区常见鼠类分子系统树Fig.1 NJ tree based on COⅠ gene fragments from 41 individuals of 4 rat species
3 讨论
黑瞎子岛地区41个鼠类样本COⅠ基因序列在NCBI数据库进行BLAST比对后,结果显示,相似性在99%~100%。在对鼠类样本进行PCR扩增时,花鼠样本未能出现特异性条带,推测原因可能是扩增体系不完全合适,以后研究中将加以改善。
基于COⅠ基因DNA条形码技术的检验标准之一是种间差异远大于种内差异。Hebert等(2003)对GenBank中13 320个亲缘关系很近的同属物种的COⅠ序列进行了分析,结果表明种内差异大多小于1%,极少大于2%,而种间差异则高达11.3%。本研究中通过对黑瞎子岛地区2科3属4种41个鼠类个体COⅠ基因序列进行分析,同一种鼠类不同个体的DNA条形码序列差异很小,为0.003~0.008,平均为0.005;种间差异则很大,为0.108~0.346,平均为0.258。平均种间遗传距离是种内遗传距离的51倍,完全符合Hebert所作出的关于DNA条形码有效性的检验标准。
DNA条形码技术鉴定物种时,种间遗传距离和种内遗传距离之间的分离程度十分重要,有效的鉴定意味着种间遗传距离和种内遗传距离没有重叠(Aliabadianetal.,2009)。本研究对黑瞎子岛地区常见的4种41个鼠类标本的条形码序列进行了分析,结果表明4种鼠类的种内遗传距离和种间遗传距离没有重叠区域,这些数据提示DNA条形码为有效区分鼠类提供了可能的手段。
构建系统发育树进行分子鉴定是非常直观的方法。NJP法构建的系统发育树聚类分析结果表明,不同种类的鼠位于不同的分支,不同种属的鼠类区别明显。每一个单系分支对应一个特异的物种,节点支持率为100%。在分支长度上,种间分支较长,种内分支则很短。本研究以实验所得的41个COⅠ序列和从NCBI和BOLD systems中得到的14个相关鼠类COⅠ序列构建系统发育树,每种鼠类形成的独立分支进一步验证了DNA条形码鉴定鼠类的有效性。
本研究以黑瞎子岛地区41个鼠类个体为样本,扩增COⅠ基因,结合Genbank和BOLD systems中相关的鼠类COⅠ序列,计算种内种间遗传距离,并进行序列聚类和系统发育分析,证明基于COⅠ基因的DNA条形码技术不仅能够方便、快捷、准确的鉴定鼠类,纠正形态学鉴定中的失误,而且还能很好的鉴别形态相近的近缘种。DNA条形码技术必将在口岸检验检疫、保护农业生产安全、保护生态安全方面发挥重大的作用。
