沉积硅藻揭示的历史时期水生植被信息——以梁子湖为例
2014-11-05张清慧董旭辉陈诗越羊向东
张清慧 董旭辉 姚 敏 陈诗越 羊向东
(1. 中国科学院南京地理与湖泊研究所, 湖泊与环境国家重点实验室, 南京 210008; 2. 聊城大学环境与规划学院,聊城 252059)
长江中下游地区是我国淡水湖泊分布最集中的地区。据统计, 全国面积大于1 km2的淡水湖泊中约40%分布在该地区, 大于 10 km2的淡水湖泊中有63%分布在该地区, 且大多数都是浅水湖泊, 具有面积大、水浅、换水周期较短及人类活动干扰强度大等特点[1]。在历史上, 该区湖泊水生植物分布广且种类丰富, 是此类湖泊生态系统的重要组分。20世纪中叶开始的大规模围湖造田和 80年代兴起的围网养殖, 破坏了湖泊水生生物的生境, 其数量、群落构成发生了显著变化; 大量农业肥料的使用, 城乡居民生活垃圾等营养物质的输入使大多数湖泊逐渐富营养化, 浮游植物异常增殖, 水生植被急剧萎缩,湖泊生态系统退化严重。据2001—2010年间沿江近100个湖泊的多次水质监测, 该区目前近 80%的湖泊已处于富营养, 甚至重富营养状态[1]。
这种浅水湖泊中由于水体富营养化引起的水生植被特别是沉水植被的衰退和消失也是世界范围内的普遍现象[2]。水生植物作为浅水湖泊生态系统重要的初级生产者之一, 对稳定湖泊生态系统的结构和功能起着重要作用[3]。首先, 作为湖泊的主要初级生产者, 为其他生物提供食物来源、栖息地与产卵场所; 其次, 作为湖泊水体净化者, 能有效地降低水体中的营养盐和污染物浓度, 改善水体溶氧状况、提高水体透明度。通常, 根据湖泊中水生植被和浮游藻类的多寡, 湖泊可划分为“草型”和“藻型”两种稳定的生态状态类型。湖泊生态修复就是把蓝藻水华频发、水质浑浊的富营养化藻型湖泊生态系统通过一定的途径转化为水生植物茂盛、水质清澈的草型湖泊生态系统[4]。因此, 水生植物群落的构成是健康浅水湖泊生态系统最为重要的特征之一。
影响水生植物生长和分布的环境因素众多, 如水位、透明度、水质、其他生物组分[5,6], 营养盐浓度、鱼类群落、自然条件(如飓风事件等)等均可能触发草型、藻型湖泊的相互转变[3,4]。然而, 湖泊水生植物群落是生态系统长期演化的产物, 其形成及演化机制必须从历史的视角加以考察。特别是现有的湖泊生态监测多始于环境恶化之后, 记录时间也相对较短(一般<50年), 并且在强烈人类活动干扰下,现今大多数湖泊水生植物群落都发生了显著变化[7]。因此, 基于现有湖泊的生态调查, 往往不足以提供不同状况下湖泊的水生植物群落演替过程和特征,以及生态系统转型的信息。另外, 在考虑利用水生植被进行湖泊生态修复时, 由于缺乏对湖泊自然过程的了解, 所选择的植被是否曾经在湖泊中出现过,出现时是否有助于改善水体生态状况等信息, 也无法从短期的监测记录中了解到。在这种情况下, 很难避免生态修复过程本身对湖泊生态系统结构造成新的扰动, 达不到有序地恢复湖泊过去的生态群落的目的[8], 甚至可能造成新的物种入侵(例如水葫芦引种)。因此, 了解历史时期水生植物群落的变化可以提供湖泊长期生态环境演化过程信息, 并为浅水湖泊生态修复提供理论支撑[9]。
湖泊沉积物包含了丰富的生物和理化方面的信息, 是过去环境变化的窗口, 可以用来重建湖泊生态系统及其流域过去变化的历史[10]。在普遍缺乏长期监测记录的情况下, 基于湖泊沉积物研究的古湖沼学, 特别是古生态学技术是唯一能获得历史环境演变的有效手段。近年来, 在古湖沼学研究中, 许多生物指标(包括硅藻[11—13]、摇蚊[14]、孢粉[15]以及植物化石[16]等)已被广泛地用来揭示水环境变化(包括营养、水文和水生植被等)的综合信息。目前, 在长江中下游平原湖泊开展的前期工作[11,12]表明, 硅藻敏感地响应于环境变化, 能很好地指示湖泊营养、水文和水生植被等特征, 是古湖沼学研究使用最多的生物指标之一[13]。例如董旭辉等通过对该区 45个湖泊表层沉积硅藻与湖泊环境数据进行分析揭示了部分附生硅藻(如 Achnanthes minutissima、Cocconeis placentula、Nitzschia palea 等)对水生植被数量有明显的指示意义[11]; Yang等对典型湖泊(龙感湖和太白湖)沉积柱中硅藻的研究表明: 在太白湖,沉积硅藻清楚地记录了湖泊由草型向藻型阶段转变的过程; 在龙感湖, 沉积硅藻则更多的体现出草型湖的特征[17]。但由于缺乏历史水生植被的直接证据,这些结论至今尚未得到证实。
本研究是以长江中下游地区一典型草型湖泊梁子湖为研究对象, 结合高分辨率测年技术, 利用沉积硅藻重建梁子湖历史时期湖泊水生植被的演替过程, 并利用梁子湖水生植被的长期监测记录作为历史水生植被的直接证据对重建结果进行验证, 试图证实沉积硅藻记录能否揭示过去水生植被的演化过程; 利用历史时期流域环境信息, 揭示梁子湖水生植被演替的驱动因素, 为梁子湖水生植被保护、长江中下游浅水湖泊水生植被恢复提供理论依据。
1 研究区概况
梁子湖(30°05′-30°18′N, 114°21′-114°39′E)是湖北第二大湖, 位于湖北东部长江南岸, 垮武昌县、鄂州市境, 与保安湖、三山湖和鸭儿湖紧邻。湖泊面积304.3 km2, 平均水深4.2 m, 最大水深6.2 m[18]。气候属于北亚热带季风气候, 年均气温 16.8℃, 极端最高与最低气温分别是 40.7℃和–12.9℃, 年无霜期为 275d, 年均降水量为 1263.4 mm。洪涝灾害时有发生, 多年平均水位 17.8 m, 历年最高水位27.8 m (1954年), 最低水位16.0 m (1992年)[18]。湖区水较浅、湖盆平坦、底泥肥沃, 水生植物种类繁多且覆盖面积大, 并且保持着丰富的生物多样性,为典型草型湖, 是世界自然保护联盟(IUCN)、国际鸟类保护联合会(ICBP)和国际重要湿地名录确定的重要湖沼湿地。
目前梁子湖中沉水植被群落占绝对优势, 主要种类有眼子菜科的种类: 以微齿眼子菜(Potamogeton mackianus)和菹草(P. crispus)为主、金鱼藻(Ceratophyllum demersum)、穗状狐尾藻(Myriophyllum spicatum)、苦草(Vallisneria spiralis)、竹叶眼子菜(P.malaianus)和黑藻(Hydrilla verticillata)等; 浮叶植物群落被次之, 主要种类有四角菱(Trapa quadrispinosa)、莲(Nelumbo nucifera)、芡实(Euryale ferox)、荇菜(Nymphoides peltatum)和萍蓬草(Nuphar pumilum)等;挺水植被主要种类是菰(Zizania caduciflora), 局部地区有芦苇(Phragmites communis)、菖蒲(Acorus calamus)等, 但未形成群落[19,20]。在空间上, 梁子湖水生植被分布差异大, 东梁子湖为草型湖, 水生植被发育良好,覆盖率为 95%左右; 前江大湖植被较少; 中湖基本上无水生植被[18]。在时间上, 该湖水生植被可能经历了复杂的演变过程。根据1951—2001年间多次调查数据显示, 期间水生植被经历了恢复(1950s)→旺盛顶期(1980s)→急剧衰退(1990s)→逐步恢复(2000s)的几个阶段[21]。2010年 8月梁子湖又遭受了一次灭顶之灾,持续的湖泊高水位导致>80%的水生植物死亡(新华社2010年8月14日报道)。
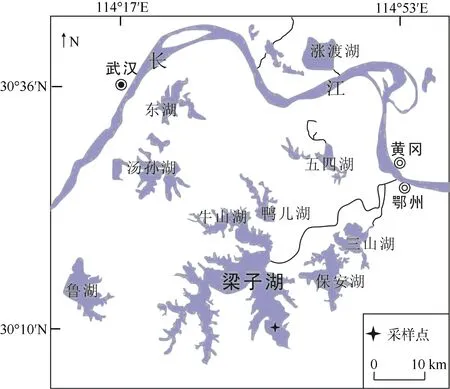
图1 梁子湖及采样点位置Fig. 1 Sampling sites of of Liangzi Lake
2 研究方法
2.1 样品采集及实验室分析
2011 年于东梁子湖(30°10′N, 114°37′E, 水深2.7 m)利用重力采样器采集了长85 cm的柱状沉积岩芯。沉积岩芯现场分样, 每个子样品厚度1 cm, 样品密封带回实验室后<4℃保存以备分析测试。210Pb和137Cs年代测试采用美国EG&GOrtec公司生产的高纯锗井型探测器(HPGe GwL-120-15)在中科院南京地理与湖泊研究所进行测定。采用Constant Rate of Supply (CRS)模式结合1963年峰值时标的复合模式方法[22]来获得梁子湖年代框架。沉积硅藻取约0.5 g沉积物, 先用 10%盐酸去钙质胶结物, 再用双氧水去有机质, 离心清洗制片。硅藻种属鉴定参照Krammer和 Lange-Bertalot的分类系统[23], 由于210Pb测年范围的局限性, 只对63 cm以上的样品进行分析。依据硅藻保存情况, 每个样品的硅藻统计数控制在 300粒以上, 属种丰度按百分比表示, 对硅藻序列不同演化阶段的划分利用Tilia-CONISS软件进行[24]。
2.2 历史时期水生植被信息的重建与对比
利用沉积附生硅藻重建梁子湖历史时期湖泊水生植被的演替过程, 应用该湖水生植被的长期监测数据对重建结果进行验证, 结果表明该湖沉积附生硅藻记录对历史时期水生植被的演替有较好的反映。基于此, 对梁子湖过去近 200年的水生植被覆盖度进行了重建, 考虑到沉积物的分辨率和沉积时存在一定的混合, 对此重建结果用 Loess平滑得到过去近200年长期的水生植被覆盖度曲线。
2.3 流域资料数据
湖北省历年洪涝灾害等级数据来源于文章《基于水位和雨量的洪涝受灾面积评估模型研究》[25]。文中指出, 根据洪涝等级指数的范围定义洪涝灾害等级, I>1.0 时为严重洪涝; 1.0≥I>0 时为较重洪涝;0≥I>–0.5时为一般洪涝; I≤–0.5 时为旱重于涝。
3 结果
3.1 沉积年代的确定
梁子湖沉积钻孔中在23 cm处出现一137Cs峰值,对应于1963年的全球核试验。考虑到长江中下游地区强烈人类活动的影响, 采用210Pb CRS模式的复合模式[22], 即用137Cs时标对CRS计算结果进行校正的模式对钻孔沉积物进行年代计算, 以此获得真实的钻孔年代标尺。历史时期沉积岩芯年代与深度对应关系如图2所示。通过CRS模式计算的沉积岩芯的平均沉积速率为0.034 g/(cm2·a)。沉积岩芯63 cm处年代约为1850AD。
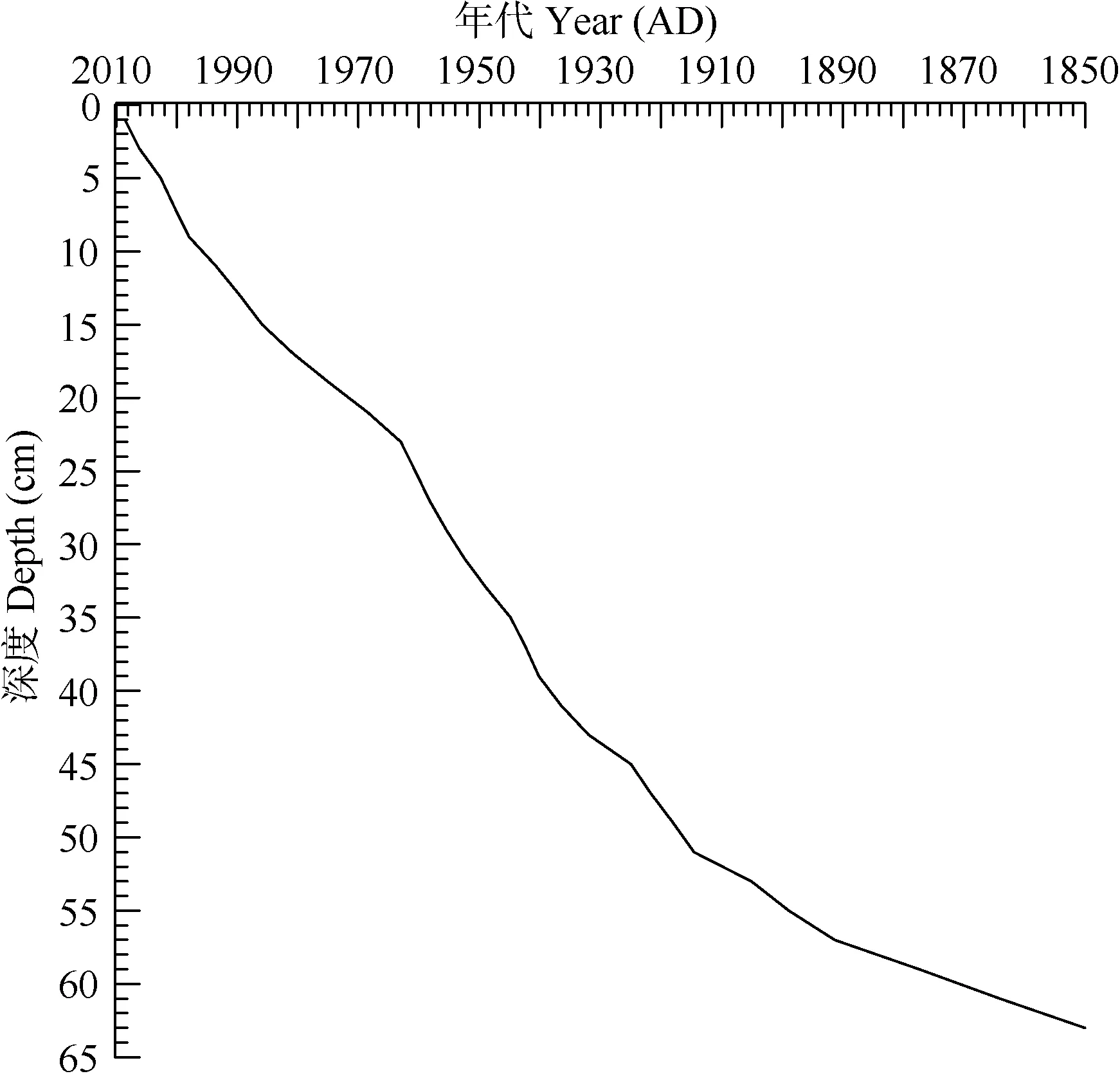
图2 梁子湖年代-深度对应曲线Fig. 2 The relationship between chronology and depth in Liangzi Lake
3.2 沉积硅藻组合
梁子湖钻孔沉积物共鉴定出硅藻28属170种,包括附生、浮游和底栖类型(图3)。其中, 附生种类总含量很高, 最高总含量达 73%, 且属种丰富, 包括 Achnanthes minutissima、Fragilaria capucina、Cymbella spp.、Eunotia spp.、Gomphonema spp.、Gyrosigma acuminatum等。浮游种包括Aulacoseira granulata、A. ambigua、A. alpigae、Cyclotella pseudostelligera, C. meneghiniana等。底栖种以 Navicula和 Nitzschia的类型以及 Gyrosigma acuminatum为主。此外, 兼附生浮游的F. ulna也常以伴生种出现。根据硅藻丰度变化可划分为 4个组合带 2个亚带,硅藻组合特征自下而上叙述如下:
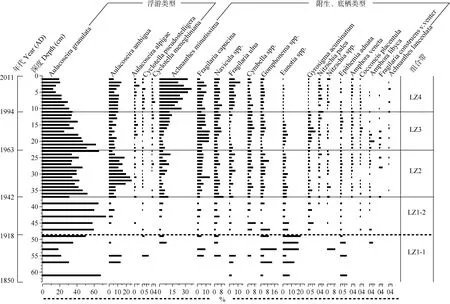
图3 梁子湖沉积柱主要的硅藻种群(图中仅列举含量高于2%的属种)演替Fig. 3 The major diatoms(species with≥2% in at least one sample) in the sediment core from Liangzi Lake
LZ1 带(63—37 cm; 1850—1942AD)总体特征是以浮游种占绝对优势, 尽管附生硅藻总含量不低,但各个附生硅藻种类含量均不高, 不能成为优势种组合。其中LZ1-1带(63—49 cm; 1850—1918AD)主要以浮游种A. granulata占优势, 最高含量达70%以上, 但波动很大。附生种类经常出现且波动较大, 如Eunotia spp.、Gomphonema spp.、A. minutissima、F.capucina等。底栖种 Navicula spp.与兼附生浮游的F. ulna偶有出现且含量极低。LZ1-2带(49—37 cm;1918—1942AD)浮游种 A. granulata含量明显增加,占绝对优势。附生种类较上一带含量减少, 且各个附生硅藻种类含量均相对不高(<10%)。底栖种Navicula spp.仍保持一定含量, G. acuminatum在此带开始出现。
LZ2带(37—23 cm; 1942—1963AD)仍以浮游种A. granulata为优势种, 与其同属的A. ambigua含量含量较上一带含量增加, A. granulata与A. ambigua互为消长关系。附生类型A. minutissima、F. capucina等亦有所增加。底栖种 Navicula spp.含量略增,Nitzschia的类型在此带少量出现。兼附生浮游种F.ulna含量略增。
LZ3带(23—11 cm; 1963—1994AD)浮游种 A.granulata呈减少趋势, A. ambigua含量也明显减少。较上一带各附生硅藻含量进一步增加, 其中以A. minutissima、F. capucina的增加最为明显。其中,F. capucina在 1972—1981年期间达到剖面的最大值。底栖种在此带呈增加趋势, 其中 G. acuminatum在此带最为丰富, 并在17 cm(1981AD)处达到剖面最大值。兼附生浮游种F. ulna含量迅速减少甚至消失。
LZ4带(11—0 cm; 1994—2011AD)附生种 A.minutissima呈明显增加趋势, 最高含量达 37%, 成为主要的优势种, 而浮游种A. granulata含量迅速下降且呈减少趋势, 为次优势种, 其他附生种如 F.capucina, Cymbella spp., Gomphonema spp.无明显变化, 仍保持着一定含量, 而Eunotia spp.含量减少甚至消失。底栖种Navicula spp.无明显变化, Nitzschia的类型含量略增, 而G. acuminatum含量明显减少。兼附生浮游种F. ulna在此带含量显著增加。
3.3 梁子湖历史时期水生植被信息的重建及验证
如图 4所示, 梁子湖沉积柱附生硅藻变化趋势与记载的水生植被不同的发展期的对比结果显示:1950s附生硅藻呈增加趋势, 对应于历史记录的水生植被恢复期; 1970s附生硅藻含量显著增加, 到80年代初到顶峰值, 为水生植被显著上升期; 1990s附生硅藻含量较80年代明显减少, 为水生植被的衰退期; 2000s附生硅藻又呈增加趋势, 为水生植被的逐步恢复期。水生植被不同的发展期与洪涝灾害等级对应结果显示: 水生植被的衰退期对应于高水位的特大洪涝年, 而恢复期、显著上升期和逐步恢复期对应于相对较低水位的洪涝灾害等级年。
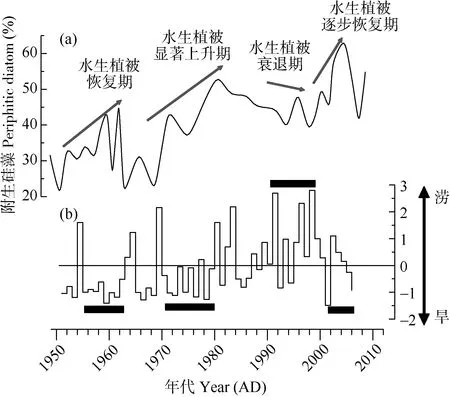
图4 梁子湖沉积柱附生硅藻变化趋势与记载的水生植被不同的发展期的对比图(a)和湖北省历年洪涝灾害等级图(b)Fig. 4 The periphitic diatom communities in sediment cores from Liangzi Lake and the aquatic vegetation growth period from history records (a), and flood changes (b) over the past 60 years in Hubei Province
对梁子湖过去近200年的水生植被覆盖度重建的结果如图 5所示: 水生植被覆盖度自1850AD以来呈快速增加趋势并在 1905AD出现一峰值, 之后逐渐下降至 1935AD左右出现一低值, 之后开始增加, 60年代后呈显著增加趋势, 80年代初期达到顶峰, 而 1990s水生植被覆盖度较 80年代减少, 至2000s又呈显著增加趋势。
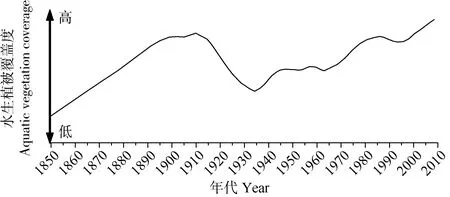
图5 梁子湖水生植被覆盖度变化Fig. 5 The aquatic vegetation coverage in Liangzi Lake
4 讨论
沉积硅藻记录可以揭示梁子湖历史时期水生植被覆盖度的变化。在古湖沼学研究中, 许多其他沉积生物指标(如植物化石、孢粉等)也能记录部分历史湖泊植物群落的信息。例如 Sayer等利用沉积物中植物化石分析, 并结合该湖孢粉和硅藻等指标研究, 重建了该湖水生植被与浮游植物群落组成、生长季节在年际上的变化过程[26]。通常, 植物大化石对湖泊植被具有直接指示意义, 可以为湖泊水生植被演替提供直接证据[27,28]。但由于一些湖泊水生植物化石保存性差, 单独利用植物大化石来推断历史时期湖泊水生植被存在一定缺陷。孢粉也是重建历史时期湖泊植被的一种有效手段, 但主要用于陆生植被的重建[29,30], 且水生植物花粉产量相对较低,保存性较差[31], 因此重建的结果往往具有片面性。然而, 与植物化石和孢粉相比, 硅藻在水生植被重建方面有着独特的优势。通常, 附生硅藻种类优势组合指示了水生植被的发育状况良好[32]。梁子湖历史记录也很好地验证了利用沉积硅藻推导的历史水生植被演替情景。根据1951—2001年间多次调查数据显示、期间水生植被经历了恢复(1950s)→旺盛顶期(1980s)→急剧衰退(1990s)→逐步恢复(2000s)的几个阶段[21,33,34], 与沉积附生硅藻记录基本一致(图 4a), 表明附生硅藻对历史时期水生植被的覆盖度有较好的反映。
沉积硅藻记录还可以揭示不同水生植物类型的变化。通过对长江中下游湖泊水生植物与其附生硅藻关系的研究发现(未发表数据), 各类附生硅藻对水生植物有明显的选择性, 由此进一步探讨了附生硅藻种类指示不同水生植物类型(沉水植物、浮叶植物、漂浮植物和挺水植物)的可行性。其中, A.minutissima、F. capucina、Gomphonema在沉水植物上出现的含量最高, 指示了其对沉水植物的选择性较为明显; Cymbella和Eunotia的种类对浮叶植物的选择性非常明显; G. acuminatum则在挺水植物及沿岸湿生植被上最为常见, 它的出现可以指示水生植物的增多或水体透明度的增加。如图 3所示, A.minutissima、F. capucina、Gomphonema在钻孔中一直存在且为含量相对较高的优势种表明沉水植被是钻孔点附近水域主要的水生植被。1972AD前, A.minutissima含量较少, 之后含量大幅度升高, 成为主要优势种且在表层含量达到最高, 其含量变化表现了梁子湖沉水植物由少到多的发育过程。1972—1981AD, F. capucina和G. acuminatum在此阶段含量最高, 指示了在这一阶段水生植物最为发育, 分别指示沉水植物和挺水植物的发育。另外, G. acuminatum的较高含量也指示了此阶段水生植物的大量发育和水体透明度的增加。这也与历史记录中80年代水生植被的显著上升期一致。1990s后, Eunotia和 G. acuminatum含量减少甚至消失, 其含量变化分别指示了浮叶植物和挺水植物及沿岸湿生植物的减少。因此, 从群落演替来看, 历史时期梁子湖以沉水植被为主且呈增加趋势, 而浮叶植物和挺水植物群落减少。这与1951—2001年间多次调查数据结果比较一致。据1951—2001年间多次调查数据结果表明, 梁子湖水生植被的优势群落生活型未发生变化,仍以沉水植物群落为主, 挺水植被中芦苇群落已消亡, 浮叶群落分布面积也变小[21]。因此, 沉积硅藻记录对历史时期水生植被不同生活型的变化也有较好地反映。
影响水生植物生长和分布的环境因素众多, 如水质、水位、透明度、其他生物组分等。近年来大量研究表明洪涝所导致水位变化是影响水生植被发育的重要因子[35]。当洪水引起的水位快速上升超过沉水植物发育的水深阈值时, 会抑制沉水植物的生长; 而洪水也会引起水体浑浊度的迅速增加, 从而影响光照和光透度, 进而使得植物的水下光合作用受阻[36]。特别是在过水性浅水湖泊, 洪水是导致水生植被突然减少的重要因素。受洪水的影响, 湖泊高水位持续时间长、湖水透明度降低、水生植被不能进行有效的光合作用, 从而导致代谢紊乱, 再加上水流造成的物理损伤、病害、被取食等原因, 使水生植被大量死亡[36,37]。据资料显示: 1998年的特大洪水导致梁子湖水生植物覆盖率减少了 50%, 水质由二类退化为四类[38]; 2010年8月份该湖持续的高水位导致>80%的水生植物死亡(新华社2010年8月14日报道)。另外, 在Okeechobee 湖28 年的观测资料显示, 沉水植物生物量与高水位呈负相关,与透明度呈正相关[39]。由此可见, 洪水是影响梁子湖水生植被发育的重要因素之一, 有效的水生植被的修复, 必须避免湖泊出现连续的高水位时期, 同时还需要保持较高的水体透明度。值得指出的是, 20世纪50年代以来, 随着的人类活动增强, 由此引起的流域土壤侵蚀速率加快和入湖营养盐的增多, 尤其是磷的增加, 促进了藻类生物的发育和营养的富集, 一定程度上抑制了水生植物的发育。据资料显示, 目前梁子湖营养状态已从贫营养型进入中营养型, 局部已接近富营养型, 主要是氮磷、超标[40]。显然, 营养富集势必对水生植被的发育产生影响,具体反馈机制仍需进一步分析。
尽管本研究证实了沉积硅藻在反演历史时期水生植被信息上的可靠性, 但同其他古生态指标一样,硅藻的保存性在一定程度上会影响水生植被重建结果的精度。硅藻在埋藏过程中会受外力的作用(如风力、水力及动物携带等)的影响进行搬运和扩散, 其坚硬的硅质壳体也会受到不同程度的磨损, 影响到其保存完整性。例如, 有研究表明较强的水动力环境不利于沉积硅藻的保存[41]。在历史上, 长江中下游地区大多数湖泊湖盆较浅平, 与长江联系密切自然相通, 保持着较高的水体交换能力, 水动力强度大[1,18]。根据作者所在团队对长江中下游地区湖泊(如涨渡湖、巢湖、太白湖、龙感湖等)大量研究发现, 硅藻的保存性随沉积年代的增加而变差。在本研究中, 梁子湖沉积硅藻50 cm以下深度的硅藻保存性逐渐变差, 因此基于该深度以下沉积硅藻的水生植被信息的反演可能存在误差, 需谨慎解释。
5 结论
该研究利用古湖沼学手段, 在建立高分辨率沉积物年代的基础上, 利用沉积附生硅藻的演化序列对梁子湖历史时期该湖水生植被的覆盖度进行了重建, 研究发现: (1)沉积硅藻所揭示的历史时期湖泊水生植被信息与该湖过去 50余年来监测所记录的水生植被演化历史一致, 是一种重建历史时期湖泊水生植被信息的有效生物指标。(2)沉积硅藻记录还可以揭示不同水生植物类型的变化。过去近200年来梁子湖以沉水植被为主且呈不断增加趋势, 而浮叶植物和挺水植物群落减少。(3)结合湖泊流域历史环境数据, 揭示了洪水是影响该湖水生植被发育的一个重要因素。因此, 有效的水生植被的修复, 必须避免湖泊出现连续的高水位时期。本研究结果不仅为梁子湖水生植被的恢复提供了科学依据, 也为该区类似浅水湖泊富营养化治理提供了重要的生态学依据。
