南海北部冷泉碳酸盐岩中钙质生物壳体和自生碳酸盐矿物的Mg/Ca、Sr/Ca及其环境指示
2014-10-27杨克红初凤友朱继浩马维林董彦辉
杨克红,初凤友,朱继浩,马维林,董彦辉
(1.国家海洋局 海底科学重点实验室,浙江 杭州310012;2.国家海洋局 第二海洋研究所,浙江 杭州310012)
1 引言
碳酸盐岩在古冷泉和现代冷泉环境中沉淀已经广为认知[1]。在硫酸盐-甲烷转换带附近,在甲烷氧化古细菌和硫酸盐还原细菌共同作用下,大量的甲烷被氧化,硫酸根离子被还原,导致环境碱度增大从而沉淀自生碳酸盐矿物[2]。冷泉碳酸盐岩是甲烷渗漏系统的重要产物,记录了甲烷渗漏活动和流体组分随时间的变化过程,是保存甲烷渗漏系统信息的最好档案[1,3]。
冷泉碳酸盐岩的主要矿物为自生碳酸盐矿物,如方解石(高镁方解石和低镁方解石)、文石、白云石等,还可见菱铁矿、草莓状黄铁矿、石英、长石、黏土等,同时也可见丰富的钙质生物,如有孔虫、颗石(coccolith)等[4—8]。碳酸盐岩的成岩作用过程除了岩石学观察外,还可以通过地球化学分布模式来进行研究。和C、O同位素一样,Sr和Mg的地球化学特征也是示踪碳酸盐岩成岩条件的主要手段[9]。事实上,碳酸盐岩的Sr/Ca是古海洋环境变化和碳酸盐重结晶相互作用的结果。ODP154航次5个站位全岩方解石的Sr/Ca和Mg/Ca的结果与年龄变化比与沉积深度变化更为吻合,表明它们代表了海洋环境的Sr/Ca和Mg/Ca,而不是由深度控制的成岩作用,但同时也指出全岩方解石可能不如有孔虫方解石能够显示受钙化深度控制的初始Sr/Ca,因为它主要受控于透光层的方解石沉积[10]。卡斯卡地亚大陆边缘现代和古冷泉碳酸盐矿物研究表明,随着成岩作用的推进,Mg/Ca和Sr/Ca呈降低趋势[11]。可见,Mg/Ca和Sr/Ca能够揭示一些碳酸盐岩形成时的环境及成岩信息。
近年来数个航次在南海北部陆坡发现了指示甲烷渗漏的自生碳酸盐岩[4,12—16],它们不仅呈现胶结块状、烟囱状、环状、椭球状、角砾状等不同于正常海相碳酸盐岩的形貌特征,并且具有较负的δ13C(变化范围约为-5.2‰PDB~-56.88‰PDB,多数小于-30‰PDB)和轻微富集的δ18O(变化范围约为0.42‰PDB~5.28‰PDB,多数大于3‰PDB),而典型海相碳酸盐岩的δ13C值变化范围约为-3‰PDB~5‰PDB,δ18O值变化范围约为-1‰SMOW~3‰SMOW①该文献中PDB标准和SMOW标准之间的转换公式为:δ18 OSMOW=1.030 86δ18 OPDB+30.86‰。[17],碳氧同位素及岩石学、矿物学等信息证实了这些自生碳酸盐岩是由于甲烷厌氧氧化作用沉淀的冷泉碳酸盐岩。因此,本文尝试通过对南海北部冷泉碳酸盐岩中的自生碳酸盐矿物及钙质生物壳体的Mg/Ca和Sr/Ca进行分析,来揭示它们反映的古环境及成岩作用。
2 地质背景
南海处于欧亚板块、太平洋板块和印度板块的汇聚带,呈北东——南西向的菱形,北部陆缘属于华南地块的一部分,为张裂性被动大陆边缘,西部为印支地块,东靠台湾-菲律宾岛弧。南海北部地质构造受三大板块互相运动所制约,形成了断裂地块并沉积了丰富的有机质[13],具有一系列的含油气盆地,常规油气资源十分丰富,研究区及附近呈NE-SW延伸的断裂构造发育,是天然气向海底渗漏的有利通道,具有形成海底天然气水合物的良好成矿环境。
3 样品和方法
样品为2006年国家海洋局第二海洋研究所利用“大洋一号”执行环境调查航次时利用电视抓斗获取的冷泉碳酸盐岩,获得冷泉碳酸盐岩的站位距离九龙甲烷礁约10~12 km(表1)。

表1 碳酸盐岩站位信息Tab.1 Carbonate sample locations
碳酸盐岩获取后直接用海水冲洗干净,然后放在冷库中。在研究中,对选取的碳酸盐岩样品首先在超声波中用去离子水清洗,尽可能地清除孔隙中的一些残留沉积物。清洗后的样品装入样品袋中,然后在冷冻干燥仪中进行冷冻干燥,对干燥后的样品进行薄片制作。
研究采用岩石学镜下观察及电子探针分析的方法,均在国家海洋局海底科学重点实验室进行。电子探针仪器主机型号为JXA-8100,能谱仪为Oxford INCA X-sight,分析加速电压15 k V,束流20 n A,束斑15~20μm;修正方法ZAF;标准样品为美国SPI公司53种矿物。
4 结果
4.1 岩石学特征
岩石学观察显示南海北部冷泉碳酸盐岩为较均匀的微晶结构(见图1a、b),未见明显的成岩重结晶作用,含有包裹结构(见图1a、c)和孔洞结构(见图1b)。碳酸盐岩中包裹结构分布不均匀,包裹体最大的可达1 mm以上,最小的大概为200μm。孔洞结构周围的碳酸盐颗粒较大,结晶程度高。碳酸盐岩中生物遗迹较为丰富(见图1c、d),包裹结构中各种钙质生物尤其丰富(见图1c)。
4.2 Sr/Ca和 Mg/Ca特征
在进行电子探针测试时,根据自生碳酸盐矿物和钙质生物壳体在岩石结构中的位置进行了分类,在包裹结构内分布的自生碳酸盐矿物和钙质生物壳体分别称之为包裹的碳酸盐矿物和包裹的钙质生物壳体(具体位置见图1c和图2a~c);其余的自生碳酸盐矿物和钙质生物壳体分别称之为基质中的碳酸盐矿物和基质中的钙质生物壳体(图1d和图2d~h)。

图1 冷泉碳酸盐岩的岩石学特征Fig.1 Petrology of the seep carbonates
对自生碳酸盐矿物和钙质生物壳体进行了Ca、Mg、Sr、Ba的电子探针分析,计算结果显示(表2):自生碳酸盐矿物的 Mg/Ca变化范围为0.50~39.19 mmol/mol,平均为12.50 mmol/mol,Sr/Ca变化范围为0.06~2.90 mmol/mol,平均为0.53 mmol/mol;钙质生物壳体的 Mg/Ca变化范围为1.14~84.57 mmol/mol,平均为24.57 mmol/mol,Sr/Ca变化范围为0.77~1.84 mmol/mol,平均为1.08 mmol/mol。
依据Mg/Ca和Sr/Ca的关系可以看出(见图3),除去个别测试样品,不论是包裹的还是基质自生碳酸盐矿物均具有低到中等的Mg/Ca、低Sr/Ca的特征,而钙质生物壳体却根据其在岩石中的结构位置呈现明显的分组现象,基质钙质生物壳体具有高Mg/Ca、中等Sr/Ca的特征,包裹的钙质生物壳体具有低Mg/Ca、中等Sr/Ca的特征。
5 讨论
5.1 Mg/Ca、Sr/Ca的矿物学指示
Ca、Mg、Sr是性质相似的碱土金属,化学性质非常活跃,它们的阳离子易与CO2-3结合形成碳酸盐,以Ca为主要阳离子的方解石、文石和以Mg、Ca为主要阳离子的镁方解石、白云石等是冷泉碳酸盐岩的重要组成部分,而Sr元素和Ca元素具有相似的离子电荷,主要以置换Ca离子的形式存在于文石和方解石中,尤其是文石中[18-20]。前人研究结果表明,文石具有较高的Sr/Ca和较低的Mg/Ca,而高镁方解石具有较低的Sr/Ca和较高的 Mg/Ca[21],因此根据 Mg/Ca和Sr/Ca结果可以基本可以判定所测矿物类型。从表2和图3中可以看出,测试的碳酸盐矿物Mg/Ca比值最大为39.19 mmol/mol,而多数样品的Sr/Ca小于1 mmol/mol,因此推测测试的碳酸盐矿物多为方解石或者低镁方解石,仅有少数为文石矿物;而对于生物壳体,其Sr/Ca差别不大,集中分布在1 mmol/mol左右,但是根据Mg/Ca明显可以分为两类,基质生物壳体明显为高镁方解石,而包裹的生物壳体为低镁方解石或文石。
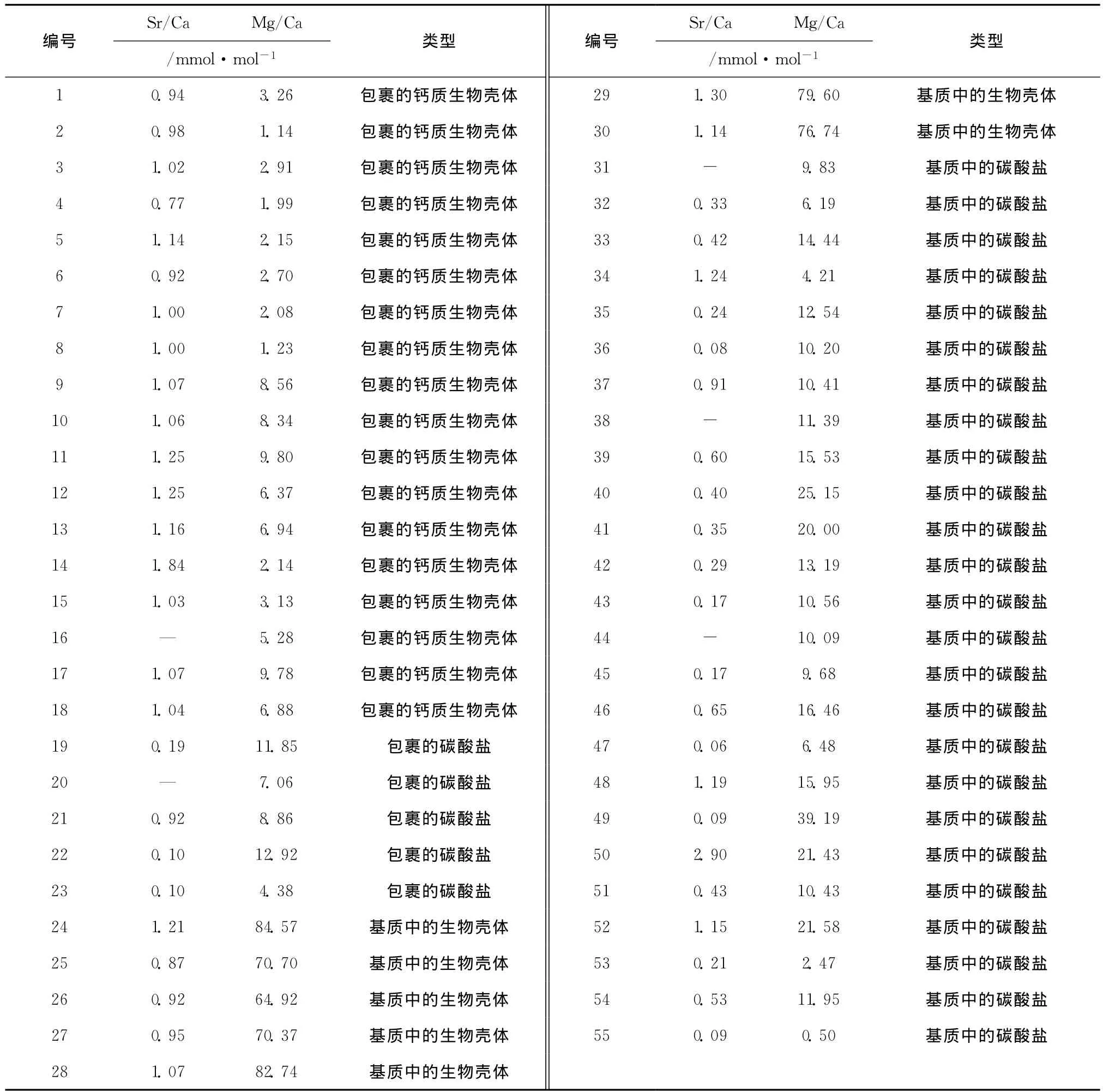
表2 自生碳酸盐矿物和钙质生物壳体的Sr/Ca和Mg/CaTab.2 Sr/Ca and Mg/Ca ratios of the authigenic carbonate minerals and the calcareous shells
5.2 生物壳体Mg/Ca比值的沉积环境指示
与甲烷渗漏有关的冷泉自生碳酸盐岩形成在海底和沉积物中[8],是环境信息变化的忠实记录者。甲烷渗漏会对沉积环境造成影响,钙质生物会对甲烷渗漏做出响应。南海北部海洋沉积物中的钙质生物壳体多种多样,有孔虫、翼足类和异足类、颗石类、介形虫和苔癣虫等均可能存在。而测试过程中所选的生物壳体均为随机过程,在薄片中明确钙质生物壳体的种属比较困难,但是根据形态可以肯定测试的钙质生物为有孔虫的有:包裹的钙质生物壳体中如图1c中测试点编号为1、2、3的生物壳体和图2b测试点编号 为14、15、16的生物壳体,基质的钙质生物壳体如图1d和图2h。而冷泉区域可以生长的钙质生物主要为有孔虫和双壳类,颗石(coccolith)也时有发现[7]。根据测试的生物壳体特征可以完全排除双壳类生物壳体的可能性。因此本文测试的生物壳体可能为不同种属的有孔虫和颗石。
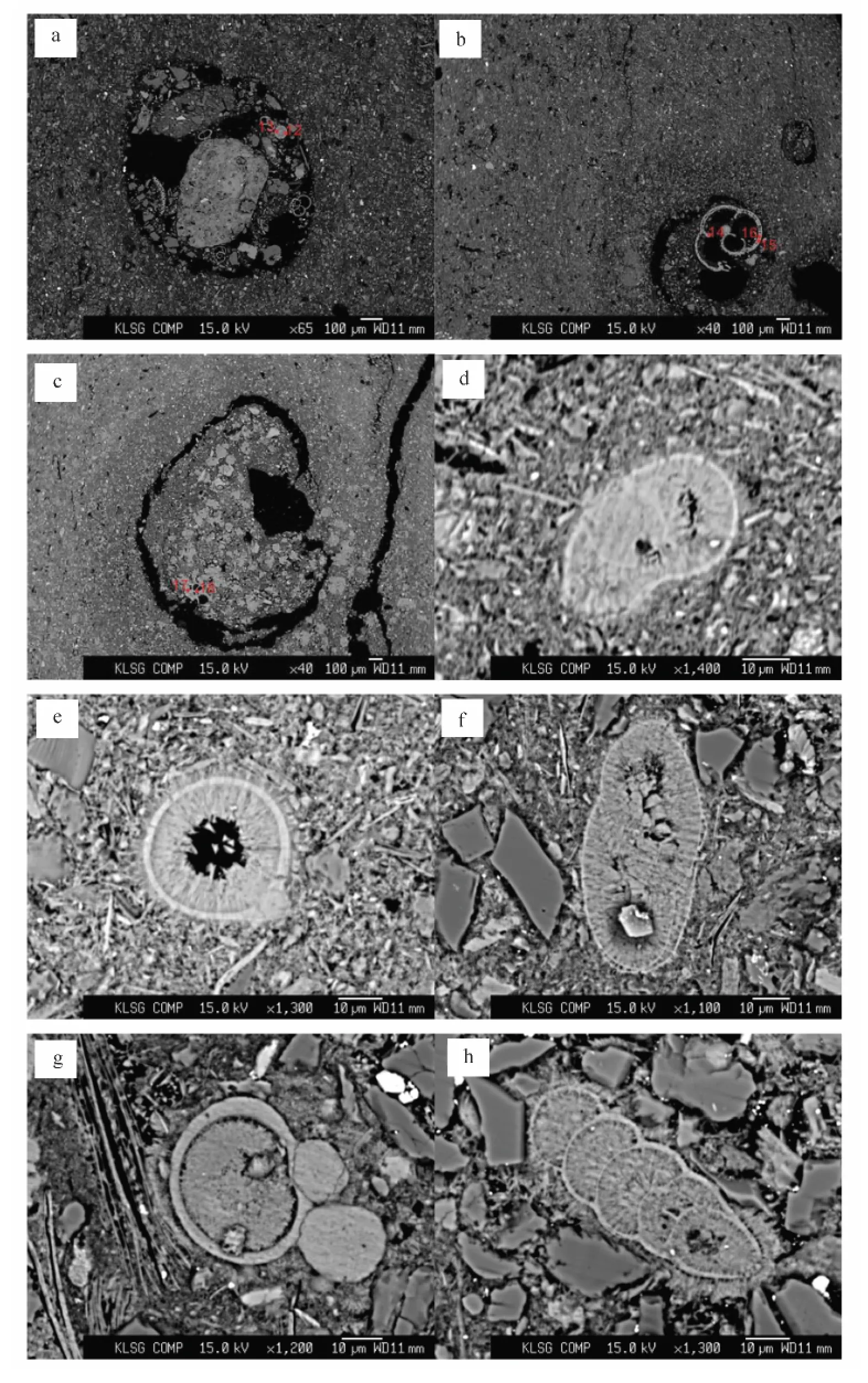
图2 测试的各种钙质生物壳体及位置(均为背散射图片)Fig.2 Studied Calcareous shells(all are the back scatter photographs)

图3 不同碳酸盐的Mg/Ca和Sr/Ca关系Fig.3 Relationships bet ween Mg/Ca and Sr/Ca ratios of the difference carbonates
有孔虫在生长过程中,从海水中吸收Ca、Mg等元素形成碳酸盐壳体,而现代海水中的Mg/Ca比值基本是一个常量(5.4 mol/mol)[22],因此有孔虫壳体Mg/Ca比值的变化是受到周围环境参数的影响而产生的。实验表明,有孔虫壳体Mg/Ca比值会随着海水温度升高而增高,这是因为Mg置换碳酸盐中的Ca是吸热过程,所以温度升高会导致壳体中Mg含量的增加[23]。有研究表明,有孔虫壳体的Mg/Ca受温度的变化范围在0.3~8 mmol/mol[24—26],但是无机碳酸盐的出现会影响生物壳体的Mg/Ca,其改变较温度效应至少大一个数量级[27]。因此,可以利用浮游和底栖种属生物的Mg/Ca来识别无机影响的印记[27]。文中包裹的钙质生物壳体Mg/Ca的变化范围1.14~9.78 mmol/mol,与温度控制的变化范围基本一致;而基质钙质生物壳体Mg/Ca变化范围为64.92~84.57 mmol/mol,远远超过了温度控制的变化,而符合无机碳酸盐的出现造成的Mg/Ca变化数量级。因此,基质钙质生物壳体Mg/Ca变化还受控于其他因素,比如无机碳酸盐的出现。而冷泉碳酸盐岩的形成过程正是无机碳酸盐的出现过程,在甲烷氧化古细菌和硫酸盐还原细菌共同作用下发生甲烷厌氧氧化导致环境碱度增加从而沉淀冷泉碳酸盐。而前人在研究热液和冷泉区双壳类时也指出,Sr和Mg是成岩作用过程中的敏感元素,它们的摩尔比值可能可以识别冷泉活动历史[28]。文中,包裹的钙质生物壳体和基质钙质生物壳体的Sr/Ca比较接近,差别较大的是Mg/Ca,因此推测它们的Mg/Ca差异源于受无机碳酸盐出现的影响与否,即基质钙质生物壳体是在甲烷渗漏环境下生长、钙化、死亡的,其较高的Mg/Ca受来自甲烷厌氧氧化作用沉淀的碳酸盐影响;而包裹的钙质生物壳体是在非甲烷渗漏环境下钙化的,由于生物扰动、灌洗等[29]作用以包裹团块的形式保存在碳酸盐岩中。
世界其他冷泉区一些生物壳体的Sr/Ca、Mg/Ca的测试结果如表3和图3所示,其中,B1-B4为沉积物中的生物碎屑;#7为双壳类壳体,是岩化的生物碎屑;G11-B和 G11-C均存在于碳酸盐岩中,G11-B的岩石学特征显示其由三期生长,主要碳酸盐矿物为黑灰色微晶文石,富含钙质微生物化石及硅质成分,测试的有孔虫为最早一期,根据文中分析富含甲烷流体沿通道(比如裂隙)上升导致碳酸盐矿物在碎屑周围沉淀,因此这些有孔虫死亡时尚未受甲烷渗漏影响;G11-C样品的形成与海底天然气水合物分解有关,气体主要沿前期存在的水合物层做水平扩散,碳酸盐岩形成时处在沉积物深部;758-1存在于碳酸盐岩的细粒基质。通过对钙质生物壳体赋存环境的分析可以看出,除了758-1沉积环境不明外,其他均为正常沉积环境下的产物,它们的Mg/Ca变化范围与文中包裹的钙质生物壳体一致,而与基质钙质生物壳体差别较大,从而也佐证了包裹的钙质生物壳体为正常沉积环境钙化死亡的,而基质中的钙质生物壳体是冷泉环境下钙化死亡的。
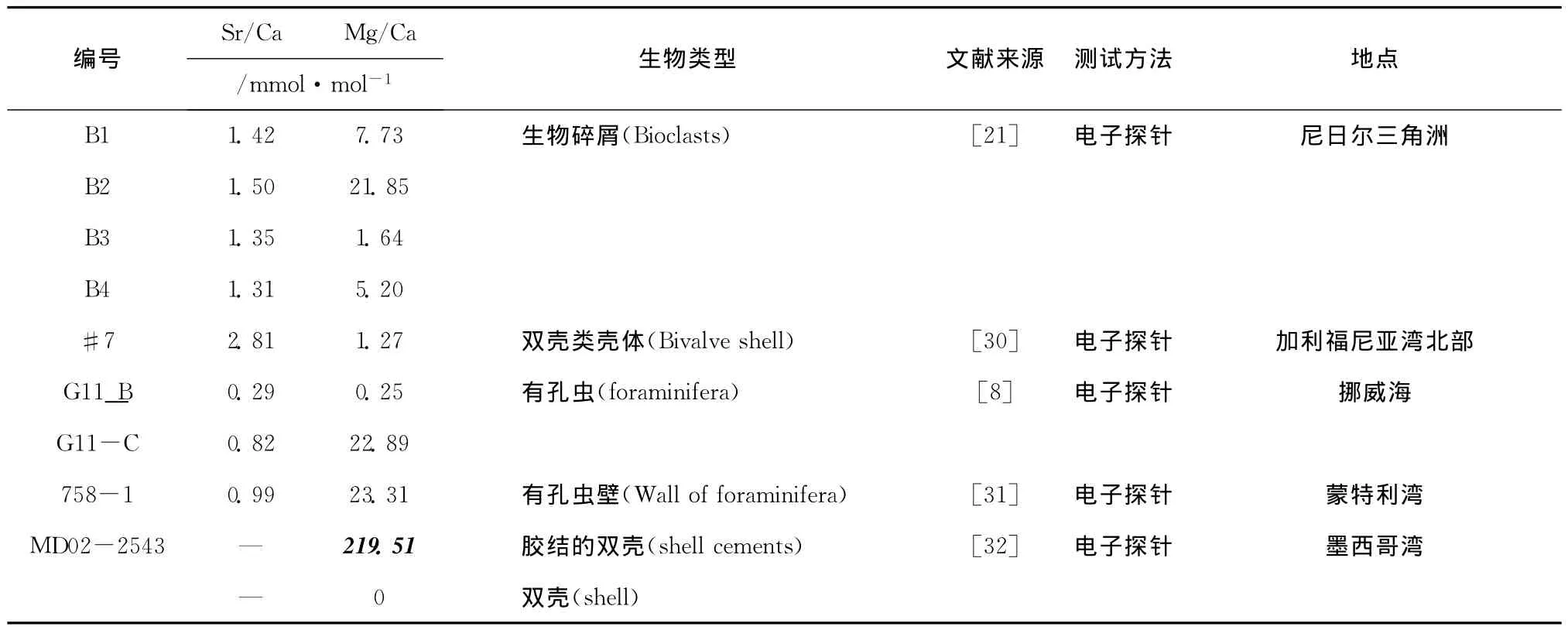
表3 其他冷泉区生物壳体的Sr/Ca和Mg/CaTab.3 Sr/Ca and Mg/Ca ratios of the calcareous shells in others cold seeps
南海各主要海区表层沉积物中G.sacculif er的Mg/Ca分析结果显示其变化范围为2.756~4.209 mmol/mol[33],与包裹的钙质生物壳体较为接近,而与基质中的钙质生物壳体差别较大,这也说明包裹的钙质生物壳体与基质钙质生物壳体沉积环境有别,包裹的生物壳体更接近正常沉积环境,而基质的生物壳体可能受甲烷渗漏影响。
有趣的是,墨西哥湾中 MD02-2543站位[32]顶部双壳类生物Mg/Ca也表现出了和本文类似的研究结果(表3),胶结的双壳类具有较高的 Mg/Ca(高达219.51 mmol/mol),其δ13C为-41.4‰[32],明显受甲烷渗漏影响;而另一双壳类生物(未胶结),基本不含Mg,其δ13C为-3.1‰[32],与正常海相沉积碳酸盐岩相差无几,这不仅证实了甲烷渗漏会对生物壳体中的Mg/Ca造成较大影响,也说明了本文研究的包裹的生物壳体更接近正常沉积环境,而基质的生物壳体可能受甲烷渗漏影响。
5.3 生物壳体Sr/Ca比值的成岩作用指示
Sr在研究成岩作用过程中非常有用,因为它的地球化学性质和Ca相似,取代文石或方解石中的Ca离子是其主要的沉淀方式,并且其在碳酸盐矿物中含量较高,而在其他非碳酸盐矿物中的含量可以忽略[9]。自生碳酸盐矿物的Sr/Ca变化范围为0.06~2.90 mmol/mol,平均值为0.53 mmol/mol;而钙质生物壳体的Sr/Ca变化范围为0.77~1.84 mmol/mol,平均值为1.08 mmol/mol(见表2)。因此自生碳酸盐矿物和生物壳体表现为明显不同的Sr/Ca(见图3),说明无机和有机作用过程对Sr元素的富集程度是不同的。
而不同期次的钙质生物壳体(见5.2)却表现为具有类似的Sr/Ca,包裹的钙质生物壳体的Sr/Ca变化范围为0.77~1.84 mmol/mol,平 均 值 为1.09 mmol/mol;基质钙质生物壳体的Sr/Ca变化范围为0.87~1.30 mmol/mol,平均值为1.07 mmol/mol。前已述及,基质钙质生物壳体生长在甲烷渗漏时期,而包裹的钙质生物壳体生长在非甲烷渗漏期。亦有研究表明,随着成岩作用的推进,Mg/Ca和Sr/Ca呈降低趋势[11]。因此推测最初形成的基质钙质生物壳体可能具有较现在高的Sr/Ca。
海洋有机成因碳酸盐中Sr含量除了受海水Sr含量影响外,还受控于多种因素,如,有孔虫和鳞鞭虫(Coccolithophores)中Sr含量主要根据种的不同变化,或者随着生长和钙化率而变化[34]。钙质生物壳体在海水中的生长时间相对于地质历史来说较短,可以认为海水Sr含量基本稳定,因此基质钙质生物壳体可能具有与包裹的钙质生物壳体不同的生长和钙化率,这种变化可能来自甲烷渗漏的影响。已有研究表明,甲烷渗漏会造成冷泉区底栖有孔虫在丰度分异度、优势种及13C等方面有别于非冷泉区底栖有孔虫的特征[35],但是甲烷渗漏对有孔虫钙化和生长率方面的影响还鲜有报道,值得进一步研究证实。
6 结论
通过对南海北部冷泉碳酸盐岩的镜下观察及对自生碳酸盐矿物及钙质生物壳体的Mg/Ca和Sr/Ca分析,得出以下几点认识和结论:
(1)南海北部冷泉碳酸盐岩具有包裹结构和孔洞结构,并且包裹结构的生物遗迹非常丰富;
(2)南海北部自生碳酸盐矿物及钙质生物壳体的Mg/Ca和Sr/Ca分析结果显示,自生碳酸盐矿物的Mg/Ca和Sr/Ca与所在位置关系不大,均表现为低到中等的Mg/Ca、低Sr/Ca的特征;而钙质生物壳体却根据其所在岩石中的不同结构位置呈现明显的分组特征,基质钙质生物壳体具有高Mg/Ca、中等Sr/Ca的特征,包裹的钙质生物壳体具有低Mg/Ca、中等Sr/Ca的特征;
(3)基质钙质生物壳体和包裹的钙质生物壳体具有截然不同的Mg/Ca,推测前者是在甲烷渗漏时期发生钙化的,而后者是在非甲烷渗漏期钙化的。
致谢:感谢2006年环境航次“大洋一号”全体人员为样品的获取所付出的艰辛劳动!
[1]Campbell K A.Hydrocarbon seep and hydrother mal vent paleoenviron ments and paleontology:Past develop ments and future research directions[J].Palaeogeography,Palaeocli matology,Palaeoecology,2006,232(2/4):362-407.
[2]Boetius A,Ravenschlag K,Schubert C J,et al.A marine microbial consortium apparently mediating anaerobic oxidation of methane[J].Nature,2000,407(6804):623-626.
[3]Teichert B M A,Eisenhauer A,Bohr mann G,et al.U/Th systematics and ages of aut higenic carbonates from Hydrate Ridge,Cascadia Margin:recorders of fluid flow variations[J].Geochimica et Cosmochi mica Acta,2003,67(20):3845-3857.
[4]陆红锋,刘坚,陈芳,等.南海台西南区碳酸盐岩矿物学和稳定同位素组成特征——天然气水合物存在的主要证据之一[J].地学前缘,2005,12(3):268-276.
[5]Feng D,Chen D F,Roberts H H.Petrographic and geochemical characterization of seep carbonate from Bush Hill(GC 185)gas vent and hydrate site of the Gulf of Mexico[J].Marine and Petroleum Geology,2009,26(7):1190-1198.
[6]Naehr T H,Birgel D,Bohr mann G,et al.Biogeochemical controls on authigenic carbonate formation at the Chapopote“asphalt volcano”,Bay of Campeche[J].Chemical Geology,2009,266(3/4):390-402.
[7]Bahr A,Pape T,Bohr mann G,et al.Authigenic carbonate precipitates from the NE Black Sea:a mineralogical,geochemical,and lipid biomar ker study[J].International Journal of Earth Sciences,2009,98(3):677-695.
[8]Mazzini A,Svensen H,Hovland M,et al.Comparison and i mplications from strikingly different authigenic carbonates in a Nyegga complex pockmar k,G11,Nor wegian Sea[J].Marine Geology,2006,231(1/4):89-102.
[9]Baker P A,Gieskes J M,Elderfield H.Diagenesis of carbonates in deep-sea sediments;evidence from Sr/Ca ratios and interstitial dissolved Sr2+data[J].Journal of Sedimentary Research,1982,52(1):71-82.
[10]Hampt G,Delaney M L.Influences on calcite Sr/Ca records from Ceara Rise and other regions:distinguishing ocean history and calcite recrystallization[M]//Shackleton N J,Curry W B,Richter C,et al.Proceedings of the Ocean Drilling Program,Scientific Result.1997:154,491-591.
[11]Joseph C,Campbell K A,Torres M E,et al.Methane-derived authigenic carbonates from modern and paleoseeps on the Cascadia margin:Mechanisms of formation and diagenetic signals[J].Palaeogeography,Palaeocli matology,Palaeoecology,2013,390:52-67.
[12]Han X Q,Suess E,Huang Y Y,et al.Jiulong methane reef:Microbial mediation of seep carbonates in the South China Sea[J].Marine Geology,2008,249(3/4):243-256.
[13]陈忠,颜文,陈木宏,等.南海北部大陆坡冷泉碳酸盐结核的发现:海底天然气渗漏活动的新证据[J].科学通报,2006,51(9):1065-1072.
[14]陆红锋,陈芳,刘坚,等.南海北部神狐海区的自生碳酸盐岩烟囱——海底富烃流体活动的记录[J].地质论评,2006,52(3):352-357.
[15]杨克红,初凤友,赵建如,等.南海北部烟囱状冷泉碳酸盐岩的沉积环境分析[J].海洋学报,2013,35(2):82-89.
[16]陈忠,杨华平,黄奇瑜,等.南海东沙西南海域冷泉碳酸盐岩特征及其意义[J].现代地质,2008,22(3):382-389.
[17]Mook W G.Environmental isotopes in the hydrological cycle:principles and applications[M]//UNESCO/IAEA.Paris,2000:91-123.
[18]Veizer J,Demovic R.Strontium as a tool in facies analysis[J].Journal of Sedimentary Research,1974,44(1):93-115.
[19]Veizer J,Demovic R.Environmental and climatic controlled fractionation of elements in the Mesozoic carbonate sequences of the western Carpathians[J].Journal of Sedimentary Research,1973,43(1):258-271.
[20]倪善芹,侯泉林,王安建,等.碳酸盐岩中锶元素地球化学特征及其指示意义——以北京下古生界碳酸盐岩为例[J].地质学报,2010,84(10):1510-1516.
[21]Bayon G,Pierre C,Etoubleau J,et al.Sr/Ca and Mg/Ca ratios in Niger Delta sediments:Implications for authigenic carbonate genesis in cold seep environments[J].Marine Geology,2007,241(1/4):93-109.
[22]Billups K,Schrag D P.Paleotemperatures and ice volu me of the past 27 Myr revisited with paired Mg/Ca and18O/16O measurements on benthic foraminifera[J].Paleoceanography,2002,17(1):1003,doi:10.1029/2000PA000567.
[23]李建如.有孔虫壳体的 Mg/Ca比值在古环境研究中的应用[J].地球科学进展,2005,20(8):815-822.
[24]Mashiotta T A,Lea D W,Spero H J.Glacial-interglacial changes in Subantarctic sea surface temperature andδ18O-water using foraminiferal Mg[J].Eart h and Planetary Science Letters,1999,170(4):417-432.
[25]Lear C H,Elderfield H,Wilson P A.Cenozoic deep-sea temperatures and global ice volu mes from Mg/Ca in benthic foraminiferal calcite[J].Science,2000,287(5451):269-272.
[26]Nürnberg D,Bij ma J,Hemleben C.Assessing the reliability of magnesiu m in foraminiferal calcite as a proxy for watermass temperatures[J].Geochi mica et Cosmochimica Acta,1996,60(5):803-814.
[27]Torres M E,Mix A C,Kinports K,et al.Is methane venting at the seafloor recorded byδ13C of benthic foraminifera shells?[J].Paleoceanography,2003,18(3):1062,doi:10.1029/2002PA000824.
[28]Rio M,Roux M,Renard M,et al.Chemical and isotopic features of present day bivalve shells from hydrother mal vents or cold seeps[J].PALAIOS,1992,7(4):351-360.
[29]Luff R,Wall mann K,Aloisi G.Nu merical modeling of carbonate crust formation at cold vent sites:significance for fluid and methane budgets and chemosynthetic biological communities[J].Earth and Planetary Science Letters,2004,221(1/4):337-353.
[30]Canet C,Anadón P,Alfonso P,et al.Gas-seep related carbonate and barite aut higenicmineralization in the northern gulf of California[J].Marine and Petroleu m Geology,2013,43:147-165.
[31]Stakes D S,Orange D,Paduan J B,et al.Cold-seeps and authigenic carbonate formation in Monterey Bay,California[J].Marine Geology,1999,159(1/4):93-109.
[32]Chen Y F,Matsu moto R,Paull C K,et al.Methane-derived aut higenic carbonates from the northern Gulf of Mexico--MD02 Cruise[J].Journal of Geochemical Exploration,2007,95(1/3):1-15.
[33]邓文峰,韦刚健,李献华.有孔虫的高精度 Mg/Ca比值的ICP-AES分析[J].海洋地质与第四纪地质,2005,25(2):147-151.
[34]Wien K,Kölling M,Schulz H D.Close correlation bet ween Sr/Ca ratios in bulk sediments from the southern Cape Basin and the SPECMAP record[J].Geo-Marine Letters,2005,25(4):265-271.
[35]向荣,刘芳,陈忠,等.冷泉区底栖有孔虫研究进展[J].地球科学进展,2010,25(2):193-202.
