滴水湖浮游动物群落结构及其与环境因子的关系
2014-09-19由文辉徐凤洁俞秋佳余华光
林 青,由文辉,2,*,徐凤洁,俞秋佳,余华光
(1. 华东师范大学资源与环境科学学院,上海 200062;2. 上海市城市化生态过程与生态恢复重点实验室,上海 200062)
滴水湖是目前国内最大的人工湖,面积5.56km2,平均水深3.7m,最深处6.2m。湖水通过大治河引自黄浦江,水源通过周边河网水系引水入湖,接纳地表径流并经过海闸排入东海,承担着防汛排涝、置换水体等重要功能。滴水湖水源本身富营养化程度较高,加上其背景底质底泥是河口海滩滩涂,底泥盐度也较一般内陆淡水湖底泥盐度高,蓄水初期水质不断演变,浮游生物群落结构也不够稳定,并随之发生相应演替,甚至发生过局部水华[1- 2]。
浮游动物在湖泊生态系统的要素循环中起着重要作用,是湖泊生态系统食物链及生物生产力的基本环节,浮游动物的存在不仅控制浮游植物的群落变化,而且一定程度上影响整个水华的过程,浮游动物在水生生态、水质监测和水污染治理方面的重要作用日益受到人们的广泛重视[3]。从滴水湖建成至今,已陆续有学者对滴水湖水质、浮游动物、浮游植物等方面进行相关研究。王延洋、周晓梅等[4- 5]对滴水湖及其引水河进行浮游动物群落结构的研究,从多方面角度对滴水湖进行水质生态学评价;而江敏[6]等从轮虫群落结构角度对滴水湖富营养状况进行研究;田华和刘水芹等对滴水湖进行了6a的水质监测,探讨了影响水体富营养化程度的主要指标[7]。作为一个每年通过引水河进行换水的新建人工湖泊,滴水湖生态系统相对脆弱,容易受到外界环境影响,随着近来滴水湖周边不断进行住宅、娱乐等设施的开发建设,滴水湖将面临更严峻的考验。因此对其浮游动物的群落演替值得长期跟踪研究和分析,以便及时采取生态修复措施,防止水华等现象再次发生。
本文调查研究了2012年滴水湖浮游动物的种类数、密度、生物量和物种多样性的动态变化,探究了滴水湖浮游动物群落结构特征与水质因子的关系,结合过去的研究分析了近年来滴水湖浮游动物群落结构的演替进展,并从水质理化指标、综合营养状态指数、浮游动物指标等多个角度评价了滴水湖水质的营养状况,旨在为滴水湖水质富营养化评价和可持续发展提供理论依据。
1 材料与方法
1.1 采样时间与样点设置
根据滴水湖的引河水系和湖区特点,于滴水湖布设8个采样点,包括湖心及各港口各一个(图1),其中C港、D港为引水河入口,A港为出水口,每月下旬采样1次。季节划分根据中国通常采用的“候温四季划分法”,上海12月至翌年2月为冬季,3—5月为春季,6—9月为夏季,10—11月为秋季。

图1 滴水湖采样点分布图
1.2 实验方法
1.2.1 浮游动物采集与处理
(1)定性标本
采用25号浮游生物网采集(网孔 0.064mm),于水面以下作“∞”状拖动浮游动物网数次,将浓缩于网头的水样收集于50ml的标本瓶,用4%甲醛溶液现场固定,以待镜检鉴定。种类鉴定参照《淡水微型生物图谱》、《淡水浮游生物研究方法》、《中国淡水轮虫志》、《中国动物志淡水枝角类》、《中国动物志淡水桡足类》[8- 12]。
(2)定量标本
用5L采水器于分别于水面以下0.5m处和1.5m处采集水样,混合均匀后置于1L塑料采样瓶中,加入鲁哥氏液现场固定,各样点均重复3次。实验室静置24h后,浓缩至200mL,再次静置24h,浓缩至50mL保存。定量计数前将沉淀样品充分摇匀,然后吸取1mL注入1mL计数框内,在10×10倍的光镜下全片计数。每个标本重复计数3—5次,取其平均值[4- 6,13- 14]。
1.2.2 理化因子测定
水体理化因子采用现场测定和实验室分析的方法。其中温度、pH、溶解氧等用WTW Multi 350i手提式多参数水质分析仪现场测定,透明度用塞氏盘测定,总氮、总磷、叶绿素a带回实验室参照《湖泊富营养化调查规范》进行测定分析。
1.3 数据处理方法
优势种是根据物种的出现频率及个体数量来确定, 用优势度来表示。优势度计算公式[15]:
Y=fiPi
式中,Y是优势度,fi是第i物种的出现频率,Pi是第i物种个体数量占总个体数量的比例, 当Y大于0.02时, 确定为优势种。
根据中国环境监测总站制定的《湖泊(水库)富营养化评价方法及分级技术规定》,运用综合营养状态指数法对滴水湖生态系统健康状况进行评价。综合营养状态指数计算公式:
TLI(∑)=∑WjTLI(j)
式中,Wj为第j种参数的营养状态指数的相关权重;TLI(j)代表第j种参数的营养状态指数。
同时采用Shannon-Wiener多样性指数H′和Margalef丰富度指数D[16- 17]评价浮游动物群落多样性并对水质进行评价:
H′=-∑PilnPi,D=(S-1)/lnN
式中,Pi=Ni/N,Ni表示样品中第i种物种的个体数,N表示样品中所有种类的总个体数,S表示样品中所有物种的种类数。两种指数的评价标准均分为5个等级:0—1为多污型,1—2为α-中污型,2—3为β-中污型,3—4为寡污型,>4为清洁水体。
相关性分析:选取浮游动物种类数(ZS)、密度(ZD)、生物量(ZB)、Shannon-Wiener多样性指数(H′)、Margalef丰富度指数(D)5个生物指标及水温(Wt)、溶解氧(DO)、pH、透明度(SD)、总氮(TN)、总磷(TP)、叶绿素a(Chla)和综合营养状态指数(TLI)8个理化指标,取自然对数后采用SPSS18.0软件对滴水湖浮游动物与环境因子做Pearson相关性分析。
多元逐步回归分析:以浮游动物种类数(ZS)、密度(ZD)及生物量(ZB)作为因变量,以水质理化因子为自变量,采用SPSS18.0软件进行多元逐步回归分析,分别建立最优回归方程,筛选出对浮游动物种类数、密度和生物量变化影响显著的水质因子。
冗余分析(RDA):运用Canoco4.5软件对浮游动物与环境因子进行RDA分析。在进行RDA分析之前,先进行降趋对应分析(DCA),当DCA分析结果中排序轴梯度最大值小于3时选择RDA分析对浮游动物与环境因子间的关系进行进一步分析。
2 结果与分析
2.1 环境因子分析
各项理化指标结果如下:水温6.2—30.2℃,年均17.6℃,夏季最高,冬季最低;pH 7.91—10.30,年均8.67,显示滴水湖水体呈弱碱性;溶解氧6.08—11.61 mg/L,年均7.63 mg/L,冬季最高,秋季最低;总氮TN、总磷TP和叶绿素a浓度全年波动较大,TN 0.20—9.34 mg/L,年均3.09 mg/L; TP 0.03—2.04 mg/L,年均0.24 mg/L;叶绿素a 6.11—54.84 mg/m3,年均21.67 mg/m3;TLI(∑) 46.12—76.73,年均60.17,四者均为秋季最高;全湖平均透明度为58.2cm,各季节变化不大。经单因素方差分析,各理化指标样点间差异不大,但不同季节差别显著(P<0.01)。水质理化指标及综合营养状态指数各月平均值及年均值见表1。通过对滴水湖近几年研究[2,4- 6,13- 14,18- 19]的水质情况对比发现,从2006年至2012年,水温、pH、透明度平均值变化不大,溶解氧各年间变化有所起伏,但都维持在6mg/L以上,TN、TP总体有逐年升高的趋势。

表1 滴水湖水质理化指标月均值
根据国外湖泊富营养化分级标准的范围、中国湖泊富营养化评分和分级标准[20]对滴水湖营养状态进行评价:通过TN、TP和TLI(∑)年均指标均可得出滴水湖处于中富营养-富营养化水平,通过溶解氧指标可得出滴水湖处于富营养化水平,通过叶绿素a和透明度年均指标可得出滴水湖处于中营养-中富营养化状态。根据湖泊(水库)富营养化评价方法及分级技术规定所示的水质类别与评分值对应表,以及各季节TLI(∑)值,2012年滴水湖各个季节营养化程度不同,富营养化程度高低为秋季(70.65)>春季(65.08)>夏季(62.75)>冬季(59.17),春、夏、秋季营养化程度较冬季高,属于中富营养,冬季则属于轻富营养。综合而言,可以认为滴水湖处于中富营养化状态。
2.2 浮游动物种类组成
2.2.1 种类数、密度及生物量季节变化
在对滴水湖的调查中,共鉴定浮游动物61种,其中轮虫种类数最多,为45种,占74%,枝角类种类数最少,为7种,占11%,桡足类9种,占15%。浮游动物种类数出现最多的在夏季,共检出40种,其次为春季37种,秋、冬季较少,分别为25和22种。四个季节均为轮虫占优势,其中春季轮虫种类数比例最高,占86%,枝角类和桡足类种类数比例分别是夏季和秋季最高,为15%和32%,枝角类仅在春季和夏季有个别发现,秋冬两季未有发现。
各类群浮游动物的密度和生物量水平分布及月变化见图2、图3。浮游动物密度作为表征湖泊营养程度的生物指标之一,<1000个/L为贫营养,1000—3000个/L为中营养,>3000个/L为富营养[21],滴水湖浮游动物月平均密度变化范围为212—2968个/L,年均1200个/L,因此滴水湖2012年处于中营养状态。浮游动物生物量月变化范围为0.11—5.69mg/L,年均1.67mg/L,二者整年的变化趋势(见图2)基本一致,最高值和最低值都分别出现在7月和1月。各样点间浮游动物年均密度波动范围在1055—1476个/L,最高点出现在样点1,其次样点2为1384个/L, 最低点出现在样点4,为1055个/L。年均生物量波动范围在1.39—2.40mg/L,最高点同样出现在样点1,最低点出现在样点3。浮游动物密度的季节变化为夏季(1940个/L)>春季(1018个/L)>秋季(1010个/L)>冬季(522个/L),存在显著的季节性变化(ANOVA,F=70.358,P=0.000),各样点间差异不显著。生物量的季节变化为夏季(2.89mg/L)>秋季(1.64 mg/L)>春季(1.33 mg/L)>冬季(0.38 mg/L),也存在显著的季节性变化(ANOVA,F=20.067,P=0.000),各样点间差异不显著。

图2 各类群浮游动物月平均密度及月平均生物量变化

图3 各类群浮游动物年均密度及年均生物量水平分布
2.2.2 优势种
滴水湖浮游动物全年平均优势度>0.02的共有12种,包括10种轮虫和2种桡足类,枝角类不形成优势种。多数优势种属于中污型指示种,其中属于α-中污型的萼花臂尾轮虫(Brachionuscalyciflorus)全年都有出现,而出现优势种最多的月份为7月和8月,表明夏季滴水湖浮游动物群落结构相对较为复杂。各优势种出现月份及指示的污染类型见表2。
2.3 浮游动物多样性指数及水质评价

图4 滴水湖浮游动物多样性指数季节变化
滴水湖浮游动物Shannon-Wiener多样性指数H′和Margalef丰富度指数D月变化和水平变化见图4、图5。Shannon-Wiener多样性指数H′八个样点全年变化范围为0.57—2.34,年均1.69,最高和最低值分别出现在5月的样点1和12月的样点6。丰富度指数D变化范围为0.00—1.69,年均1.03,最高和最低值分别出现在5月的样点6和10月的样点8。Shannon-Wiener多样性指数H′的季节变化均为春季最高,冬季最低,Margalef丰富度指数D则是春季最高,秋季最低,二者季节差异均非常显著(ANOVA,F=25.872,P=0.000;F=20.953,P=0.000),而各样点间的差异不大。根据多样性指数评价法,滴水湖水质春、夏两季属于α-中污型,秋、冬季则属于α-中污型-多污型,总体而言属于α-中污型。
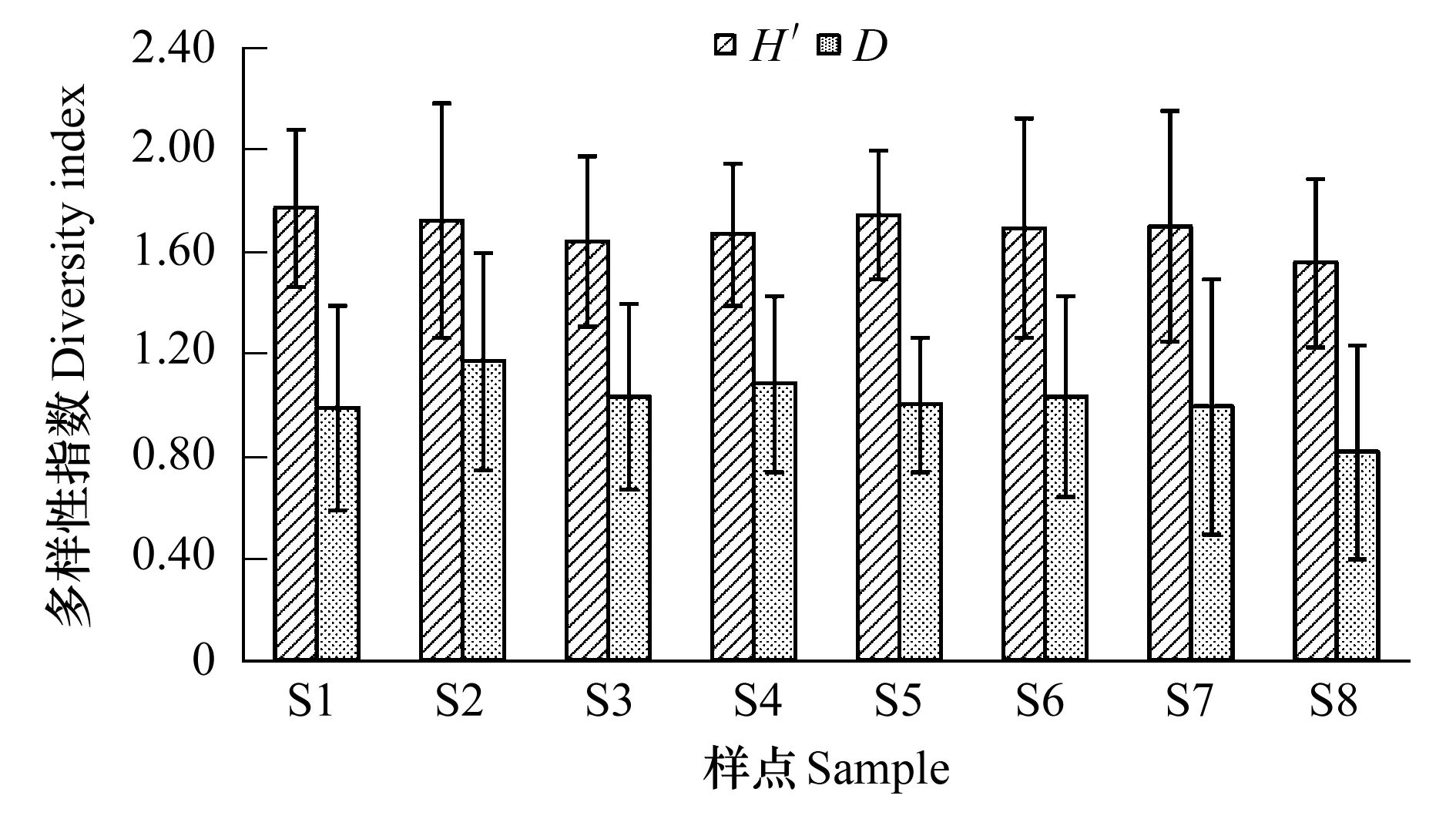
图5 滴水湖各样点浮游动物多样性年均值
2.4 浮游动物与环境因子的关系
2.4.1 相关性分析
对滴水湖各样点主要理化指标与浮游动物种类数、密度和生物量取自然对数后进行SPSS相关性分析(表3),表中数据为相关系数,ZS表示浮游动物种类数,ZD表示浮游动物密度,ZB表示浮游动物生物量。从表中可见,浮游动物种类数与水温、透明度、叶绿素a呈显著正相关,而与溶解氧、pH和TP极显著负相关;浮游动物密度与水温、透明度、TN、叶绿素a及TLI(∑)呈显著正相关,而与溶解氧、pH呈显著负相关;浮游动物生物量与水温、TN、叶绿素a、TLI(∑)呈显著正相关。表明在一定范围内,随着水温和作为表征浮游植物现存量的叶绿素a以及水体营养程度的提高,浮游动物会因为食物来源的增加而会有所增加,但水体碱性的提高和过高的营养盐可能会导致敏感种数量下降。

表2 滴水湖浮游动物全年优势种及指示污染类型

表3 浮游动物与环境因子相关性分析
2.4.2 多元回归分析
通过多元逐步回归分析筛选出对滴水湖浮游动物种类数、密度和生物量变化相对重要的水质因子,建立最优多元线性回归方程,并进行显著性检验。由表4可知,滴水湖浮游动物种类数的回归方程有水温、溶解氧、TP和pH 4个因子入选;浮游动物密度的回归方程有水温、TN、叶绿素a和透明度4个因子入选;生物量的回归方程有水温和TN 两个因子入选。其中水温在3个回归方程中都入选,表明水温是对滴水湖浮游动物群落结构变化起决定性作用的水质因子。TN也在浮游动物密度和生物量两个回归方程中入选,表明作为浮游动物重要食物来源的营养盐也是影响浮游动物群落结构变化的重要因素。经F检验,方差分析F值的显著水平P=0.000,均远小于0.05,表明回归极显著。

表4 浮游动物与环境因子的多元逐步回归方程
2.4.3 浮游动物与环境因子的RDA分析
运用Canoco4.5软件,根据浮游动物密度至少在一个样点中的百分比≥1%并且在8个样点中出现的频度≥30%,选取34种浮游动物进行DCA分析,浮游动物代码见表4。结果显示最大的梯度长度为0.515,小于3,因此选择对物种数据与环境因子数据作RDA分析,即选取8个样点的7个水质理化因子及34种浮游动物生物量数据,研究浮游动物与环境因子间的关系。结果显示,前两个轴的特征值分别为0.555和0.220,共解释了浮游动物群落变异程度的77.5%,物种和环境因子相关系数都达到1,表明排序能够很好地反映浮游动物与水质因子间的关系。物种的两个排序轴之间相关系数和环境因子的两个排序轴之间的相关系数均分别为0,表明排序结果是可靠的。

表5 RDA分析浮游动物代码

图6 浮游动物与环境因子的RDA分析排序图
RDA分析排序图(图6)显示,排序轴1与TN的正相关性最大,相关系数为0.5847,其次是TLI和水温,相关系数分别为0.4219、0.3166,pH则与排序轴1的负相关性最大(-0.4806);排序轴2与TP、TLI指数正相关性最大(0.8726,0.6434),而与透明度呈负相关(-0.2720)。曲腿龟甲轮虫和褶皱臂尾轮虫与叶绿素a的正相关性最大,而盘状鞍甲轮虫、前节晶囊轮虫、中华窄腹剑水蚤和桡足类无节幼体与之呈负相关性,可能难以承受浮游植物的大量繁殖;小巨头轮虫与TLI正相关关系明显,表明其是喜富营养化条件下生长的种,反之暗小异尾轮虫则最偏向于营养程度较低的水体;角突臂尾轮虫和小巨头轮虫受水温影响最大,与之呈现密切的正相关关系;萼花臂尾轮虫和矩形龟甲轮虫喜好在氮营养程度高的水体中生存,与TN密切相关,而缘板龟甲轮虫和壶状臂尾轮虫则与TP的正相关性最大。因此TN、TP、叶绿素a和pH也是影响滴水湖浮游动物群落的重要因素。
DCA分析排序图能较好地解释物种与环境因子的关系,同时反映出不同物种在生态习性上的异同。如曲腿龟甲轮虫和褶皱臂尾轮虫、前节晶囊轮虫和桡足类无节幼体、壶状臂尾轮虫和长圆疣毛轮虫在排序图中的位置比较接近,表明它们在生态适应性上有较为相似;而同一科属的浮游动物对环境的要求也可能存在一定差异,如同为臂尾轮属的方形臂尾轮虫和裂足臂尾轮虫在排序图中位置较远, 表现出对环境资源的不同依赖。
3 讨论
3.1 滴水湖浮游动物群落特征及优势种演变
滴水湖浮游动物种类组成中,轮虫占绝对优势,桡足类次之,而枝角类仅在春夏个别月份有出现。浮游动物的种类数、密度及生物量受到轮虫的影响最大,总体呈现出春夏季较秋冬季高的现象,这与过去的研究[4- 6,13- 14,18]一致。滴水湖作为人工湖泊,每年会在春节前后进行不定期换水,引水河靠近生活区,水体环境复杂,营养程度高,且相对于水生植物极度缺乏的湖区水体而言,引水河不同河段生长有不同的高等水生植物。更高的有机质含量可以为轮虫提供更多食物来源,而水生植物也能为轮虫创造相对适宜的多样性生境。滴水湖春夏两季轮虫的种类数相对较高,多样性也是春季最高,而且作为引水河入口的样点1和样点2浮游动物密度、生物量也最高,除了水质因子的影响和自身的季节演替外,与春季换水也有一定关系。不同类型的湖泊,浮游动物出现的高峰期不同,一般趋势是有水草湖泊的浮游动物密度高峰出现于春季和冬季,无水草的湖泊多出现于夏季[5]。本研究中,滴水湖作为水生植物缺失的湖泊,浮游动物密度和生物量高峰均出现于夏季,且浮游动物中占绝对优势的轮虫世代交替周期较短,对环境敏感的种类生存力弱,春、秋两季滴水湖水体相对较高的营养水平及可能出现的藻类季节性爆发也会影响轮虫数量的发展。
滴水湖不同季节出现的优势种有所差异,所指示的污染等级也不同,多数种类被认为是喜富营养化条件下生长的种类。除了优势种之外,滴水湖也出现了多种富营养化水体的指示种,包括螺形龟甲轮虫、前节晶囊轮虫、剪形巨头轮虫、卵形鞍甲轮虫、长刺异尾轮虫(Trichocercalongiseta)、蚤状溞(Daphniapulex)、短尾秀体溞(Diaphanosomarachyurum)、椎尾水轮虫等[4- 6,22- 24]。参考近年来对滴水湖浮游动物的研究[4- 6,25],从2006年至今,浮游动物种类数呈不断上升的趋势,特别是轮虫种类数明显增加,总密度总体呈现出逐年下降的趋势,轮虫密度在2008—2009年略有上升,但之后便大幅下降,枝角类密度略微有所上升,但变化甚微,桡足类密度各年间有所波动。(2010年的研究中湖区采样点较少,可能使偶见种的出现概率降低,因而种类数和密度均要小于其他几次研究。)优势种与过去的研究相比,主要的变化是轮虫优势种的增加,尤其是耐污的臂尾轮属有所增加,表明滴水湖水体富营养化程度有升高的趋势。螺形龟甲轮虫和汤匙华哲水蚤是首次成为滴水湖优势种,其他种则均曾在过去对滴水湖浮游动物的研究中成为过优势种,但这两个种曾经是地理位置邻近的淀山湖、千岛湖的浮游动物优势种[23- 24]。螺形龟甲轮虫是广布性种类,具有较强的生态适应性和耐污能力,汤匙华哲水蚤也是广泛生活于我国亚热带和温带湖泊、河流的淡水种类[10,12]。滴水湖浮游动物优势种和长江中下游地区一些自然湖泊的常见优势种类似,表明其水域生态系统开始逐步向自然湖泊生态系统方向发展。
3.2 浮游动物与环境因子的关系
大量研究表明,水温是影响浮游动物生长发育、群落组成和数量变化等最重要的环境因子之一,也是影响浮游动物分布的重要因素[26- 29]。田湾核电站海域水温与浮游动物密度及种类数均有显著相关性[30];淀山湖浮游动物种类数与水温呈显著正相关[31]。本研究中,滴水湖浮游动物的种类数、密度和生物量从春季到夏季均呈现出逐步上升的趋势,在夏季达到高峰,而后随着水温的下降而逐步减少,相关性分析和回归分析也显示浮游动物种类数、密度和生物量与水温均呈极显著正相关,相关系数和回归系数在理化因子中都是最高。RDA分析也显示多种优势种与水温呈正相关关系。
浮游植物是浮游动物的重要食物来源之一,水体中浮游植物数量的高低与轮虫的生长繁殖密切相关。广东省城市湖泊轮虫密度与叶绿素a呈极显著正相关[32],宁夏鸣翠湖水体叶绿素a也与轮虫的密度和生物量有显著的相关关系和回归关系,且对轮虫的密度和生物量具有较强的直接影响作用[33]。滴水湖浮游动物密度也与生物量也与叶绿素a呈极显著正相关关系和回归关系。N、P等营养元素能调节浮游植物的生长,在一定浓度范围内对浮游植物的生长有促进作用,从而对浮游动物的数量增长产生间接影响,但过高的营养盐也会导致藻类爆发,蓝藻、绿藻作为食物而言对浮游动物不利,除了藻毒会抑制浮游动物的摄食之外,鱼类因藻类不适口,也会改食浮游动物[34],从而影响浮游动物数量的变化和导致敏感种消失。有研究表明,在广范围的温带-热带湖泊,随着营养上升,浮游动物密度也上升[35],小型种类有时也会上升[36]。本研究中,浮游动物种类数、密度和生物量均与综合营养状态指数TLI呈显著正相关;RDA分析也显示,优势种中超过一半的种与水温、TN、TP、叶绿素a及TLI指数呈正相关,而与透明度呈负相关,此外, TN和TP分别入选了浮游动物密度和生物量的逐步回归方程,并产生正向作用,但TP与种类数呈显著负相关关系和回归关系,同时显示出TP对浮游动物种类的抑制作用和对生物量增长的间接影响。
水体的pH值对浮游动物也具有重要影响,不同种类的轮虫,其适宜生长的pH不同,许多枝角类也对较高及较低的pH值非常敏感[37]。在东湖,由于pH值的上升,导致一些酸水性轮虫种类几近消失[38]。本研究中,浮游动物密度和生物量也随着pH的升高而相应地减少,RDA分析也表明第一排序轴与pH的负相关性最大。
除了水温、叶绿素a、pH、营养盐等会对滴水湖浮游动物群落产生重要影响外,鱼类、水生植物、盐度等也是不可忽略的环境因素。2004年底滴水湖初次引水,2005—2006年,滴水湖水质基本处于Ⅳ类,为轻富营养化程度,但明显已经向中富营养化方向发展[2],由于其水源大治河水质在Ⅴ类甚至劣Ⅴ类,给滴水湖水体造成一定的营养负荷,加上底泥盐度较一般淡水湖高,使滴水湖建成初期营养物质过剩,而湖区水生生物链缺乏,小型浮游动物轮虫占优势,生态系统脆弱,浮游动物群落的变化趋势与引水河主要入水口的C港和D港相似[4,13], 2006—2007年一度产生蓝藻爆发及局部水华现象[1,7];2008—2009年的研究[5]显示,滴水湖与外围水体相比,浮游动物种类较贫乏,群落结构较为均一,而外围水体则明显种类更丰富,密度和生物量均较湖区要高,群落结构差异显著;其后,滴水湖管理公司加强了水质监测管理,多所高校也在引水河道内进行了生态浮床、滨岸带植物种植、沉水植物移植、大型溞投放等生态修复工程项目[39, 40],希望改善滴水湖引水河水质,在一定程度上阻截入湖污染源,湖区内也开始投放鲢、鳙等滤食性鱼类,加上湖水自净作用,水质曾有所好转[41],随着湖水盐度的逐年降低和化学需氧量COD的增加,浮游动物及优势种和多样性也在增加,优势种不断变化,推进了滴水湖浮游动物的演替进程[18]。但滴水湖仍然非常缺乏高等水生植物为浮游动物提供繁殖和栖息的场所,不合理地放养鱼类反而会浮游动物的群落变化产生负面影响,因鲢、鳙等大多优先捕食大型甲壳浮游动物如枝角类等,间接控制了小型浮游动物的竞争者和捕食者,加剧了浮游动物群落的小型化,降低了群落的多样性,这与滴水湖浮游动物群落中枝角类贫乏的现状不无关系。近年来的研究均显示滴水湖多处于中富营养化程度,还可能与周边临港新城的住宅、商业、娱乐设施建设等施工污染、海事大学等学校生活污染、工业污染等外源污染增加有关[42- 43]。因此,在滴水湖富营养化防治过程中,应将严格控制引水河及周边污染源与水生植被的种植及鱼类的合理投放结合起来。
4 结论
本研究着重分析了滴水湖浮游动物群落的季节动态及其与环境因子的关系。滴水湖浮游动物以小型浮游动物轮虫占绝对优势,浮游动物群落结构在各样点间差异不大,但季节差异显著。由于受到外围水体引水河生境影响,轮虫种类数和密度在春夏季明显高于秋冬季,优势种中轮虫种类也比过去有所增加。对浮游动物与环境因子的相关性分析和回归分析显示,水温是影响滴水湖浮游动物群落结构变化的决定性因子,同时,叶绿素a、TN和TP也是影响群落结构的重要因素。近年来随着滴水湖盐度的降低和营养盐的增加,浮游动物淡水种类逐渐增加,优势种和多样性也不断变化。综合理化指标、浮游动物多样性指数、综合营养状态指数及浮游动物优势种指示种等多种方法,得出2012年滴水湖水质属于α-中污型,营养程度为中富营养水平。总的来说,利用理化指标和利用浮游动物指标对滴水湖水质评价得到的结果基本一致。
:
[1] Liu X Q, Tian H, Liang G K. Large artificial lake ecological environment evolution analysis and countermeasures investigation. Shanghai Water, 2009, 25(3): 25- 29, 33- 33.
[2] Wang H Y, Zhou M J. Lingang new city---eutrophication status evaluation and control measures of dishui lake. Shanghai Water, 2006, 22(4): 24- 26, 28- 28.
[3] Sun G, Lang Y, Fang Y. Characteristics of zooplankton community in the aquatic ecosystem of nanhu lake of Changchun. Journal of Jilin University, 2006, 44(4): 663- 667.
[4] Wang Y Y. Community Structure of Zooplankton and Ecological assessment of water quality in Dishui Lake[D]. Shanghai: Shanghai Normal University, 2008.
[5] Zhou X M. Comparative Study on Community Structures of Zooplankton in Dishui Lake and its surrounding waters[D]. Shanghai: Shanghai Normal University, 2010.
[6] Jiang M, Ren Z A, Xing B, Lu L, Hu W T, Chen L J, Li J Y. Community structure of rotifer and water quality assessment in dishui lake river system of Shanghai. Science & Technology Review, 2012, 29(22): 31- 37.
[7] Tian H, Liu S Q, Fang W, Bian J Y, Li J Y. Evolution tendency of water quality and eutrophication analysis of an artificial Lake— dishui lake. Journal of Hydroecology, 2012, 32(6): 26- 31.
[8] Zhou F X, Chen J H. Freshwater Micro-organisms Map (Second Edition). Beijing: Chemical Industry Press, 2010: 303- 303.
[9] Zhang Z S, Huang X F. Freshwater Plankton Research Methods. Beijing: Science Press, 1991: 232- 232, 256- 258.
[10] Wang J J. Chinese freshwater rotifer Blog. Beijing: Science Press, 1961: 20- 21.
[11] Jiang X Z, Du N S. Fauna Sinica (Crustacea Freshwater Cladocera). Beijing: Science Press, 1979: 43- 74.
[12] Shen J R. Fauna Sinica (Crustacea Freshwater Copepoda). Beijing: Science Press, 1979: 27- 27, 31- 36.
[13] Wang Y Y, Li X B, Wu B, Wu Q, Xu X L, Wang Q X. Notes on zooplankton in dishui lake. Journal of Shanghai Normal University, 2008, 37(2): 167- 172.
[14] He W, Xue J Z, Wu H X. Community structure and spatial-temporal pattern of planktonic rotifers of lake dishui in the reclaimed beach land,Shanghai. Journal of Lake Sciences, 2011, 23(2):257- 263.
[15] Mchaughton S J. Relationships among functional properties of Californian grassland. Nature, 1967, 216(5111): 168- 168.
[16] Shannon C E, Weaver W. The Mathematical Theory of Communication. Urbana: University of Illinois Press, 1949.
[17] Margalef R. Information theory in ecology. General Systems Yearbook, 1958, 3: 36- 71.
[18] Wang Z F, Zhou X M, Wang Y Y, Luo Y T, Wang Q X. Journal of Shanghai Normal University. 2011, 40(3): 295- 300.
[19] Liu Z Y, Xu J P. Eutrophication assessment of dishui lake from 2006 to 2010. Environmental Science and Management, 2012, 37(4): 169- 172.
[20] Jin X C, Tu Q Y. Lake Eutrophication Survey Specification. (Second Edition). Beijing: China Environmental Press, 1990.
[21] Zhang J M, He Z H. Inland water fisheries Natural Resources Survey Manual. Beijing: China Agriculture Press, 1991.
[22] Wang F J, Hu Z Q, Tang J, Liu L S, Zhao H Q. Evaluation of water quality and the type of nourishment in the eastern zone of lake chaohu by means of zoopIankton. Ecologic Science, 2006, 25(6): 551- 552.
[23] Feng D X, Liu Y, Yu N, Chen L, Li E C, Chen L Q. Study on meta-zooplankton community structure of dianshan lake. Journal of East China Normal University, 2011, (6): 126- 128.
[24] Shi W. An Analysis of the Spatial and Temporal Variation of Zooplankton Community Structure and the Correlation with Influencing Factors in Qiandao Lake[D]. Shanghai: Shanghai Ocean University, 2010.
[25] Ren Z A. Research on the Chief Zooplankton Community in Dishui Lake and Its River Course[D]. Shanghai: Shanghai Ocean University, 2012: 12- 13.
[26] Dumont H J. Biogeography of rotifers. Hydrobiologia, 1983, 104(1): 19- 30.
[27] Dussart B H, Fernando C H, Matsumura-Tundisi T, Shiel R J. A review of systematics, distribution and ecology of tropical freshwater zooplankton. Hydrobiologia, 1984, 113(1): 77- 91.
[28] Sommer U. Plankton Ecology, Succession in Plankton Communities. Berlin: Springer-Verlag, 1989.
[29] Chen G R, Zhong P, Zhang X F, Xie Y F, Li C H. Zooplankton and its relationship with water quality in Huizhou West Lake. Journal of Lake Sciences, 2008, 20(3): 351- 356.
[30] Wu J X, Yan B L, Feng Z H, Li Y, Xu J T, Li S H, Shen X. Zooplankton ecology near the tianwan nuclear power station. Acta Ecologica Sinica, 2011, 31(22): 6902- 6911.
[31] Liu Y, Studies on Community Structure of Zooplankton in Dianshan Lake[D]. Shanghai: East Normal University, 2010.
[32] Wang Q, Liu L J, Yang Y F. Response of rotifer community characteristics to environmental factors in a reservoir, Southern China. Acta Ecologica Sinica, 2010, 30(13): 3385- 3395.
[33] Qiu X C, Zhao H X, Sun X X. Rotifer community structure and its relationships with environmental factors in mingcui lake. Journal of Ningxia University Natural Science Edition, 2012, 33(1): 62- 68.
[34] Nilssen J P. Tropical lake-functional ecology and future development: the need for a process-orientated approach. Hydrobiologia, 1984, 113(1): 231- 242.
[35] Patalas K. Crustacean Plankton and the eutrophication of the St. Lawrence Great Lakes. Journal of the Fisheries Research Board of Canada, 1972, 29(10): 1451- 1462.
[36] Harper D. Eutrophication of Freshwaters: Principles, Problems and Restoration. London: Chapman and Hall, 1992: 120- 127.
[37] Huang X F, Hu C Y, Wu D T. Rotifers in donghu lake. Acta Hydrobiologica Sinica, 1985, 9(2): 129- 143.
[38] Liu J K. Ecological Research in Donghu Lake. Beijing: Science Press.1990.
[39] Huo Y Z, He W H, Luo K, Wang Y Y, Zhang Y J, Tian Q T, He P M. Bioremediation efficiency of applying daphniamagna and submerged plants a case study in dishui lake of Shanghai China. Chinese Journal of Applied Ecology, 2010, 21(2): 495- 499.
[40] Tong Y, Xu C Y, Hu X Q, Zhang W Y, You W H. Research of the ecological restoration results of the combined ecological project in yinshui river, dripping lake in spring. Journal of Shanghai Ocean University, 2011, 20(6): 930- 937.
[41] Zhou X L. Evaluation of water quality for dishuihu lake with the dethod of comprehensive water quality index. Water Purification Technology, 2012, 31(2):66- 71.
[42] Tong Yan, Ma M R, Lin Q, Wang Q, You W H. Dynamics of spatial and temporal distribution of phytoplankton and related affecting factors in the dishui lake. Resources and Environment in the Yangtze Basin, 2012, 2012, 21(12): 1499- 1506.
[43] Xu J G. Situation and protection of water quality in Shanghai dishui lake. The Administration and Technique of Environmental Monitoring, 2010, 22(1): 64- 66.
参考文献:
[1] 刘水芹, 田华, 梁国康. 大型人工湖泊生态环境演变分析及对策探讨. 上海水务, 2009, 25(3): 25- 29, 33- 33.
[2] 汪海英, 周敏杰. 临港新城——滴水湖富营养化现状评价及调控对策. 上海水务, 2006, 22(4): 24- 26, 28- 28.
[3] 孙刚, 郎宇, 房岩. 长春南湖水生生态系统中浮游动物群落特征. 吉林大学学报: 理学版, 2006, 44(4): 663- 667.
[4] 王延洋. 滴水湖浮游动物群落结构及水质生态学评价 [D]. 上海: 上海师范大学, 2008.
[5] 周晓梅. 滴水湖及其外围水体浮游动物群落结构比较研究[D]. 上海: 上海师范大学, 2010.
[6] 江敏, 任治安, 邢斌, 卢柳, 胡文婷, 陈立婧, 李娟英. 上海滴水湖水系轮虫群落结构分析及水质评价. 科技导报, 2012, 29(22): 31- 37.
[7] 田华, 刘水芹, 方伟, 边佳胤, 李娟英. 人工湖泊滴水湖水质演变趋势及富营养化分析. 水生态学杂志, 2012, 32(6): 26- 31.
[8] 周凤霞, 陈剑虹. 淡水微型生物图谱 (第二版). 北京: 化学工业出版社, 2010: 303- 303.
[9] 章宗涉, 黄祥飞. 淡水浮游生物研究方法. 北京: 科学出版社, 1991: 232- 232, 256- 258.
[10] 王家楫. 中国淡水轮虫志. 北京: 科学出版社, 1961: 20- 21.
[11] 蒋燮治, 堵南山. 中国动物志(节肢动物门甲壳纲淡水枝角类). 北京: 科学出版社, 1979: 43- 74.
[12] 沈嘉瑞. 中国动物志(节肢动物门甲壳纲淡水桡足类). 北京: 科学出版社, 1979: 27- 27, 31- 36
[13] 王延洋, 李晓波, 吴波, 吴琼, 许夏玲, 王全喜. 上海滴水湖浮游动物研究初报. 上海师范大学学报: 自然科学版, 2008, 37(2): 167- 172.
[14] 何玮, 薛俊增, 吴惠仙. 滩涂围垦湖泊(上海滴水湖)轮虫的群落结构与时空分布. 湖泊科学, 2012, 23(2): 257- 263.
[18] 王站付, 周晓梅, 王延洋, 罗永婷, 王全喜. 上海市滴水湖浮游动物的群落结构及动态. 上海师范大学学报: 自然科学版, 2011, 40(3): 295- 300.
[19] 刘振宇, 徐建平. “十一五”期间滴水湖富营养化评价. 环境科学与管理, 2012, 37(4): 169- 172.
[20] 金相灿, 屠清瑛. 湖泊富营养化调查规范 (第二版). 北京: 中国环境出版社, 1990.
[21] 张觉民, 何志辉. 内陆水域渔业自然资源调查手册. 北京: 中国农业出版社, 1991.
[22] 王凤娟, 胡子全, 汤洁, 刘连生, 赵海泉. 用浮游动物评价巢湖东湖区的水质和营养类型. 生态科学, 2006, 25(6): 551- 552.
[23] 冯德祥, 刘一, 禹娜, 陈亮, 李二超, 陈立侨. 淀山湖后生浮游动物群落结构特征分析. 华东师范大学学报: 自然科学版, 2011, (6): 126- 128.
[24] 史文. 千岛湖浮游动物群落结构的时空变化及其与影响因子的相关性研究 [D]. 上海: 上海海洋大学, 2010.
[25] 任治安. 滴水湖及其引水河道主要浮游动物群落研究 [D]. 上海: 上海海洋大学, 2012: 12- 13.
[29] 陈光荣, 钟萍, 张修峰, 谢贻发, 李传红. 惠州西湖浮游动物及其与水质的关系. 湖泊科学, 2008, 20(3): 351- 356.
[30] 吴建新, 阎斌伦, 冯志华, 李玉, 徐加涛, 李士虎, 申欣. 田湾核电站海域浮游动物生态特征. 生态学报, 2011, 31(22): 6902- 6911.
[31] 刘一. 淀山湖浮游动物群落结构研究 [D]. 上海: 华东师范大学, 2010.
[32] 王庆, 刘丽君, 杨宇峰. 南方某水库轮虫群落特征及其对环境因子的响应. 生态学报, 2010, 30(13): 3385- 3395.
[33] 邱小琮, 赵红雪, 孙晓雪. 鸣翠湖轮虫群落特征及其与水环境因子的关系. 宁夏大学学报: 自然科学版, 2012, 33(1): 62- 68.
[37] 黄祥飞, 胡春英, 伍悼田. 武汉东湖的轮虫. 水生生物学报, 1985, 9(2): 129- 143.
[38] 刘健康. 东湖生态学研究(一). 北京: 科学出版社, 1990.
[39] 霍元子, 何文辉, 罗坤, 王阳阳, 张饮江, 田千桃, 何培民. 大型溞引导的沉水植被生态修复对滴水湖水质的净化效果. 应用生态学报, 2010, 21(2): 495- 499.
[40] 童琰, 徐春燕, 胡雪芹, 张维砚, 由文辉. 滴水湖引水河段组合型生态工程春季对水体净化效果研究. 上海海洋大学学报, 2011, 20(6): 930- 937.
[41] 周新龙. 综合水质指数法对滴水湖水质的评价. 净水技术, 2012, 31(2): 66- 71.
[42] 童琰, 马明睿, 林青, 王强, 由文辉. 滴水湖浮游植物时空分布动态及其影响因子. 长江流域资源与环境, 2012, 21(12): 1499- 1506.
[43] 徐建官. 滴水湖水质现状及保护初探. 环境监测管理与技术, 2010, 22(1): 64- 66.
