文冠果花性状多尺度分析1)
2014-09-18杨韫嘉
徐 欢 杨韫嘉 樊 简
(北京林业大学,北京,100083)
刘春和 徐红江
(北京市大东流苗圃)
石春鸿
(北京青神园林工程有限公司)
关文彬
(北京林业大学)
尺度问题是所有生态学研究的基础[1]。尺度包括现象尺度、观测尺度、分析尺度[2]。对现象的研究是基于一定的观测和分析尺度下进行的。因此,选取不同的观测和分析尺度,将检测到不同的现象[3]。尺度研究根本目的在于通过适宜的观测和分析来揭示和把握不同尺度中现象的规律性[4]。应用尺度方差分析法,可以观察某一变量的尺度方差随分析尺度的增大是否发生转折,这不仅可以检测该变量在多尺度上的空间变异性,也可以检测和描述该变量的等级结构特征[5]。性状变异是基因型与环境共同作用的结果,在一定程度上反映物种遗传变异的程度,可以揭示植物与环境之间的关系,有助于认识植物适应和进化的方式、机制及其影响因素[6-7]。由于对空间多样性环境的响应,花性状是否会随环境因子的变化而变化呢?
文冠果(Xanthoceras sorbifolium Bunge.)为无患子科单型属,中国特有种,为第三纪孑遗种[8],是被子植物中的“活化石”,有“北方油茶”之称,被誉为中国北方最有潜力的能源植物之一[9];同时也是珍稀的药用树种、优美的城市绿化树种和重要的绿化先锋树种以及优良食品与特种用材树种[10]。关于文冠果栽培技术的研究已开发出育苗、定植技术、保花、保果、增产的果园管理技术[11];繁殖技术也有突破,针对落花落果、雄性不育等方面问题,从细胞学、解剖学、生理学、生物化学和分子生物学角度进行了深入探讨[12-18]。目前,还没有文冠果的品种登记,根据性状表型不同将嫩梢和叶部是否有毛可分为两种类型,并认为无毛文冠果是丰产类型[19]。性状选优的研究仅从单一的表型性状进行选择[20],尚未达到优良类型选择的水平[21]。研究证实,同一分布区内文冠果花表型性状在植株间变异较大[19],而多尺度分析性状差异尚未有报道。植物新品种DUS(特异性、一致性、稳定性 )是新品种的本质特征,为了开展建立文冠果DUS性状指标体系与判别方法的需要,文中以北京和辽宁地区的文冠果花性状为研究对象,从多尺度分析性状差异、认识性状特征、揭示性状间的关系,并构建性状模型,为文冠果品种鉴定与分类,提供花性状的数据与依据。
1 研究地概况
研究地区包括北京市昌平区大东流苗圃基地和位于辽宁省西部朝阳市北端的建平县。北京昌平全区地形东、南低,为平原;西、北高,为山区,平原海拔高度30~100 m。辽宁建平地处燕山山脉向辽沈平原的过渡地带,为辽西山地丘陵区。辽宁的年平均气温较北京低3.4℃,北京和辽宁的其他气象因子差异也很大(表1)。

表1 北京、辽宁气象因子
2 材料与方法
试验于文冠果盛花期,花型饱满时对两地点文冠果花性状进行测定,北京地区采样时间为4月下旬,辽宁地区采样时间为5月初。两地区文冠果物候期相差7~8 d,由于文冠果花期较短,在北京地区选取4株文冠果植株(其中3株白花类型、1株红花类型)、辽宁地区选取7株文冠果植株(其中6株白花类型、1株红花类型),所选的11株具有代表性的无病虫灾害单株,均可最大限度地降低采样树间的遗传差异,以保证性状的可比性。所选对象的表型性状都是序柄色为绿色、新生枝为红色、枝条嫩梢无毛。因而,基于目前文冠果尚没有品种认定,加之实生植株的遗传分化较大等原因,对上述取样作如下假设:两种花色为不同的遗传基因型;同一花色具有新生枝无毛等相同性状特征为相近的基因型。
取样设计和性状指标的测定:为了从不同尺度对文冠果花朵性状和花序性状进行分析,设计了单花—花序—植株—区域4种尺度进行取样。在参试植株朝南方位上随机选取1个枝条,在枝条的上、中、下3个部位各随机选取1个花序为研究对象,进行花序性状指标的测定。因此,北京地区共选取9个白花花序和3个红花花序;辽宁地区共选取18个白花花序和3个红花花序。将每个取样花序中的花朵作为研究对象,进行花朵性状指标的测定。在北京地区共测定了195个白花花朵和61个红花花朵;在辽宁地区共测定了361个白花花朵和51个红花花朵。由于花期时间的限制、加之红花植株数量极少所限,调查取样已经是最大程度的取样。
测定的9个花序性状指标为:花序基径、花序长、花序宽(花序最宽处的值)、花序质量、序宽/序长、序基径/序宽和每个花序上的雄花数、两性花数、小花数。其中,花序质量是将花序置于 80℃恒温下烘干48 h至恒质量。测定的9个花朵性状指标为:不育花药数、花药总数、花长、花柄长、冠径(花朵盛开时的自然直径)、花瓣长、花瓣宽、瓣宽/瓣长、朵径长比,其中,朵径长比=冠径/花长。花瓣长和花瓣宽的测量方法是依次用镊子将所有花瓣分开,固定到纸板上,用游标卡尺测量每个花瓣的长度和宽度,游标卡尺的测量精度为0.1 mm。
数据分析:为了探讨花朵性状在花序尺度上的差异,则分别对北京和辽宁两地文冠果花序之间的花朵性状进行单因素方差分析;为了探讨花朵性状和花序性状在植株尺度上的差异,采用单因素方差分析分别对北京和辽宁两地各文冠果植株之间的花朵和花序性状进行检验;为了探讨花色和地区对花朵性状和花序性状的影响,采用双因素多变量方差分析分别对花朵和花序性状进行差异检验;而为了探讨花朵性状和花序性状在地区尺度上的差异,采用独立样本T检验来检验北京和辽宁地区之间花朵性状和花序性状的差异。对18个花(花朵和花序)性状采用Pearson相关性分析和线性回归分析确定性状相关关系,数据处理通过Excel和SPSS18.0软件完成。
3 结果与分析
3.1 性状差异分析
花朵尺度性状分析:对两地区花序间不同花色的9个花朵性状进行单因素方差分析(表2)表明,白花花朵性状在不同花序间表现出不同的差异性。其中,北京和辽宁两地,白花花序间的8个花朵性状均呈极显著性差异(P<0.001),而花药总数北京呈显著差异,辽宁无显著差异(P>0.05)。
红花在两地区的花序间花朵性状的差异性与白花不同。北京地区,红花的花药总数、花长和朵径长比呈极显著差异,冠径呈显著差异;这4个性状在辽宁均表现为无显著性差异。花瓣长和瓣宽/瓣长在两地区表现一致,前者均无显著性差异,后者呈极显著差异。红花不育花药数和花柄长在北京都表现为极显著差异,但这两个性状在辽宁均表现为显著差异。除红花的花瓣宽性状外,其他白花和红花的花朵性状都表现为北京地区的显著性大于辽宁地区(表2)。
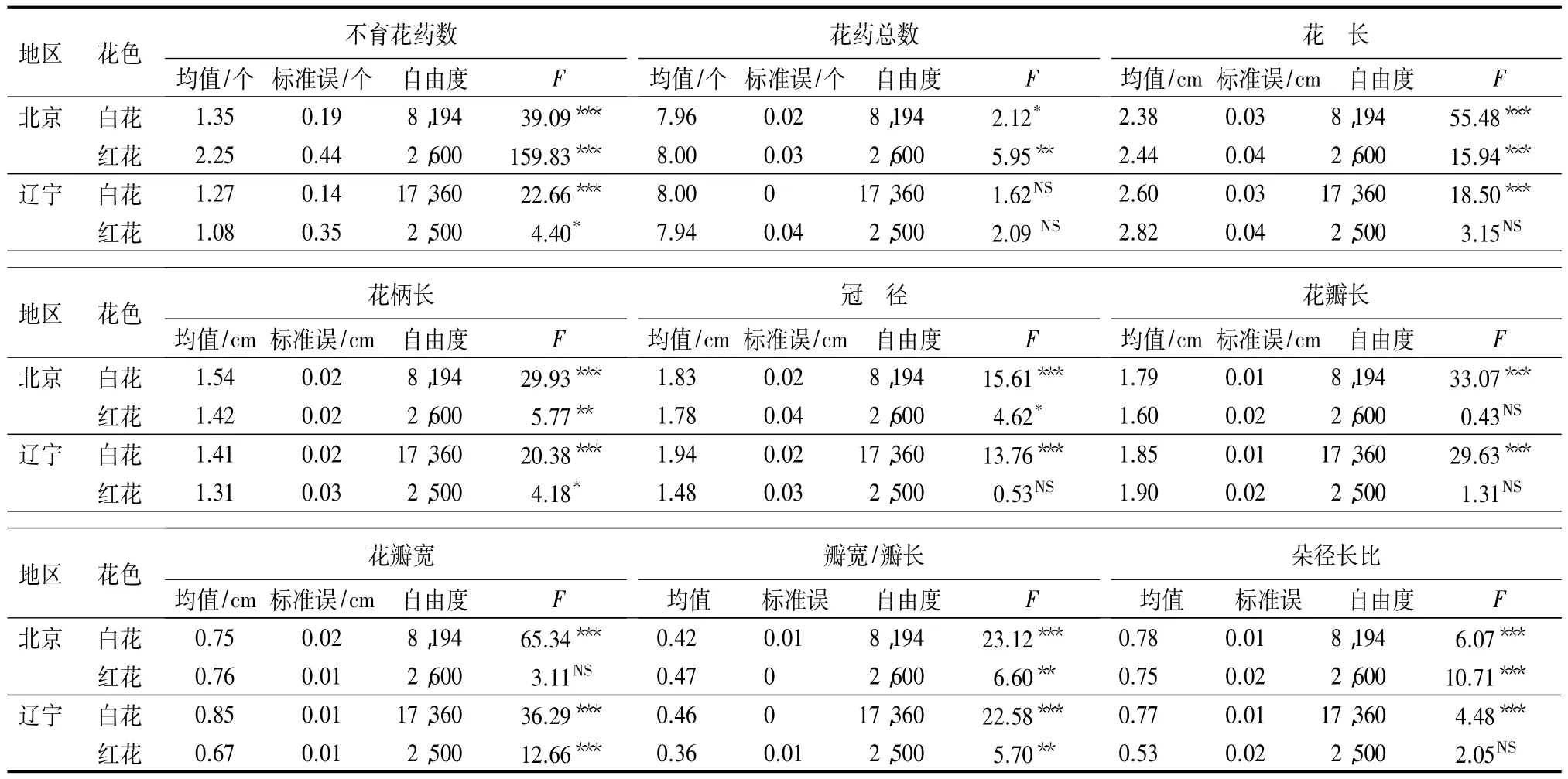
表2 两地区花序间不同花色花朵性状的单因素方差分析
花序尺度性状分析:两地区植株间白花花序性状单因素方差分析(表3)表明,白花花序性状除雄花数外,其他8个花序性状在两地区植株间都表现一致,只有花序宽在植株间有极显著差异,其余7个性状差异均不显著。北京地区植株间雄花数表现为显著性差异,辽宁地区该性状无显著性差异。
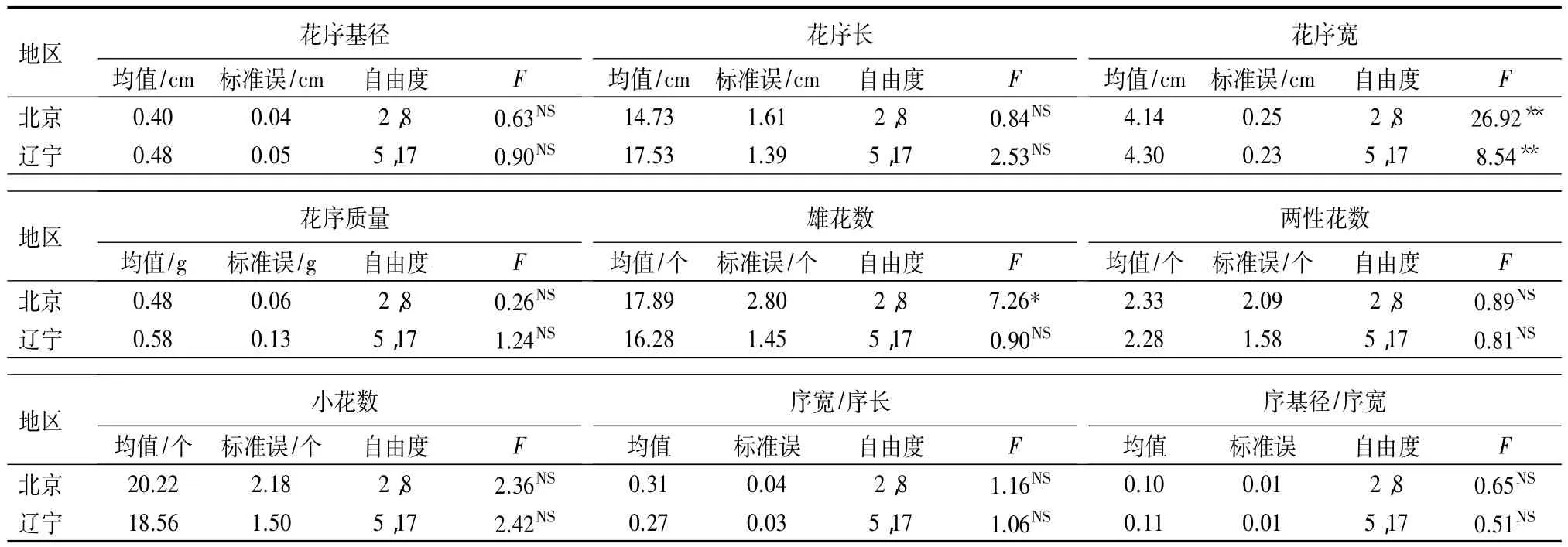
表3 两地区植株间白花花序性状的单因素方差分析
植株尺度性状分析:两地区植株间白花花朵性状方差分析表明(表4),8个花朵性状在两地区的表现一致,植株间均为极显著差异。花药总数在北京地区表现为差异极显著,而在辽宁地区无显著差异。花药总数在两地区植株间的表现与两地区花序间的表现不一致,显著性程度不同,其他性状在两个尺度上均表现为极显著差异。两地区植株间白花花序性状的方差分析见表3,两地区植株间红花花性状的方差分析与地区尺度红花花性状的方差分析相同(表5)。
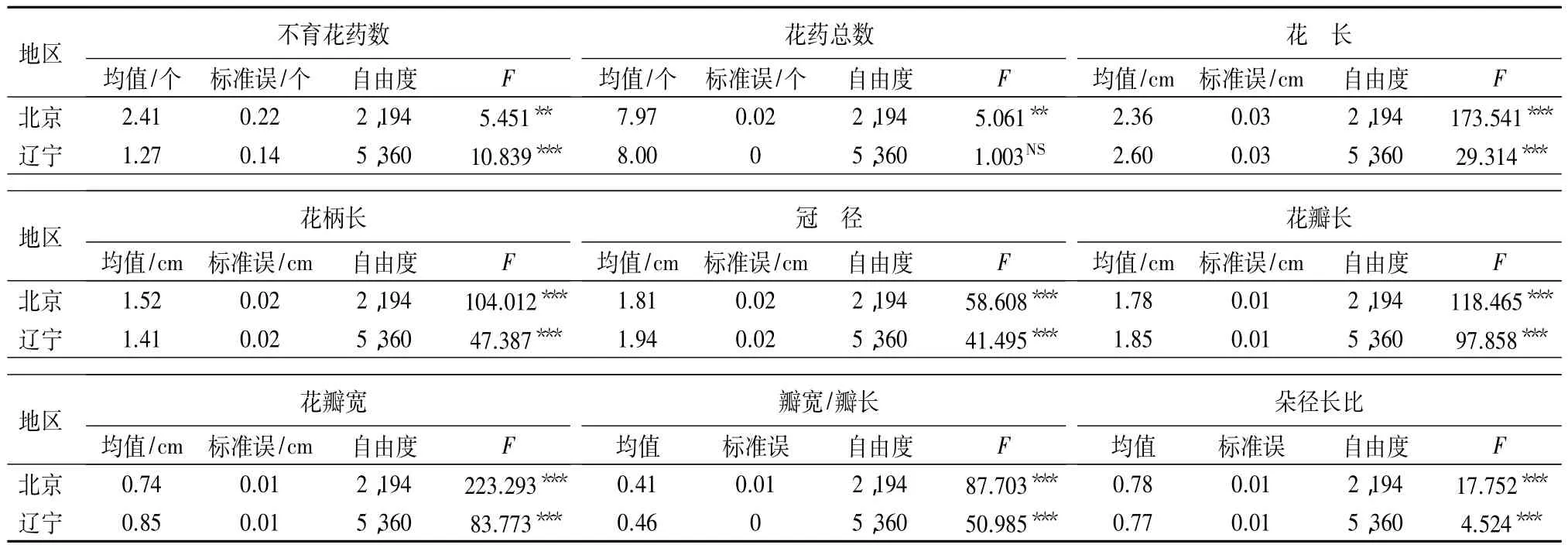
表4 两地区植株间白花花朵性状的单因素方差分析
区域尺度性状分析:两种花色、两地区花性状的多因素方差分析(表5)可知,白花与红花二者间,花朵性状只有不育花药数、花药总数两个性状差异不显著;其余7花朵性状差异极显著,而二者花序性状无显著差异。
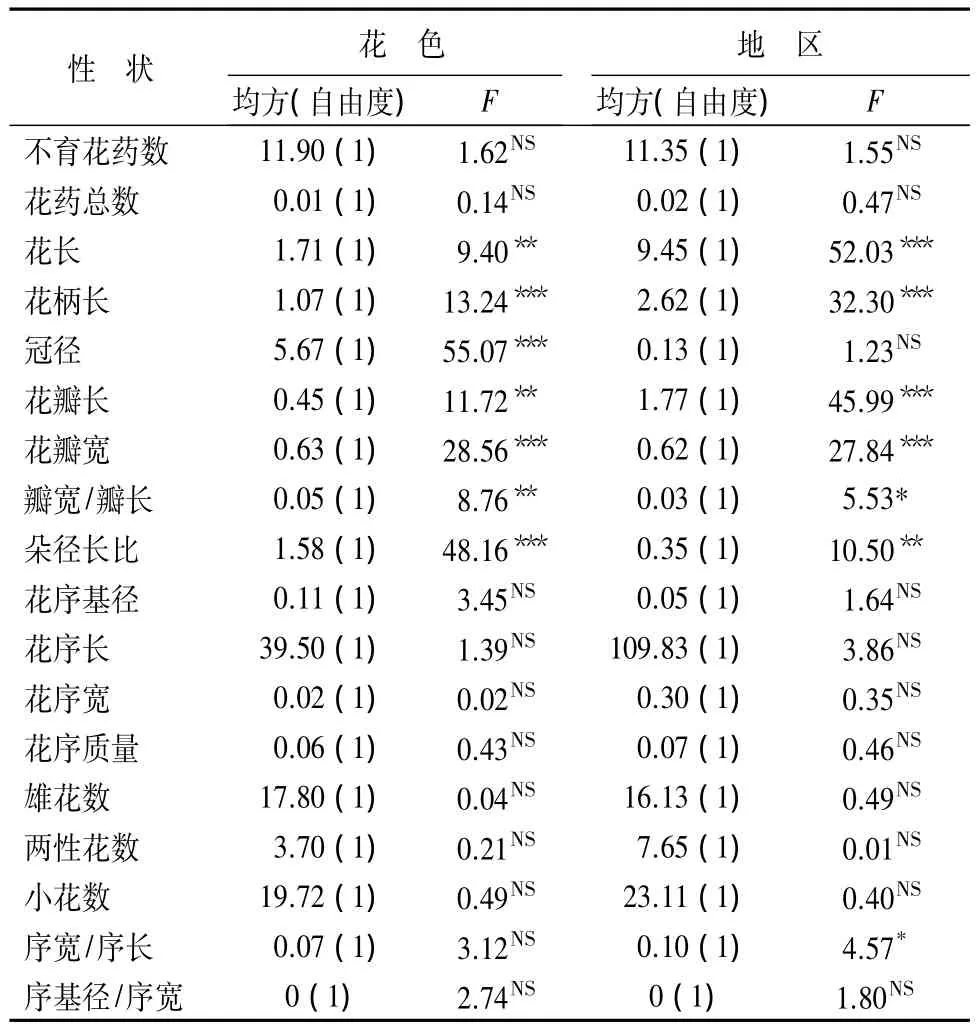
表5 两花色两地区花性状的双因素多变量方差分析
两地区之间,花朵性状中不育花药数、花药总数和冠径3个性状差异不显著,其他6个性状差异显著,其中花长、花柄长、花瓣长、花瓣宽和朵径长比5个性状差异极显著;花序性状中序宽/序长差异显著,这点与白花和红花二者均不同。
进一步对两地区不同花色花性状进行独立样本T检验分析(表6)可知,花朵性状的差异性大于花序性状,红花花朵性状的差异大于白花花朵性状。例如,白花花朵性状中有5个性状(花长、花柄长、冠径、花瓣宽、瓣宽/瓣长)达到极显著水平。红花花朵性状中有6个性状(花长、冠径、花瓣长、花瓣宽、瓣宽/瓣长和朵径长比)达到极显著水平,2个性状(不育花药数、花柄长)达到显著水平。花序性状中,只有红花的花序宽呈显著差异,其他花序性状均无显著性差异。不同花色花朵性状在两地区数值上有差异,有4个白花花朵性状是北京地区数值小于辽宁地区,分别为花长、冠径、花瓣宽和瓣宽/瓣长,1个白花性状(花柄长)是北京地区数值大于辽宁,差异均达到极显著水平。红花花朵性状中,有4个性状是北京地区数值大于辽宁地区,分别为冠径、花瓣宽、瓣宽/瓣长和朵径长比,2个性状是北京地区数值小于辽宁,分别为花长和花瓣长,差异也达到极显著水平(表6)。
3.2 性状相关性与回归分析
花性状的相关性分析:为研究花性状间的相关程度,对两地区的18个性状进行Pearson相关性分析(表7~表10)。结果表明:白花花朵性状在北京地区和辽宁地区性状相关性表现不一致,例如,北京白花的不育花药数与花瓣长呈极显著负相关,与花瓣宽和朵径长比呈显著负相关;而辽宁的不育花药数与朵径长比呈极显著负相关,与花药总数呈显著负相关,与花长呈极显著正相关,与花瓣宽呈显著正相关,花朵性状相关性在两地区表现不同(表7)。红花花朵性状在北京和辽宁的性状相关性也不一样,例如,北京红花的不育花药数与花长呈极显著正相关;辽宁红花不育花药数与花长无显著相关性(表8)。红花花朵性状间相关性情况与白花花朵性状有差异,白花不育花药数与花长在北京无显著相关性;而在辽宁有正相关性。白花冠径与花瓣长、花瓣宽在两地区都是在0.01水平上显著正相关,表现一致;红花冠径与花瓣长、花瓣宽在北京呈极显著、显著正相关,在辽宁没有显著相关性。

表6 同一花色两地区花性状的T检验
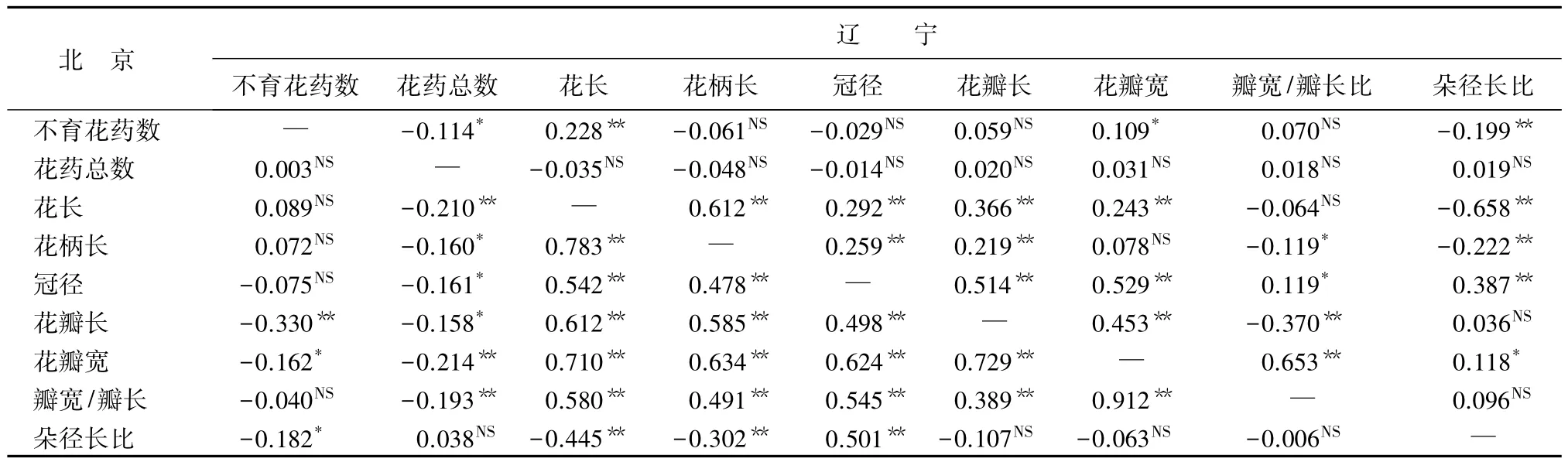
表7 两地区白花花朵性状间相关系数
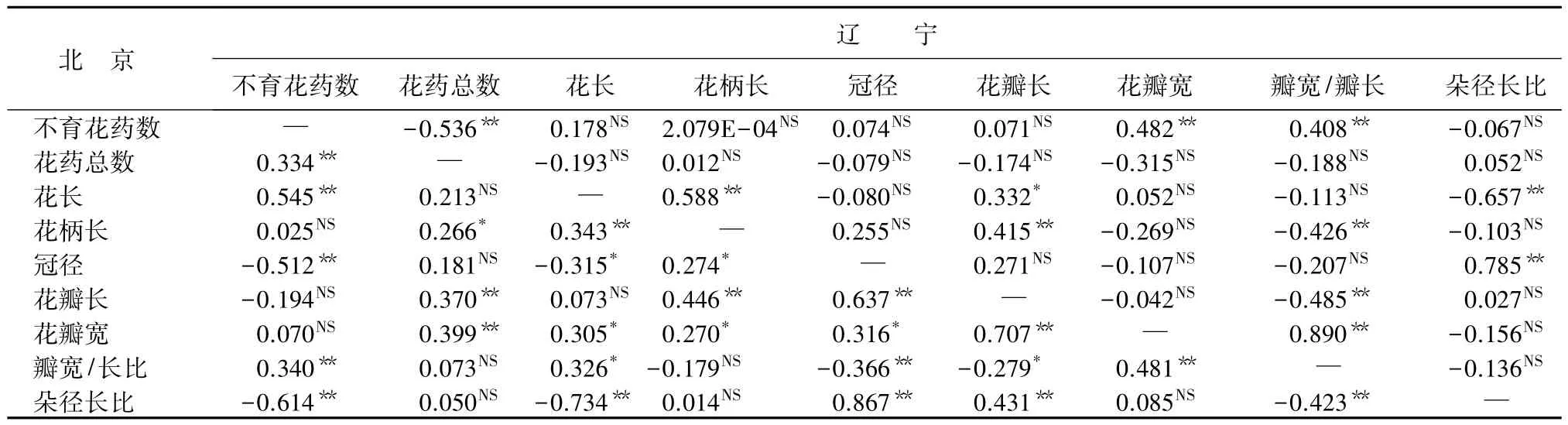
表8 两地区红花花朵性状间相关系数
北京白花花序基径与序基径/序宽呈极显著正相关,与其他花序性状无显著相关性;辽宁白花花序基径与花序长、花序质量、两性花数、小花数、序基径/序宽呈极显著正相关,与序宽/序长呈显著负相关(表9)。从表10中可以看到,北京和辽宁的红花花序基径与花序长、花序宽、雄花数、两性花数、小花数、序宽/序长无显著相关性,两地区表现一致,辽宁红花花序基径与花序质量和序基径/序宽分别呈显著、极显著正相关。
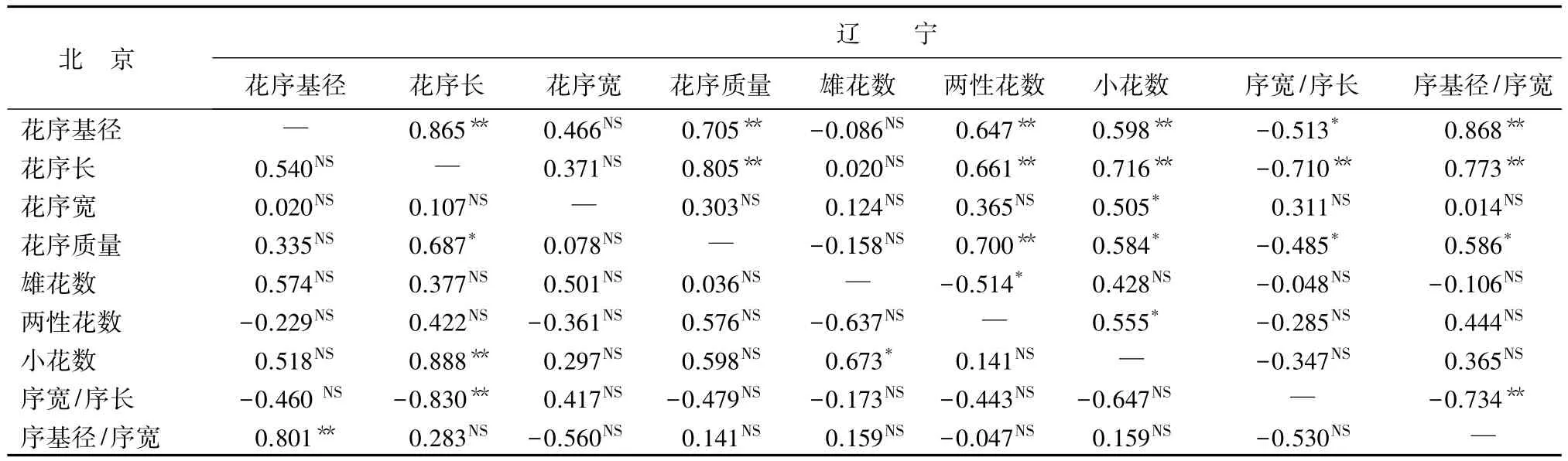
表9 两地区白花花序性状间相关系数
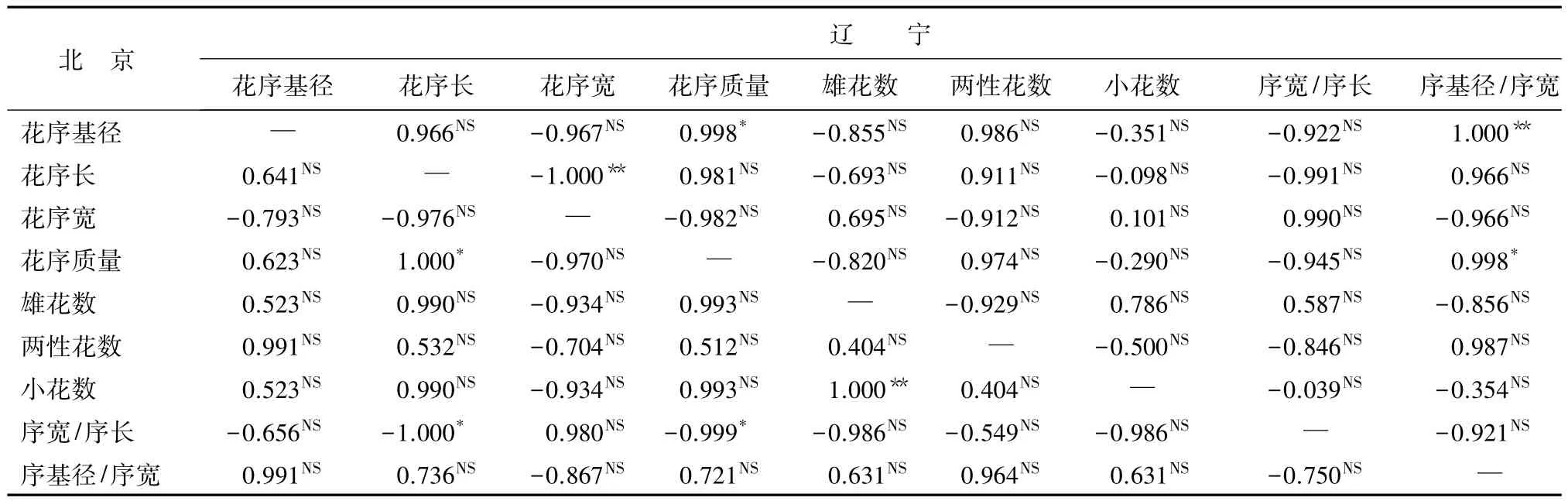
表10 两地区红花花序性状间相关系数
花性状的回归分析:根据上述相关分析,借鉴生物量模型,对花冠面积(A)、花序质量(B)与其他性状进行回归分析,优化筛选,给出回归估计模型。花冠面积(A)与花瓣长(X1)、花瓣宽(X2)的回归估计模型为:

如图1所示,根据公式(1)给出白花两地区花冠面积估计模型:

式中:AB是北京白花花冠面积;AL是辽宁白花花冠面积;XB是北京白花花瓣长与花瓣宽的函数式;XL是辽宁白花花瓣长与花瓣宽的函数式。
图1 两地区白花花瓣长、宽和花冠面积的关系
如图2所示,根据公式(1)给出红花两地区花冠面积估计模型:
式中:A'B是北京红花花冠面积;A'L是辽宁红花花冠面积;X'B是北京红花花瓣长与花瓣宽的函数式;X'L是辽宁红花花瓣长与花瓣宽的函数式。

图2 两地区红花花瓣长、宽和花冠面积的关系
花序质量与花序基径 (D1)的回归估计模型为:

根据公式(2)给出两地区白花花序质量关于花序基径的一元估计模型(图3):

图3 两地区白花花序基径与花序质量的关系
花序质量(B)和花序长 (D2)的回归估计模型为:

根据公式(3)给出两地区白花花序质量关于花序长的一元估计模型(图4):
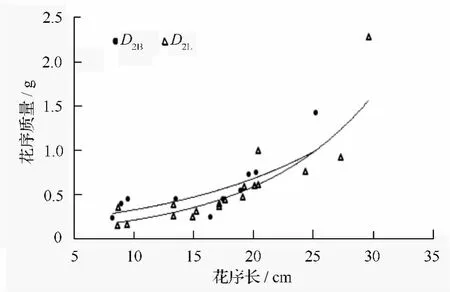
图4 两地区白花花序长与花序质量的关系
花序质量(B)与花序基径(D1)、花序长(D2)、花序宽(D3)的回归估计模型:

根据公式(4)给出两地区白花花序质量关于花序基径、花序长和花序宽的三元估计模型(图5):
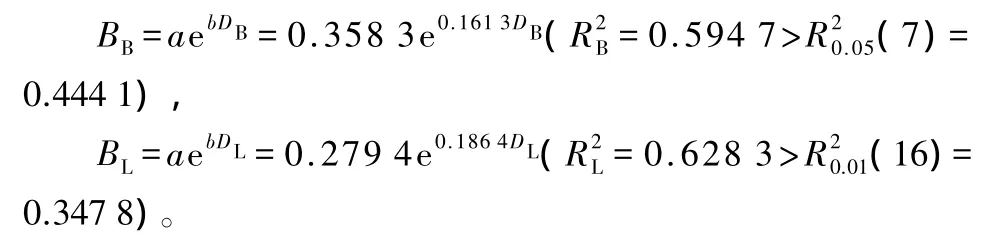
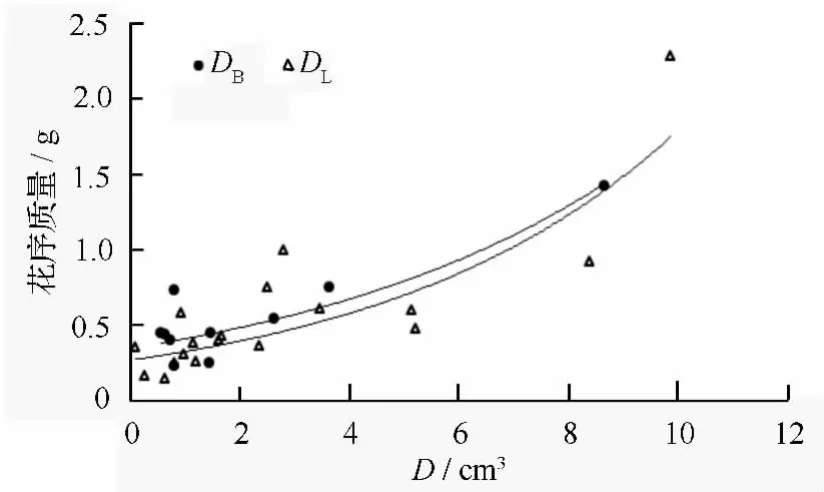
图5 两地区白花花序基径、长、宽与花序质量的关系
4 结论与讨论
植物性状反映了植物对气候和土壤等环境因子适应的重要信息[22]。文冠果性状受其生长的地理位置、立地条件及小气候的影响,使其在不同生境下的相关变异也不同[19]。采用相同的遗传型作为供试对象,进行单花—花序—植株—区域多尺度分析可以得出明显的规律。
花性状的遗传变异大于尺度差异,但主要表现在单花尺度而不是花序以上尺度,红花变异大于白花。白花与红花二者间,花朵性状只有不育花药数、花药总数两个性状差异不显著;其余7花朵性状差异极显著,而二者花序性状无显著差异。两地区之间有6个花朵性状差异显著,除瓣宽/瓣长外,其他5个性状差异极显著;花序性状的序宽/序长差异显著,这点和白花与红花二者遗传之间的差异有所不同。
白花文冠果花性状的差异,两地区花朵性状的不育花药数、花药总数、花瓣长和朵径长比等4个性状无显著差异,花长、花柄长、冠径、花瓣宽和瓣宽/瓣长5个性状差异极显著;花序性状无显著差异。单花尺度,北京地区花药总数差异显著,辽宁地区花药总数在花序—植株—区域3个尺度的分析中均为无显著差异,北京地区存在花药不发育现象;植株尺度上,花序性状的花序宽两地均有极显著差异;雄花数北京地区差异显著,其他性状均无显著差异。
红花文冠果花性状的差异,两地只有花药总数无显著差异,其余8个性状差异显著;花序宽差异显著。北京地区不育花药数和花柄长在花序尺度上有极显著差异,在区域尺度上具有显著差异,而辽宁地区不育花药数和花柄长在两个尺度上都表现为显著差异。北京花朵性状中花瓣长、花瓣宽无显著差异,辽宁花朵性状中花长、冠径、花瓣长、朵径长比两地均无显著差异。
花性状不同尺度下花性状变异程度由大到小的排序为单花尺度、花序尺度、植株尺度、区域尺度。白花花朵性状在花序、植株尺度上有显著差异的性状数一致,区域尺度有差异的性状减少;花序性状比花朵性状稳定,植株尺度上,除白花花序宽和雄花数有差异外,其他性状差异不显著,区域尺度上,所有性状均无显著差异。但红花花序宽在区域尺度上表现为显著差异。性状变异反映了群体遗传稳定性与环境复杂性的相互关系及其适应环境压力的广泛程度[23],这种多层次的变异成为种质选择的源泉[23-26],而基于多尺度分析,会得出更明确的规律。
植物的形态特征是在环境中长期适应和进化的结果,各个形态性状间往往存在一定联系,一个性状的变异可能导致其他相关性状的变异[27]。
通过对9个花朵性状和9个花序性状相关性分析发现,花朵性状和花序性状间的相关性都是白花大于红花,但花序尺度相关性程度远低于花朵尺度,区域尺度上花朵性状间相关性是北京大于辽宁,花序性状间是辽宁大于北京。花朵性状间的相关性,白花性状间相关性大于红花,北京地区性状间相关性大于辽宁地区。9个花朵性状中,白花的形态性状花长、花瓣宽、花瓣长与其他性状相关性更密切,北京地区是花瓣宽、辽宁是花长与其他性状最密切;红花的则是分配性状朵径长比、瓣宽/瓣长与其他性状相关性更密切,北京远高于辽宁的。花序性状间的相关性也是白花的大于红花的,但远低于花朵尺度,而区域尺度上辽宁大于北京。9个花序性状中,白花在北京地区仅有花序长与其他性状相关性密切,辽宁地区形态性状的花序基径、花序长与结构性状花序质量和其他性状相关性更密切。红花仅仅是辽宁的序基径/序宽,而北京的是花序长、序宽/序长与其他少量性状显著相关,如花序质量。
性状的相关性差异进一步验证了前述的性状方差分析与差异检验的结论。值得强调的是,红花与白花相比是为了吸引更多的访花昆虫的进化表现,从这种分配性状与其他形态性状有更高的相关性可以推断,红花具有对环境更好的适应性,这有待通过分子数据深入研究。
根据性状相关分析结果,首次构建了花冠面积与花序质量估计模型。花冠面积与(花瓣长、花瓣宽)呈二元指数模型;花序质量与花序基径(D1)、花序长(D2)呈一元指数方程,与(花序宽D3)呈三元指数方程。这些模型的性状,正是前述相关程度高的性状,而这种结构性状指标与形态性状指标的回归关系,奠定了性状评价的重要理论基础。两种花色的估计模型一致,但两种花色方程的参数不同充分反映其遗传差异。给出的模型可用于性状评价,简化复杂性状调查等。
目前,文冠果由于长期处于野生状态,并且粗放的管理和人们大量砍伐造成早期选育的优良单株无法查起,所以文冠果选育优良单株的工作迫在眉睫[28-29]。文冠果优良单株的选择还需要从树型、枝型、芽型、果实、种子、SSR分子标记等性状进行全面筛选,建立性状评价指标体系,进而建立DUS测试指南的性状指标体系,构件判别准则;采用定量与定性相结合的方法,建立判别标准,对优良类型、优良无性系进行数量分类研究。
[1]Wiens J A.Spatial scaling in ecology[J].Functional Ecology,1989,4(3):385-397.
[2]Wu Jianguo,Bruce J K,Li Harbin,et al.Scaling and uncertainty analysis in ecology[M].New York:Columbia University Press,2006:3-16.
[3]张娜.生态学中的尺度问题:内涵与分析方法[J].生态学报,2006,26(7):2340-2355.
[4]吕一河,傅伯杰.生态学中的尺度及尺度转换方法[J].生态学报,2001,21(12):2096-2105.
[5]Wu Jianguo,Jelinski D E,Luck M,et al.Multiscale analysis of landscape heterogeneity:scale variance and pattern metrics[J].Geographic Information Sciences,2000,6(1):6-19.
[6]王娅丽,李毅,陈晓阳.祁连山青海云杉天然群体表型性状遗传多样性分析[J].林业科学,2008,44(2):70-77.
[7]钱迎倩,马克平.生物多样性研究的原理方法[M].北京:中国科学技术出版社,1994.
[8]徐东翔.文冠果落果机理及提高座果率和种子品质的研究[J].西北植物学报,1990,10(2):117-127.
[9]王涛.中国主要生物质燃料油木本能源植物资源概况与展望[J].科技导报,2005,23(5):12-14.
[10]毕泉鑫,蔡龙,马兴华,等.中国特有能源植物文冠果的遗传学及产业化[J].中国野生植物资源,2011,30(5):37-41.
[11]白金友.文冠果增产技术的研究[J].辽宁林业科技,1988(5):53-57.
[12]Zhang Na,Guo Jinping,Zhang Yunxiang,et al.Plant regeneration via direct organogenesis of Xanthoceras sorbifolia bunge[J].Propagation of Ornamental Plants,2012,12(1):63-71.
[13]董亚芳.文冠果优良单株选择及光合特性的初步研究[D].郑州:河南农业大学,2010.
[14]杜希华,陆海,高述民,等.文冠果雄性可育性相关cDNA片段的克隆与序列分析[J].北京林业大学学报,2003,25(5):29-33.
[15]刘波,王力华,阴黎明,等.两种林龄文冠果的生长和结实特性[J].中国科学院研究生院学报,2011,28(1):73-79.
[16]刘波,王力华,阴黎明,等.两种林龄文冠果叶N、P、K的季节变化及再吸收特征[J].生态学杂志,2010,29(7):1270-1276.
[17]彭伟秀,李凤兰,王保柱.文冠果同源异型变异株的发现[J].北京林业大学学报,1999,21(5):92-94.
[18]彭伟秀,沈昕,李凤兰,等.文冠果变异株和野生型植株基因组差示杂交研究初报[J].北京林业大学学报,2000,22(1):29-32.
[19]牟洪香.木本能源植物文冠果(Xanthoceras sorbifolia Bunge)的调查与研究[D].北京:中国林业科学研究院,2006.
[20]高述民,马凯,杜希华,等.文冠果(Xanthoceras sorbifolia)研究进展[J].植物学通报,2002,19(3):296-301,289.
[21]芦娟,柴春山,蔡国军,等.甘肃定西文冠果花的表型多样性研究[J].林业资源管理,2011(6):49-53.
[22]孟婷婷,倪健,王国宏.植物功能性状与环境和生态系统功能[J].植物生态学报,2007,31(1):150-165.
[23]姚淑均,张守攻,王军辉,等.滇楸花部性状的表型多样性研究[J].中南林业科技大学学报,2013,33(10):19-24.
[24]赵冰,张启翔.蜡梅种质资源表型多样性[J].东北林业大学学报,2007,35(5):10-13,35.
[25]黄佳聪,何俊,尹瑞萍,等.腾冲红花油茶自然和人工种群种实性状变异研究[J].北京林业大学学报,2010,32(5):94-101.
[26]李斌,顾万春,卢宝明.白皮松天然群体种实性状表型多样性研究[J].生物多样性,2002,10(2):181-188.
[27]顾婧婧,金则新,熊能.濒危植物夏蜡梅花的形态变异[J].植物研究,2010,30(4):461-467.
[28]敖妍.因子分析法在文冠果优良单株选择中的应用[J].华南农业大学学报,2009,30(4):70-73.
[29]牟洪香,侯新村.文冠果的研究进展[J].安徽农业科学,2007,35(3):703-705.
