林分密度对水曲柳人工林吸收根生物量和根长密度的影响1)
2014-09-18谢玲芝李俊楠王韶仲王文娜谷加存
谢玲芝 李俊楠 王韶仲 王文娜 谷加存
(东北林业大学,哈尔滨,150040)
细根(直径≤2.0 mm)是树木吸收水分和养分的主要器官,对其生长发育起决定性的作用[1-2]。林分密度是指单位面积林地上的林木株数,它不仅决定了林木个体的生长空间和发育过程,而且对森林生产力产生重要的影响[3]。然而,关于林分密度与细根生物量和根长之间关系的研究仍较少。一些研究指出细根生物量随林分密度增加而增加[4-5],如徐程扬等[6]发现侧柏(Platycladus orientalis)细根(直径≤1.0 mm)根长密度随林分密度的的增加而显著增大;另一些研究发现随林分密度的增大细根生物量减小[7-8],如田宇明等[9]对 10 年生水曲柳(Fraxinus mandshurica)人工林研究发现,细根(直径≤2.0 mm)生物量随林分密度的增加而显著降低;还有一些研究发现细根生物量与林分密度不是线性关系[6,10-11]。显然,树种、立地、林龄、密度梯度等差异可能会导致研究者获得的结论不同[12-13]。然而,以往研究在很大程度上忽视了根系内在结构和功能异质性对研究结果的影响[14]。
长期以来,国内外的研究都将根系简单的分为细根(直径≤2.0 mm)和粗根(直径>2.0 mm),认为直径≤2.0 mm的根具有相同的生理生态功能(如资源吸收),但近10年来的研究表明这样的认识过于简单。Pregitzer等[15]对北美9个树种的研究发现,细根的比根长和氮浓度随着根序(最末端的为1级根,1级根长在2级根上,依此类推)的增大而降低,推测不同根序细根生理生态功能(如维持呼吸与资源获取)存在很大差异。Guo等[14]研究进一步发现,不同根序细根在解剖结构上存在显著差异,低级根(前3级根)具有皮层组织,是主要的吸收根,高级根(4~5级根)皮层组织消失,次生生长特征明显,是典型的运输根[14,16]。由此可见,传统的细根(直径≤2.0 mm)系统至少由2部分组成,根系统末端非木质化的吸收根(1~3级根)和木质化较高的非吸收根(4级根以上)组成。林分密度的直接作用是影响树木个体的生长空间和资源获取,与非吸收根相比,吸收根对林分密度的响应可能更加敏感。然而,在区分根系功能基础上,探讨人工林林分密度对树木根系作用规律的研究仍然缺乏。为此,本研究以株行距分别为 1.0 m×1.0 m、1.5 m×1.5 m、2.0 m×2.0 m 3种林分密度水曲柳人工纯林为研究对象,于2012年8月采用根钻法,研究了林分密度对单位面积总细根和吸收根生物量、根长密度及其垂直分布的影响,同时也计算了单株水平上的根系指标。依据前期水曲柳根系解剖学和形态学研究[14,17],将直径≤0.5 mm细根定义为吸收根。本研究的主要目的是:①揭示不同林分密度对水曲柳吸收根生物量和根长密度的影响;②比较不同林分密度下吸收根垂直分布的差异;③明确影响林分和单株水平上根系生长的主要因子。这些研究结果,将增进对林分密度与树木根系生长之间相互关系的认识和理解。
1 研究地自然概况
研究地点位于黑龙江省尚志市东北林业大学帽儿山实验林场(127°36'~127°37'E,45°17'~45°18'N),海拔350~450 m,坡度10°左右。该地区属温带大陆性季风气候,年平均气温2.8℃,年平均降水量723.8 mm,年平均蒸发量1094 mm,无霜期120~140 d。水曲柳密度试验林位于山坡中部,土壤为暗棕壤,土层厚度35~40 cm。水曲柳密度试验林于1998年在皆伐林地上营造,有3种不同的造林密度,株行距分别为 1.0 m×1.0 m(S1)、1.5 m×1.5 m(S2)、2.0 m×2.0 m(S3)。2012年8月初进行调查取样,此时各林分郁闭度均达到0.9。在每种密度试验林内分别设置3个重复固定标准地,面积为20 m×30 m,进行每木检尺、土壤性质调查及根系取样工作,林分概况见表1,各个处理林分的立地条件基本一致。
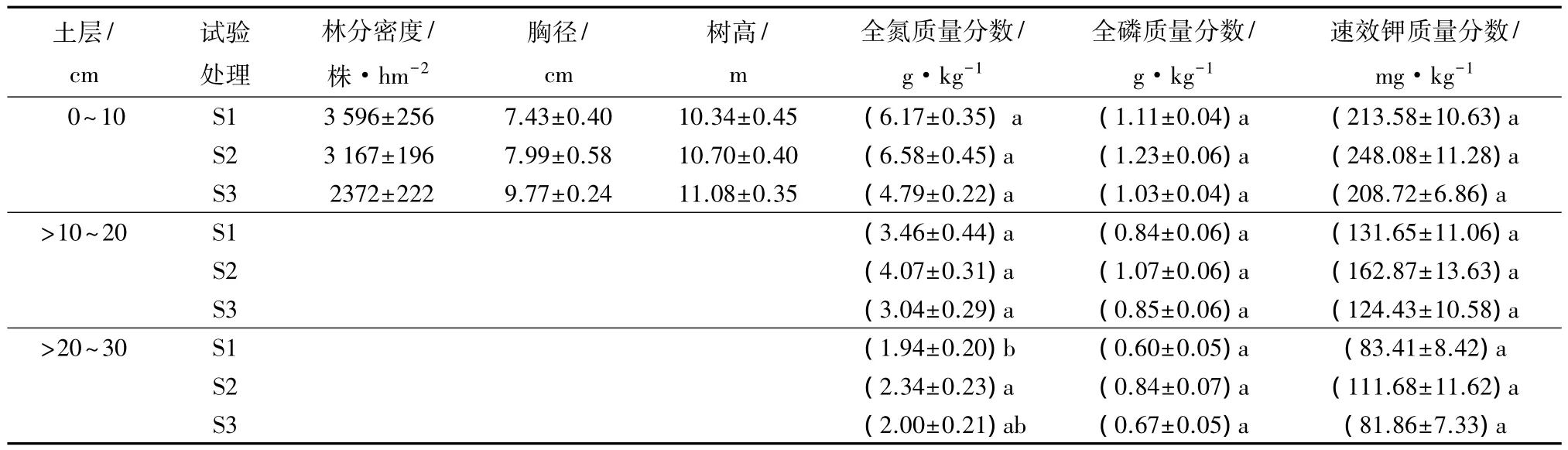
表1 不同密度水曲柳人工林林分与土壤特征
2 研究方法
2.1 样品采集与处理
在每块标准地内随机选择5个样点,使用根钻(内径为60 mm)采集包含着根系的土芯,取样深度设为30 cm,分为3层(0~10 cm为表层,>10~20 cm为亚表层,>20~30 cm为底层),合计取样数为135个。将样品装入已标号的封口袋内,4 h内带回实验站进行处理。用2 mm筛筛出各约100 g土芯部分鲜土,并手工将土壤中的根系个体挑出,风干后依据相应国家标准,进行理化性质分析。其余含有根系的土芯放入小桶内充分浸泡(约2 h),以便根系与土壤分离,在流水中过40目筛网,去除杂质及土壤颗粒,洗净后将根系低温冷冻保存。实验室内进行根系形态指标测定。根据分支结构和形态特征,区分水曲柳、其他木本植物和草本植物根系,进一步根据外形、颜色、弹性等区别死根和活根[18],将水曲柳活根按直径≤0.5、>0.5~1.0、>1.0~2.0 mm 分级。在每一块标准地随机选择3个样点的细根,按照取样的土层及划分的直径等级进行扫描(扫描仪:EPSON EXPRESSION 10000 XL,分辨率设为400 dpi),采用软件(WinRhizo 2004b,Regent instruments Inc,Canada)进行根系平均直径和总长度的测定。将所有细根样品置于65℃下烘干至恒质量并称质量(精确度=0.0001 g),结合形态测定结果计算比根长(1 g干质量根系的生物量),依据各土芯干质量和对应直径等级的比根长,推算每个土芯内的根长密度以及根生物量[19]。
2.2 数据处理
取样单元为样地,每个密度林分下重复样地数为 3。依据每一个直径等级(≤0.5、>0.5~1.0、>1.0~2.0 mm)和土层(0~10、>10~20、>20~30 cm),分别计算每个土壤深度上,单位面积对应的根系生物量和根长密度,整个土壤剖面(0~30 cm)则以相同直径等级根系指标加和计算。根据调查时的林分密度,结合单位面积生物量和根长,计算单株水平上不同直径等级根系生物量和根长密度。在单因素方差分析基础上,进行多重比较(LSD),比较不同密度上单位面积和单株水平上根系生物量和根长密度的差异;采用三因素方差分析方法,检验林分密度、土壤深度及直径等级对根系的影响,显著性水平设为P<0.05。所有数据统计分析均采用 SPSS 13.0(2010年,SPSS,USA)完成。
3 结果与分析
3.1 根系生物量和根长密度
林分水平上,随着密度从高到低,单位面积水曲柳总细根(直径≤2.0 mm)生物量(0~30 cm)呈小幅度增加,S1、S2、S3 分别为 279.4、285.9、287.3 g·m-2,而吸收根(直径≤0.5 mm,0~30 cm)生物量则是随密度的降低呈下降趋势,S1、S2、S3分别为168.2、167.2、160.8 g·m-2(表 2)。总细根与吸收根根长密度的变化趋势相同,处理S2具有最高的根长密度,其中吸收根比S1、S3分别高出16.9%和15.4%。相比较,直径>1.0~2.0 mm 根群生物量随密度减少而明显增加,而直径>0.5~1.0 mm 和>1.0~2.0 mm 根群的根长密度则均在S2中最高,而S1非吸收根(直径>0.5~1.0 mm 和>1.0~2.0 mm)生物量和根长与S3的相互关系并不恒定。吸收根占总细根生物量比例随密度降低而减少,S1、S2、S3分别减少60%、58%、56%,但是根长密度比例则几乎无变化,约96%。由于各密度处理内重复样地间根系生物量和根长变异较大,因此,不同处理间的差异并未达到显著性水平(表2)。单株水平上,各直径等级根群生物量和根长密度均随着密度的降低而增加(表2)。处理S3吸收根生物量和根长密度分别比S1、S2高出 50.4%、30.9%和 56.7%、17.9%。相比较,非吸收根中直径>1.0~2.0 mm根群生物量和根长在不同密度之间差异较大,处理S3生物量和根长分别比S1、S2 高出 88.6%、47.3%和 112.3%、30.0%(表 2)。

表2 不同密度水曲柳人工林单位面积及单株细根生物量和根长密度(0~30 cm土层)
3.2 根系的垂直分布特征
对于吸收根和直径>0.5~1.0 mm 根群,各密度下土壤表层(0~10 cm)均具有最高的生物量和根长密度,而直径>1.0~2.0 mm 根群则并不规律(表 3)。不同密度处理下吸收根的生物量和根长密度(除S2外)均随土壤深度加深规律性地减少。无论是吸收根还是非吸收根,其生物量和根长在不同土层中的相对比例均没有表现出随造林(林分)密度发生系统的改变。然而,底层(>20~30 cm)吸收根系占整个土壤剖面的相对比例,随着吸收根和总细根根长密度的增加而降低,表现为S2底层吸收根系生物量和根长相对比例分别为18.5%和22.4%,而根长密度稍低的S3分别为13.4%和11.8%,根长密度最低的 S1 则为 6.7%和 9.9%。直径>0.5~1.0 mm 根群垂直分布的比例也随根长密度发生变化,其规律与吸收根相同。但是,直径>1.0~2.0 mm 根群则无类似规律。

表3 不同密度水曲柳人工林单位面积细根生物量和根长密度的垂直分布
3.3 影响根系生长的因子
密度对根系生物量和根长的影响不显著(表4),这可能与同一处理重复样地间数值差异较大有关(表2)。根系直径等级和土壤深度(土层),以及二者的交互作用,对林分和单株水平上的根系生物量和根长密度均存在显著影响(表4)。
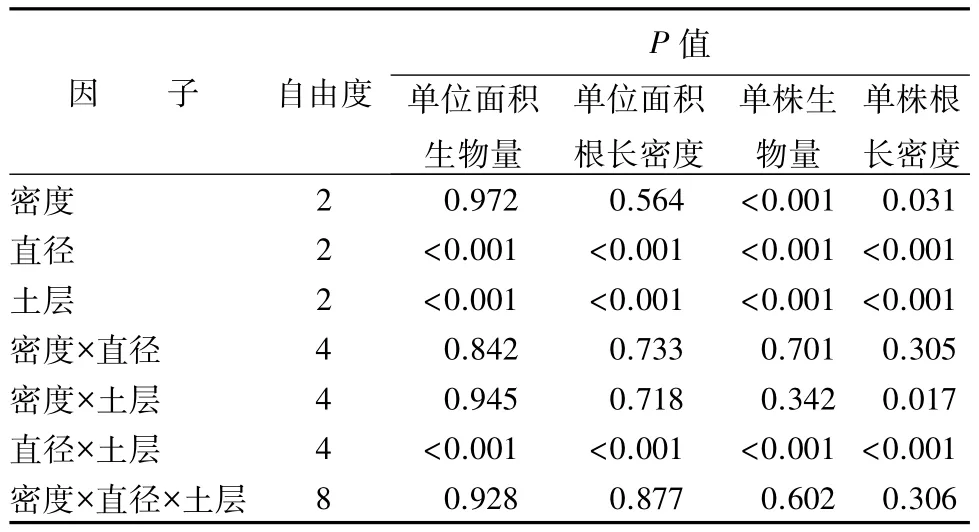
表4 林分密度、直径等级、土壤深度(土层)对单位面积和单株水平上根系生物量与根长密度的影响
4 讨论
4.1 密度对水曲柳根系生物量和根长密度的影响
本研究中,3种林分密度下水曲柳总细根(直径≤2.0 mm)生物量(0~30 cm 土层)为 279~283 g·m-2,低于或略低于以往相同树种的研究。例如,梅莉等[20]报道了本地区水曲柳人工林(17年生,株行距 1.5 m×2.0 m)细根生物量为 290.77 g·m-2;而朱胜英等[21]对50年生水曲柳天然次生林研究发现细根生物量(0~30 cm 土层)为487.8 g·m-2。这些研究说明,林分密度、林龄和林分类型均对细根生物量有重要影响,但是这些因子的相对贡献则需要进一步研究。根长密度是评价林分水平上根系对水分和养分吸收能力的一个重要指标。在本研究中,吸收根(直径≤0.5 mm)根长密度(14811~15017 m·m-2)均高于以往的研究报道。例如,Wang等[22]发现本地区水曲柳人工林前3级根(平均直径均小于0.5 mm)在9月份的根长密度(0~10 cm土层)为6400 m·m-2;梅莉等[20]的研究显示水曲柳年平均根长密度(直径≤1.0 mm;取样深度0~20 cm)为6102 m·m-2。本研究于8月初采样,是水曲柳根系现存量的秋初高峰期[23-24],同时考虑到根系取样深度和直径等级的差异,因此,本研究结果高于以往的结果也具有合理性。
尽管林分密度对细根生物量有重要影响,但是已有研究呈现出来的规律并不一致。随着造林(或者林分)密度增加,细根生物量表现出了单调增加、减少或者非线性的特征[5-7,9]。本研究结果显示,对于当前发育阶段的水曲柳人工林,单位面积总细根(直径≤2.0 mm)生物量随着初植密度增加呈降低趋势,这与李志辉等[8]和田宇明等[9]研究结果一致。然而,因为处理内根系生物量的较大变异,各密度处理间并未达到统计上的显著性。针对本密度实验林的前期研究发现,10年生水曲柳细根(直径≤2.0 mm)生物量为 80~145 g·m-2,也表现出随密度增加而降低的趋势[9]。该研究细根生物量远小于本研究的结果(279~283 g·m-2),一定程度上这可能与其采用了方形整段剖面法有关,而笔者采用的是根钻法。整段剖面法手捡根系会在很大程度上会丢失许多细小的根系,尤其那些起到重要资源获取功能的吸收根。因此,除了林分自身生长导致根系生物量增加以外(本研究中林分为14年生),根钻法能够较为完整的回收根系,也会使得本研究中根系生物量数据高于以往。根长密度与根系生物量不同,各直径等级根系均以中等密度的S2最高,这很大程度上是因为S2处理具有最大的比根长,其0~30 cm土层吸收根比根长为89~101 m·g-1,而S1为77~97 m·g-1,S3 为 71~94 m·g-1。因此,尽管S2生物量并非最高,其根长密度却最大。虽然有研究指出造林密度和间伐会影响不同根序的比根长和直径[10,25],然而,为何中等密度下水曲柳根系具有最大的比根长,还需要进一步的研究。
需要注意的是,单位面积水曲柳吸收根的生物量随密度降低而减少,而非吸收根则随着密度降低而增加。这说明密度较低情况下,土壤资源相对丰富,树木不需要在单位面积(或体积)上投入更多生物量的根系来进行资源的争夺,而将较多的生物量投入那些直径较粗的木质化、起运输和储存作用的非吸收根系中。因此,通过将细根系统按照其执行的主要生理功能进行划分,研究根系生物量与林分密度的联系,获得了以往研究中未发现的信息(即总细根随密度增加而减少,吸收根则增加),这些研究结果,对于了解密度的作用规律具有重要的意义。
此外,单株水平上各等级根系的生物量和根长密度均随密度降低而增大,这与一些树种,如尾巨桉[8]、杉木[11]、马尾松[25]和红松[26]的报道一致。显然,密度降低后,树木个体获取营养空间增大,促进了地上和地下各组分的生长。不过,研究显示,直径、土层及二者交互作用均对根系生物量和根长密度有显著影响。因此,未来的人工林密度与根系生长的研究中,需要考虑这些因子的重要作用。
4.2 密度对根系空间分布特征的影响
水曲柳吸收根生物量主要集中在0~10 cm土层,约占整个采样土壤剖面的45%,且在不同密度处理下差异较小。这主要是因为表层土壤的温度和湿度较高,土壤微生物活动旺盛,养分有效性较高,有助于吸收根对土壤资源的高效获取和利用[21,27]。本研究中,各直径等级根系生物量和根长在不同土层中的相对比例均未随林分密度发生有规律的变化,这与一些研究者报道不同[9,11,24,28]。但是,研究发现>20~30 cm土层吸收根占整个土壤剖面的相对比例,随着吸收根(或总细根)根长密度的增加而降低。这表明,很可能不是林分密度直接影响了根系特别是吸收根的垂直分布,而是林分水平上根长密度的直接作用。因为不同树木个体的根系对土壤资源的竞争,是直接通过根长密度来体现出来的。以往研究中根系垂直分布特征随造林(林分)密度的系统变化,很可能在于根长密度在不同密度间也发生了有规律的变化。进一步分析以往的报道[9,24,28],发现在根长密度高的林分中,底层细根生物量也较高。本研究中吸收根在土壤下层分布比例的增加,很大程度上在于避免根系在表层的激烈竞争。不过需要指出的是,试图回答究竟是林分密度还是根长密度直接导致了根系垂直分布的改变,则有待于对多个树种和多次取样来进一步验证。
5 结论
通过对水曲柳3种林分密度研究表明,单位面积总细根(直径≤2.0 mm)生物量随密度降低而增加,而吸收根(直径≤0.5 mm)生物量则是随密度的降低呈下降趋势,但是二者在不同密度处理间的差异均未达到统计上显著水平。中等林分密度下总细根和吸收根均表现出最高的根长密度。单株水平上,各直径等级根群生物量和根长密度均随着密度的降低而增加。直径等级和土壤深度均是影响水曲柳根系生长的重要因子。此外,吸收根根长密度的变化,是导致吸收根在底层分配比例增加的一个重要因素。通过将传统的细根依据其功能划分成吸收根和非吸收根组分,能够获得密度对林木根系作用规律的准确信息,从而增进对二者相互作用机制的理解。
[1]Eissenstat D M,Wells C E,Yanai R D,et al.Building roots in a changing environment:implications for root longevity[J].New Phytologist,2000,147(1):33-42.
[2]王政权,郭大立.根系生态学[J].植物生态学报,2008,32(6):1213-1216.
[3]沈国舫.森林培育学[M].北京:中国林业出版社,2001:64-75.
[4]姚东和,杨民胜,李志辉.林分密度对巨尾桉生物产量及生产力的影响[J].中南林学院学报,2000,20(3):20-23.
[5]李志辉,陈少雄,黄丽群,等.林分密度对邓恩桉生物产量及生产力的影响[J].中南林业科技大学学报,2007,27(5):1-5.
[6]徐程扬,张华,贾忠奎,等.林分密度和立地类型对北京山区侧柏人工林根系的影响[J].北京林业大学学报,2007,29(4):95-99.
[7]Kerr G.Effects of spacing on the early growth of planted Fraxinus excelsior L.[J].Canadian Journal of Forest Research,2003,33(7):1196-1207.
[8]李志辉,陈少雄,谢耀坚,等.林分密度对尾巨桉生物量及生产力的影响[J].中南林业科技大学学报,2008,28(4):49-54.
[9]田宇明,王庆成.初植密度对10年生水曲柳人工林生物量及根系的影响[J].林业科学,2011,47(7):102-107.
[10]周伟,陈信力,李瑞霞,等.林分密度对侧柏人工林细根形态的影响[J].林业科技开发,2012,26(4):35-38.
[11]吕士行,余雪标.杉木造林密度与根系生长关系研究[J].林业科技通讯,1990(11):1-3.
[12]Vanninen P,Mäkelä A.Fine root biomass of Scots pine stands differing in age and soil fertility in southern Finland[J].Tree Physiology,1999,19(12):823-830.
[13]Wirth C,Schulze E D,Kusznetova V,et al.Comparing the influence ofsite quality, stand age, fire and climate on aboveground tree production in Siberian Scots pine forests[J].Tree Physiology,2002,22(8):537-552.
[14]Guo Dali,Xia Mengxue,Wei Xing,et al.Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species[J].New Phytologist,2008,180(3):673-683.
[15]Pregitzer K S,Deforest J L,Burton A J,et al.Fine root architecture of nine North American trees[J].Ecological Monographs,2002,72(2):293-309.
[16]许旸,谷加存,董雪云,等.海南岛4个热带阔叶树种前5级细根的形态、解剖结和组织碳氮含量[J].植物生态学报,2011,35(9):955-964.
[17]王向荣,王政权,韩有志,等.水曲柳和落叶松不同根序之间细根直径的变异研究[J].植物生态学报,2005,29(6):871-877.
[18]刘聪,项文化,田大伦,等.中亚热带森林植物多样性增加导致细根生物量“超产”[J].植物生态学报,2011,35(5):539-550.
[19]刘佳,项文化,徐晓,等.湖南会同5个亚热带树种的细根构型及功能特征分析[J].植物生态学报,2010,34(8):938-945.
[20]梅莉,王政权,韩有志,等.水曲柳根系生物量、比根长和根长密度的分布格局[J].应用生态学报,2006,17(1):1-4.
[21]朱胜英,周彪,毛子军,等.帽儿山林区6种林分细根生物量的时空动态[J].林业科学,2006,42(6):13-19.
[22]Wang Zhengquan,Guo Dali,Wang Xiangrong,et al.Fine root architecture,morphology,and biomass of different branch orders of two Chinese temperate tree species[J].Plant and Soil,2006,288(1/2):155-171.
[23]史建伟,王政权,于水强,等.落叶松和水曲柳人工林细根生长、死亡和周转[J].植物生态学报,2007,31(2):333-342.
[24]梅莉,韩有志,于水强,等.水曲柳人工林细根季节动态及其影响因素[J].林业科学,2006,42(9):7-12.
[25]丁贵杰.马尾松人工林生物量和生产力研究Ⅰ:不同造林密度生物量及密度效应[J].福建林学院学报,2003,23(1):34-38.
[26]陈硕芃,王韶仲,王政权,等.密度结构对大青川红松人工林细根生物量与根长密度的影响[J].森林工程,2013,29(4):1-7.
[27]王祖华,李瑞霞,关庆伟.间伐对杉木不同根序细根形态、生物量和氮含量的影响[J].应用生态学报,2013,24(6):1487-1493.
[28]刘运科,范川,李贤伟,等.间伐对川西亚高山粗枝云杉人工林细根生物量及碳储量的影响[J].植物生态学报,2012,36(7):645-654.
