福建地区杉木枝条基径最优模型1)
2014-09-18高志雄王新杰李海萍
高志雄 王新杰 李海萍 张 鹏
(省部共建森林培育与保护教育部重点实验室(北京林业大学),北京,100083)
枝条是树叶着生的场所,枝条的大小、结构直接影响着树冠。而树冠的形状、大小和结构决定了树木的生长活力和生产力[1-2]。人们通过对树冠枝条的分析,可以了解一株树木的基本轮廓。通过对不同条件下枝条变化的研究,能了解树木对不同生态条件适应与变异的形式。树冠枝条在树干上的分布方式也在一定程度上决定了树冠的冠形[3]。枝条是组成树冠的骨架,决定了树冠的形状及其大小。由于枝条生长在树干上,基径较大的枝条会产生较大的节子,也直接影响木材材质。枝条直径的大小直接决定了节子在树干上的大小,而节子的大小是评价木材品质的重要指标之一[4]。枝条的长短决定了树冠的大小形态和树木材质[5-6]。在国外,枝条基径的大小已经作为评价森林经营效益的一个重要指标[7-8]。由此可见,枝条基径在森林经营中是非常重要的。对于活立木而言,我们几乎不能直接获取枝条基径数据。在研究枝条基径模型时,很多学者以林木变量(胸径、树高、冠长等)和枝条的着枝深度(枝条着生的位置距离树梢的长度)等作为自变量,建立不同树种的枝条基径预测模型。提出的模型大致分为3类:第一类模型是经验模型,这类模型主要采用线性回归技术[9],模拟枝条基径与林木因子之间的线性关系,该类模型的精度较高,由于变量因子之间可能存在自相关,需要考虑共线性问题,而且模型的林学解释性较差[10-13];第二类模型是理论模型,这类模型的结构多以非线性模型为主,如Weibull模型、Richards方程、单分子 Mitschelich方程、异速生长模型等[14-15],模型中变量因子较少,模型参数具有林学或生态学方面的解释,在林业中应用较广泛;第三类是复合模型,也称半经验模型,这类模型以理论模型为基础模型,在理论模型参数中引入与因变量有显著相关的变量因子,构建的模型参数具有较好的林学或生物学意义,而且模型拟合精度较高。
以往的研究,对单分子理论模型的没有做出详细的比较,本文通过对福建将乐地区杉木枝条基径数据对常用的单分子理论模型进行详细的比较,并在此基础上对最优的单分子理论模型添加变量得到复合模型。经过复合模型与线性模型的拟合优度、残差分析和预测精度等比较得出最优模型。
1 研究区概况
福建省将乐国有林场位于福建省西北部,地处武夷山脉东南部、金溪河畔,其地理坐标为东经117°05'~117°40',北纬 26°25'~27°04',多为以中、低山地貌以低山丘陵为主,海拔400~800 m;年平均气温18.7℃,无霜期273 d,生长期长,降水量1672.3 mm;属亚热带季风气候区,具有海洋性和大陆性气候特点。林场总面积7173.4 hm2,有林地面积6131.7 hm2,森林覆盖率 93.5%。
2 研究方法
2.1 实验数据的采集
结合立木径级与年龄分布的规律,按典型取样的原则,在研究区内设置杉木人工林标准地,对标准地的杉木进行每木检尺,在标准地外选取平均木一株作为标准木,样木的选取要避开林缘,选择没有干折或分杈的正常树木。共获得解析标准木36株,逐株进行树干解析、枝解析及生物量的测定。
将野外调查数据按解析样木、枝解析进行归类、统计(见表1)。对伐倒的解析木主要记录:胸径(DBH)、年龄(A)、树高(HT)、活枝高(HCB)、死枝高、冠幅(CW)、冠长(CL)等林木因子;枝解析主要记录:枝条基径(BD)、着枝深度(DINC)、平均枝条基径(BDm)、活枝高(HCB)、高径比(HD)、长度,并且记录活枝最宽冠幅。

表1 数据整体情况
按照检验样本占总样本数20%的原则,在解析木中,取出7株不同生长状态的样木作为检验数据,其余的29株作为拟合数据(见表2)。共测量了2315个活枝条,其中,作为拟合数据的枝条有1863个,作为检验数据的枝条数据有452个。

表2 数据龄组分布 株
2.2 模型及检验
研究采用树冠主轴分层分割法[16],对树冠进行分层。本文中以0.5 m为一段进行分层,首先用粉笔将其划分成0.5 m一段,而后按段将每段的枝条进行测量。包括枝条的基径、长度、着枝角度、质量等。对每段的总枝条进行称质量,由每段枝条的数据,分别计算活枝、死枝每段枝条的基径、枝长平均值。根据每段枝长、基径平均值,在活枝、死枝中,分别选取2至3根标准枝。对标准枝称质量,将标准枝叶子摘掉,分别称取枝和叶的质量。
2.2.1 线性模型
对数据进行逐步回归,发现用枝条平均基径对数(ln(BDm))作为因变量,模型的拟合效果明显较优。当采用树高、胸径作为自变量进行模型拟合,结果存在严重的自相关现象。因此,对BDm有显著相关性的变量因子做一些适当的变换,得到了DINC取自然对数(ln(DINC))、HT与 DINC的乘积(HT×DINC)、树高比胸径(HD)、着枝深度的平方(DINC2)、以及胸径和着枝深度的乘积(DBH×DINC)。
取以上的变量因子为自变量,枝条平均基径取对数ln(BDm)为因变量,利用SPSS进行逐步回归,经过对数据的分析,以枝条基径取对数逐步回归,利用SPSS逐步回归的方法,进行逐步回归分析,选取最优线性模型。
线性模型的一般形式如下:
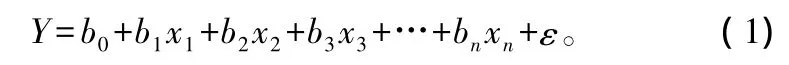
式中:Y 为因变量 ln(BDm),x 为自变量(DBH、CL、HT、HD、DINC等),bi为模型的参数和截距,ε为模型的随机变量。
2.2.2 复合模型
通过分析可以看出,枝条基径主要与枝条着枝深度有关,而且枝条基径随着枝条着枝深度的增加有明显的增加趋势,而且在枝条基部达到一个渐进值。可以认为枝条基径是关于枝条着枝深度的函数。目前应用较多的林分生长模型有Weibull模型、Richards方程、单分子 Mitschelich方程、Logistic方程、考尔夫方程、坎派兹方程等[17]。通过以着枝深度为自变量,ln(BDm)为因变量,对各个模型进行拟合。根据以往研究,理论模型主要有:改进的单分子Mitschelich模型、Logistic模型、Richards模型、修正的Weibul Korf模型和Gomperz模型:

非线性模型的建立,通过对常用的单分子理论模型进行筛选,得出最合适的单分子模型。选定好理论模型以后,对理论模型引入其他与因变量相关的变量,以增加模型的拟合优度。
2.3 模型的独立性检验与最优模型的选取
模型的独立性检验是通过拟合的模型来对检验样木的枝条基径进行预估。主要包括绘制的各自变量及模型估计值与模型残差值的散点图以及计算模型用于预估的效果和精度的各个统计量,主要包括平均偏差(ME)、平均绝对偏差(MAE)、平均相对偏差(MPE)、平均相对偏差绝对值(MAPE)以及精度(P)[18]。


式中:t0.05为置信水平0.05时的 T分布值;n为样本数;T为回归模型中参数的个数;yi为实测值;为模型预估值为平均预估值,即,n 为样本数。ME、MAE、MRE、MARE越小,P 越大的模型其预测能力越强,预测精度越高。
3 结果与分析
根据对数据的分析,得到基径与各因子的相关关系(见表3)。由表3可知,枝条基径与树木变量的相关性尤为明显,与着枝深度的相关性最大。为了进一步反映不同林分条件下枝条基径的变化规律,又以枝条基径与林分密度、断面积、平均胸径、优势木高等林分因子做了相关性分析,结果显示BD与这些林分因子相关性不显著。因此,所筛选出的相关性显著的变量为:DBH、HT、CL、DINC、CW及其组合变量等。以相关性显著的变量,进行逐步回归或非线性回归。

表3 各变量的相关性
3.1 线性模型
通过以枝条平均基径取对数(ln(BDm))为因变量,添加相关变量进行逐步回归,在得到的方程中,选取最佳的方程为:ln(BDm)=-0.386+0.056×DBH-0.033×CL-0.054×CW+0.212×ln(DINC)。
3.2 复合模型
对理论模型进行拟合结果如表4。
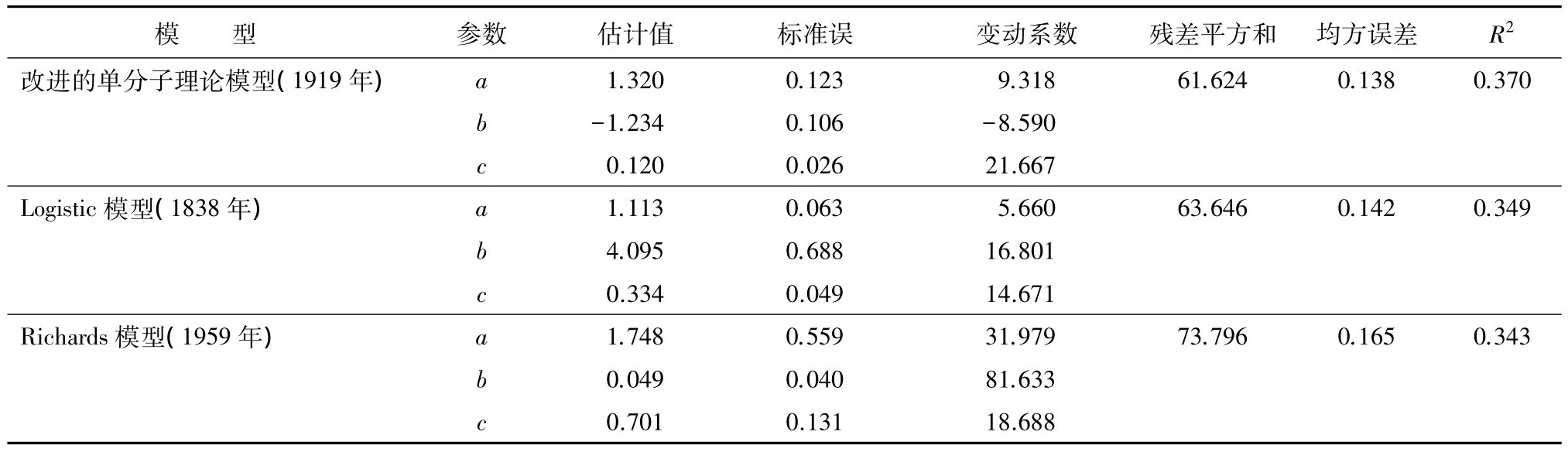
表4 理论模型拟合效果对比
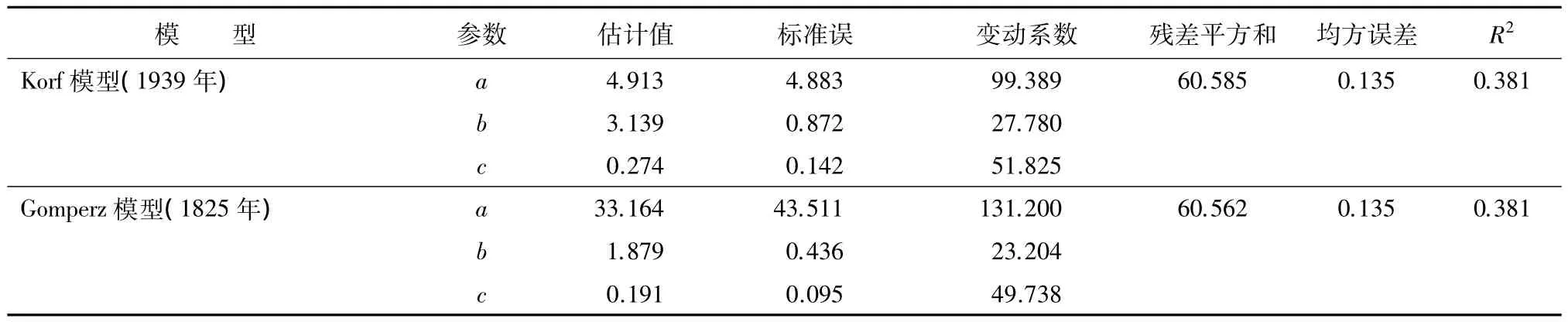
续(表4)
通过对几个常用的理论模型参数的标准误、残差平方和、均方误差和R2进行比较。通过比较发现korf模型、Gomperz模型的拟合效果略较好,其次是修改的weibull模型、logistic模型、richard模型。而对参数的标准误进行比较,发现单分子Mitschelich方程的参数标准误、参数变动系数均要小于korf模型和Gomperz模型。结合以上分析,最终选出改进的单分子Mitschelich方程,该方程的拟合效果最佳。
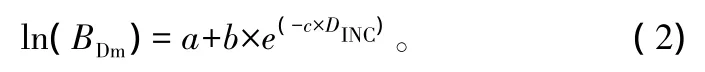
式中:a、b、c为模型参数,DINC为着枝深度。
理论模型只考虑着枝深度单个因子,对理论模型(2)的参数中引入其他与枝条基径相关的变量以及变量之间的组合,得到的方程为:ln(BDm)=(0.151×-0.031 ×CL)×(l-e-0.825×DINC)-0.0465。式中:ln(BDm)为枝条平均基径取对数,DBH为胸径,CL为冠长。
3.3 最优模型的选择
最优模型选择遵循的一般原则:模型精度比较高、测定因子容易获得、模型的结构比较简单及模型预估精度稳定性较高。通过计算各统计量指标(剩余残差平方和、剩余均方差、剩余标准差、决定系数),对各备选模型进行比较,选择剩余残差平方和最小、剩余标准差Sy,x最小、剩余均方差最小、相关指数(R2)(或调整的相关指数R2a)最大的模型作为最佳模型。结合各备选模型参数检验结果,进一步比较各模型拟合统计量。对以上三个模型进行对比(见表5)。
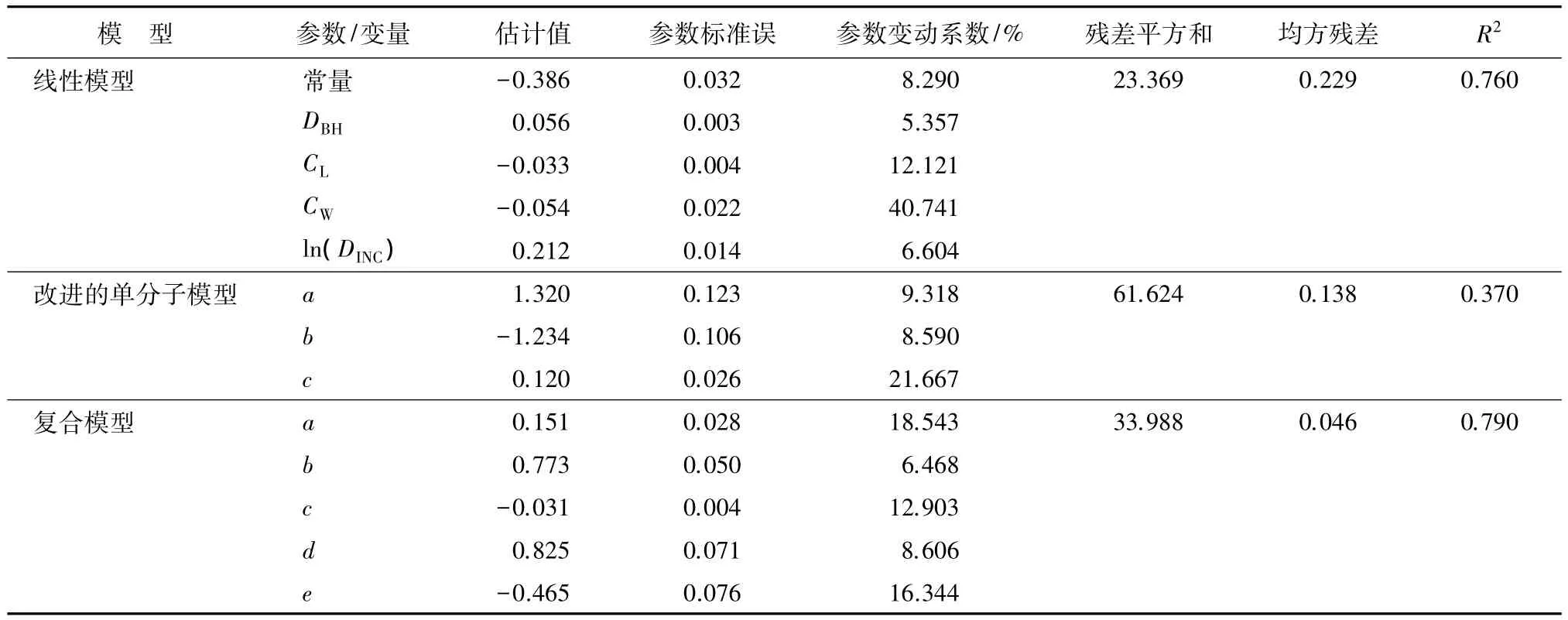
表5 模型拟合效果对比
由表5可见,复合模型的拟合优度最高,R2为0.790,均方残差为 0.046,复合模型的参数变动系数均小于20%,且平均参数变动系数为12.6%。说明模型变量对枝条平均基径的预测均有显著的影响,模型也比较稳定。线性模型的拟合精度较高,R2为0.760,均方残差 0.229,平均参数变动系数达 14.6%,参数最大变动系数为40.74%。改进的单分子理论拟合精度较低,平均参数变动系数为13.19%,由于该模型中只有一个变量,不足以表达枝条平均基径的变化。
残差分析(见图1)。残差点在残差值为0的线上下分布均匀,基本都落在±1的标准残差之内,这就表明方程的拟合效果良好。也有个别残差较大的样本,经过对比发现这些差异大的点大多都是在树冠的下层,由于树冠下层遮阴、林木之间的相互竞争以及人为因素的破坏所造成的。复合模型和线性模型的残差值分布更为均匀,且残差分布图较为相似;由于单分子模型只有一个变量,残差值分布随着枝条基径有上升的趋势。
通过以上分析,和残差分布图的比较,复合模型和线性模型的拟合效果较好。因此,对这两个模型进行检验(见表6)。

图1 不同枝条平均基径模型的残差分布

表6 模型检验
由表6可知,复合模型的检验精度为91.33%,高于线性模型的88.41%。平均偏差、平均偏差绝对值、平均相对偏差绝对值、复合模型的稳定性均要优于线性模型。综合各方面考虑复合模型为预估杉木枝条平均基径的最优模型。
4 结论与讨论
通过对常用的单分子模型筛选,得出最适合枝条平均基径的理论模型为改进的Mitscherlich单分子模型,说明该理论模型的结构最适合枝条基径与着枝深度的相关性,这与其他相关研究一致。
通过对多元逐步线性回归模型、单分子理论模型和复合模型的比较,基于单分子理论模型的复合模型优于多元线性逐步回归的经验模型,经验模型优于单分子理论模型。相比于线性模型而言,理论方程逻辑性强、适应性广等特点。并且理论方程是按照生物学理论或假设而推导出来的,有明确的生物学意义。由于枝条的采集数据较为复杂繁琐,以及人为或动物活动因素的干扰,导致枝条平均基径预估模型的拟合优度不太高,R2为 0.34~0.79。不同模型拟合精度,与模型变量的选择和模型的结构都有着密切的联系。
通过相关性分析,枝条基径与林木的胸径、冠长和枝条的着枝深度有关。最优模型的表达式可知,胸径一定时,着枝深度越大枝条基径越大;同一着枝深度,不同胸径的枝条的基径随树木的胸径的增大而增大;冠长与树高、活枝高密切相关,冠长越长说明树冠最底层的枝条保存的较好,而树冠底层的枝条着枝深度大,枝条的基径就会更大。本文中所选取模型的参数和变量较少,经过检验模型的预测精度为91.33%,精度较高。
通过杉木枝条基径最优预测模型的研究,可以在不伐倒树木的情况下对枝条的生长状况有一个了解,对枝条的基径做出预估。枝条基径直接影响木材节子的大小,是评价木材材质的一个指标。枝条基径与枝条的生物量显著相关,对于研究枝条生物量具有重要的意义。
[1]Marshall D D,Johnson G P,Hann D W.Crown profile equations for stand-grown western hemlock trees in northwestern Oregon[J].Canadian Journal of Forest Research-Revue Canadienne de Recherche Forestiere,2003,33(11):2059-2066.
[2]Chmura D J,Rahman M S,Tjoelker M G.Crown structure and biomass allocation patterns modulate aboveground productivity in young loblolly pine and slash pine[J].Forest Ecology and Management,2007,243(2/3):219-230.
[3]Gilmore D W,Seymour R S.Crown architecture of Abies balsamea from four canopy positions[J].Tree Physiology,1997,17(2):71-80.
[4]Gort J,Zubizarreta G A,Peltola H,et al.Differences in branch characteristics of Scots pine(Pinus sylvestris L.)genetic entries grown at different spacing[J].Annals of Forest Science,2010,67(7):705-705.
[5]Doruska P F,Burkhart H E.Modeling the diameter and locational distribution of branches within the crowns of loblolly pine trees in unthinned plantations[J].Canadian Journal of Forest Research,1994,24(12):2362-2376.
[6]Maguire D A,Moeur M,Bennett W S.Models for describing basal diameter and vertical distribution of primary branches in young Douglas-fir[J].Forest Ecology and Management,1994,63(1):23-55.
[7]Maguire D A,Johnston S R,Cahill J.Predicting branch diameters on second-growth Douglas-fir from tree-level descriptors[J].Canadian Journal of Forest Research,1999,29(12):1829-1840.
[8]Mäkinen H,Song T.Evaluation of models for branch characteristics of Scots pine in Finland[J].Forest Ecology and Management,2002,158(1):25-39.
[9]刘兆刚,舒扬,李凤日.樟子松人工林一级枝条基径和枝长模型的研究[J].植物研究,2008,28(2):244-248.
[10]Hein S,Mäkinen H,Yue C,et al.Modelling branch characteristics of Norway spruce from wide spacings in Germany[J].Forest Ecology and Management,2007,242(2/3):155-164.
[11]姜立春,李凤日,张锐.基于线性混合模型的落叶松枝条基径模型[J].林业科学研究,2012,25(4):464-469.
[12]卢军.帽儿山天然次生林树冠结构和空间优化经营[D].哈尔滨:东北林业大学,2008.
[13]刘星丽,安晨霞,吴继东,等.樟子松人工林一级枝条基径模型的研究[J].林业科技情报,2009,41(1):24-26.
[14]李凤日.长白落叶松人工林树冠形状的模拟[J].林业科学,2004,40(5):16-24.
[15]刘艳艳.樟子松人工林树冠结构的研究[D].哈尔滨:东北林业大学,2005.
[16]Chiba Y.Plant form based on the pipe model theory II.Quantitative analysis of ramification in morphology[J].Ecological Research,1991,6(1):21-28.
[17]Soares P,Tome M.A tree crown ratio prediction equation for eucalypt plantations[J].Annals of Forest Science,2001,58(2):193-202.
[18]郭孝玉.长白落叶松人工林树冠结构及生长模型研究[D].北京:北京林业大学,2013.
