天目山国家级自然保护区毛竹林扩张对生物多样性的影响1)
2014-09-18林倩倩马元丹吴呈昱
林倩倩 王 彬 马元丹 吴呈昱
(浙江农林大学亚热带森林培育国家重点实验室培育基地,临安,311300)
赵明水
(浙江天目山国家级自然保护区管理局)
生物多样性是地球的生命基础,保护生物多样性已成为全球性的问题。自然保护区在保护生物多样性中发挥着十分重要的作用。建立自然保护区不仅能保护生态系统多样性、物种多样性,而且能保护遗传多样性,是保护生物多样性的最有效途径。但目前在亚热带地区,自然保护区内的毛竹扩张现象比较普遍和严重,正威胁着保护区核心区的重点保护物种,影响了自然保护区的生物多样性及整个森林生态系统的生态安全的维护。
毛竹(Phyllostachys pubescens)原产我国亚热带地区,禾本科竹亚科刚竹属,为单轴型散生竹类。毛竹为多年生木本植物,通过地下竹鞭进行无性繁殖,一个林分即为若干基株克隆的无性系种群[1]。毛竹地下鞭根系统发达,其生长、繁殖和穿透能力极强。毛竹靠地下扩鞭可渗透到其他森林类型,然后迅速长成幼竹,在其他森林类型中形成毛竹的分布前沿。随着时间的推移,其分布面积不受人为控制地增加[2]。毛竹在保护区内的扩张不仅对植物群落生物多样性造成显著影响,亦会对林地表层土壤基本理化性质[3]、土壤水稳性团聚体及化学性质产生影响[4];对本地鸟类多样性造成较大负面效应[5];使大部分森林类型树冠对酸雨的缓冲作用大大减少[6];森林植被生物量、森林土壤碳含量和生态系统总碳储量均大大下降[7];使林下土壤逐渐贫瘠,生境质量下降。
由于毛竹强大的扩鞭能力,近年来处于亚热带的多个自然保护区如天目山自然保护区[8-10]、鸡公山自然保护区[11]、武夷山自然保护区[12]、大岗山自然保护区[13]、井冈山自然保护区[14]等自然保护区的毛竹林不断扩张,逐年蚕食常绿落叶阔叶混交林、针阔混交林等天然植被,造成了保护内毛竹林纯林化,从而影响保护区丰富的生物多样性。据1956年天目山森林资源调查记载,当时毛竹林面积为55.1 hm2,2004 年调查的毛竹林面积为 87.5 hm2,48 年间增长58.7%,近年来扩张速度明显加快。丁丽霞等[8]用遥感和地理信息系统技术,监测该自然保护区在1985、1991、2003年3个时期的毛竹林面积及其变化。结果显示,区内毛竹林面积以平均4.47 hm2·a-1的增长速度扩张,并有继续快速蔓延的趋势,严重侵占了周围原始植被(特别是阔叶林分)。因此,对天目山国家级自然保护区毛竹林扩张对生物多样性的影响进行调查和分析迫在眉睫。
1 研究区概况
天目山国家级自然保护区位于30°18'30″~30°21'37″N,119°24'11″~119°27'11″E。现有面积为4284 hm2,其中国有林部分1018 hm2,集体林部分3266 hm2。年平均气温为12.4℃,≥10℃积温2500~5100℃,无霜期 209~235 d,相对湿度76%~81%,年降水量1390~1870 mm,年太阳总辐射3770~4460 MJ·m-2。土壤为红壤、黄壤、黄红壤或棕黄壤,pH 值在 4.7~6.0,凋落物厚度小于 5 cm,有机质质量分数2.37%~10.74%,全氮质量分数0.1%~0.5%,总磷质量分数 0.07%~0.15%,总钾质量分数0.31%~1.27%。这些自然条件适宜毛竹的温湿习性和其他生理学、生物学及生态学特性,有利于毛竹生长。
2 研究方法
2.1 样地设置及调查方法
在研究区内,分别在毛竹林、常绿阔叶林、毛竹-常绿阔叶林、针叶林、毛竹-针叶林、针阔混交林、毛竹-针阔混交林等7个林型中设置样地。每个群落中对乔木层、灌木层、草本层进行分层调查,记载每块样地地理位置、地形、地貌、海拔高度、土壤状况、植物种类组成等。
每个林型设一个20 m×20 m的样方,调查乔木植被组成,并在其中分别设置3个2 m×2 m的灌木样方,3个1 m×1 m的草本样方。乔木样方内每木检尺(胸径大于3 cm的所有植株),即测量乔木层胸径(DBH)和树高(H);灌木层测量高度大于0.3 m的灌木基径、盖度和高度;草本层测量物种的盖度、多度和高度指标。
2.2 数据分析
根据样地调查数据,分别计算乔木层、灌木层及草本层的重要值。计算公式为:
乔木层重要值:IV=(相对密度+相对优势度+相对高度)/3;
灌木层和草本层的重要值:IV=(相对密度+相对高度+相对盖度)/3;
按马克平等[15-16]所评述的植物群落多样性测度方法,本文采用Patrick丰富度指数(即物种数R)、Simpson多样性指数(D)、Shannon-Wienner多样性指数(H)和Pielou均匀度(E)指标,进行群落多样性分析。样地中每个样方分别计算乔木层、灌木层、草本层的各多样性指标,揭示毛竹林扩张过程中植物群落多样性的水平空间分布格局。

式中:S为样地中物种数目;N为样地中多度指标总和(分层或总体);Ni为第i个种多度指标;Pi=Ni/N,i=1、2、3、4、…、n。
3 结果与分析
3.1 毛竹立竹度及其他树种立木度变化
毛竹的立竹度是指在某一区块内毛竹的株数,是毛竹地上部分生物量的重要组成因素,是竹林林分结构变化的具体反映[17]。根据调查,毛竹在扩张的过程中,其立竹度随着群落类型的改变而改变(见表1)。在7个群落类型中,除毛竹林立竹度最大以外,立竹度由大到小的顺序为:毛竹-针阔混交林、毛竹-常绿阔叶林、毛竹-针叶林。另外三个群落针叶林、针阔混交林和常绿阔叶林因没有毛竹的存在,毛竹立竹度为0。
7种群落类型中,总立木数由大到小的顺序为:针叶林,毛竹-针叶林、针阔混交林,毛竹-针阔混交林、常绿阔叶林,毛竹-常绿阔叶林。可见毛竹的入侵分别降低了相应的群落类型的总立木数。
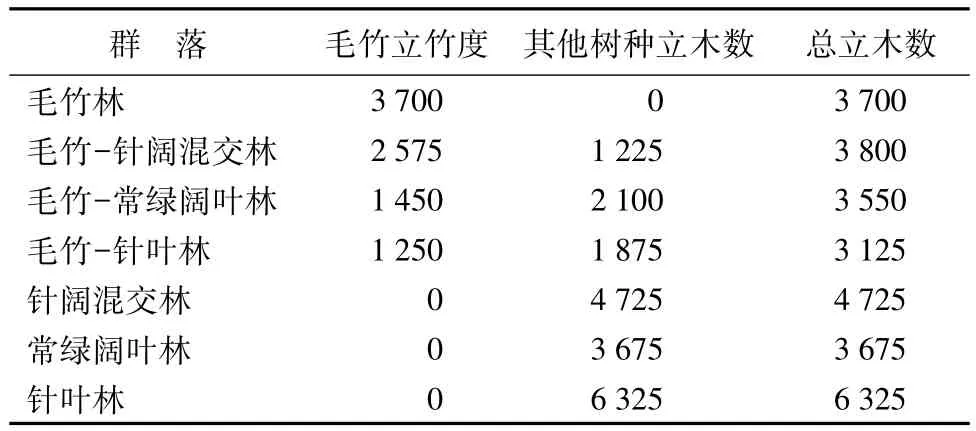
表1 毛竹扩张过程中立竹度及其他树种立木数 株·hm-2
3.2 物种组成和群落特征
经统计,在天目山自然保护区的7个样地中,共发现植物有:樟科、壳斗科、蔷薇科、山茶科、豆科、鳞毛蕨科、漆树科等61科104属142种。数量最多的为樟科共有6属14种,其次是壳斗科4属11种、蔷薇科8属11种、山茶科2属10种、豆科6属6种,这5科占到了总种数的32.64%,占总属数的25.01%(见表2)。
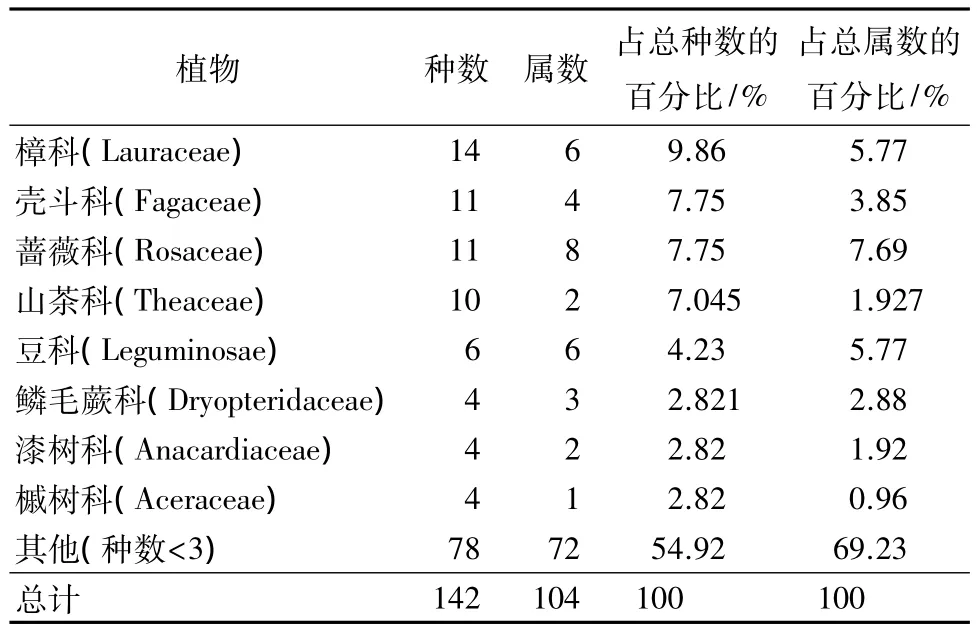
表2 不同群落中植物主要物种科属组成
3.3 不同森林类型物种组成及其数量特征
以样地内的调查数据为基础,统计天目山国家级自然保护区7种森林类型物种组成的数量特征(限于篇幅,仅列出前5种物种的重要值)。通过各群落乔木层物种重要值的排序(见表3)。
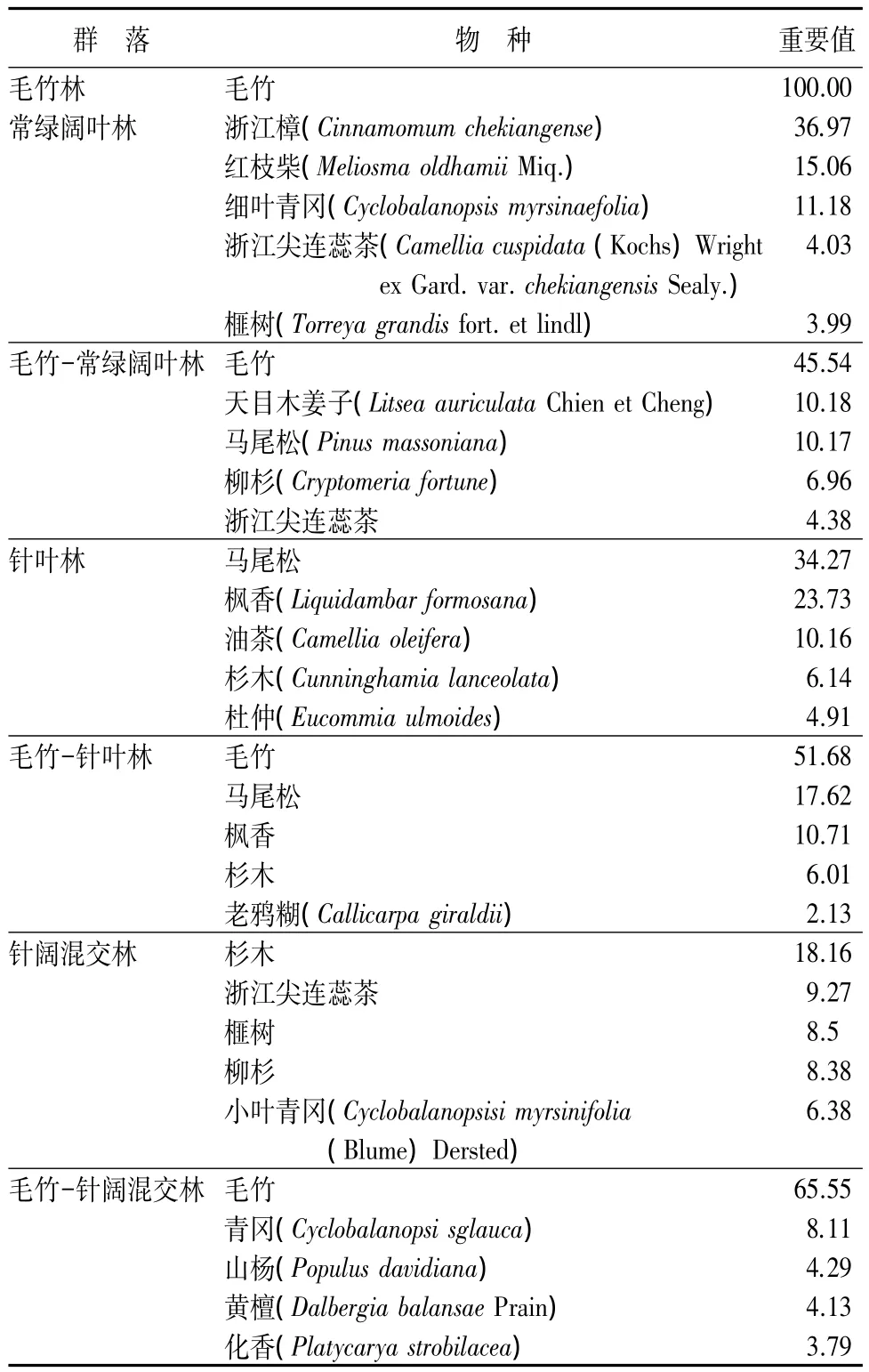
表3 不同群落乔木层主要物种重要值
由表3可知,毛竹林中毛竹占绝对优势;受毛竹扩张影响的毛竹-针阔混交林、毛竹-常绿阔叶林、毛竹-针叶林中毛竹亦占优势地位,其种群重要值分别为 65.55、45.54、51.68,明显高于其他物种;针阔混交林中杉木种群优势较明显,其重要值为18.16,其余物种重要值均小于10;常绿阔叶林中浙江樟优势明显,其重要值达36.97;针叶林中马尾松和枫香占优势,重要值分别为 34.27 和 23.73。
不同群落灌木层主要物种重要值(见表4)。毛竹林中调查仅得两个物种,分别为中国金菊花和华桑,其重要值为58和42;毛竹-针阔混交林、毛竹-常绿阔叶林中化香和中国绣球分别以16.72和30.59占得优势;毛竹-针叶林中青榨槭、腊莲绣球、茶优势明显,其重要值之和达80.94;针阔混交林中浙江连尖蕊茶优势较为明显,重要值为32.05;常绿阔叶林中细叶青冈和浙江连尖蕊茶其重要值之和为60.85,占主要优势;针叶林中小果蔷薇重要值为25.06,优势明显。

表4 不同群落灌木层主要物种重要值
不同群落草本层主要物种重要值(见表5)。毛竹林中箬竹是唯一的草本;毛竹-针阔混交林中箬竹以64.01的重要值占绝对优势;毛竹-常绿阔叶林中箬竹、山麦冬均以31.71的重要值占据优势;毛竹-针叶林中凤尾蕨占优势,重要值为19.27,但其重要值大于10的有三种;针阔混交林和常绿阔叶林中分别箬竹和苔草优势明显;针叶林中络石的重要值较高,为 28.23。
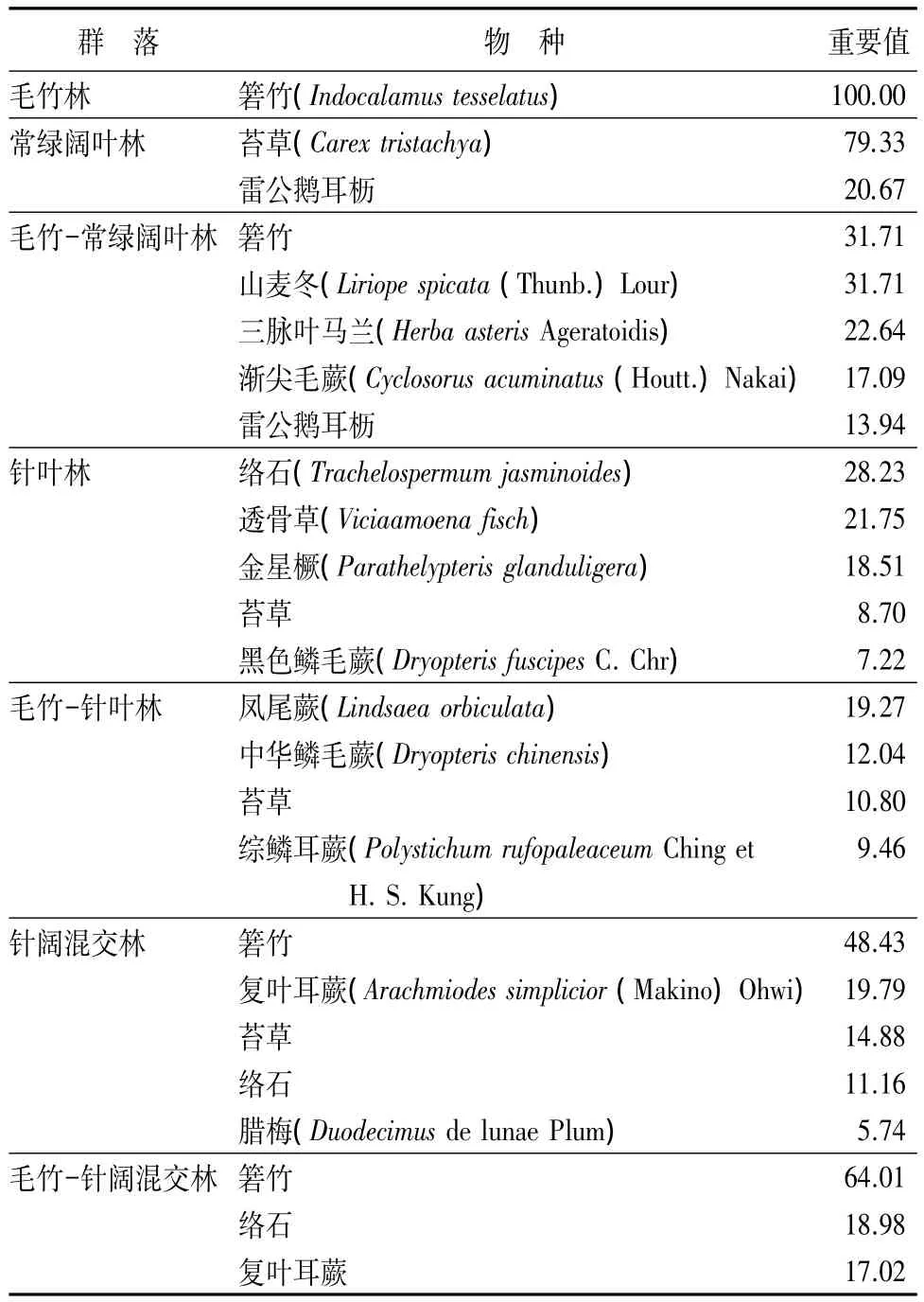
表5 不同群落草本层主要物种重要值
3.4 毛竹扩张对不同群落生物多样性的影响
通过对野外调查资料进行统计分析,天目山毛竹种群所处群落层次明显,可以分为乔木层、灌木层和草本层,地被层不发达。经过分析计算,本研究对所调查的7种不同林分的乔木层、灌木层和草本层进行了Patrick丰富度指数(R)、Simpson指数(D)、Shannon—Wienner指数(H)以及Pielou指数(J)的计算。
3.4.1 Patrick 丰富度指数(R)
Patrick丰富度指数(见表6)。乔木层中,受毛竹扩张影响的林分比未受影响的相应林分的R值均有所降低,毛竹的扩张对三种林分的物种数目均有一定影响。灌木层中,毛竹林的R值最低;毛竹-针阔混交林R值较高,毛竹-针叶林比针叶林的R值有所降低,毛竹-针阔混交林的R值则有所上升。在草本层中,毛竹林的R值最低;常绿阔叶林与毛竹-常绿阔叶林、针叶林与毛竹-针叶林、针阔混交林与毛竹-针阔混交林的R值均较为接近。
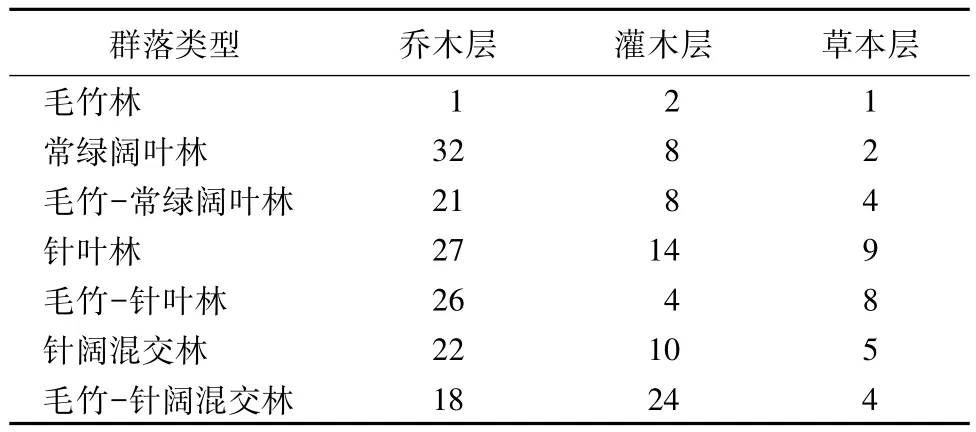
表6 不同群落的Patrick丰富度指数
3.4.2 Simpson 多样性指数(D)
不同样方乔木层D值的测度结果(见表7)。各群落的D值随毛竹的出现呈现下降趋势。其中毛竹-针阔混交林受影响趋势最为明显。灌木层和草本层中,D值变化趋势较为复杂。灌木层中,毛竹-常绿阔叶林和毛竹-针叶林受毛竹扩张的影响,D值均发生明显的下降,而毛竹-针阔混交林的D值比针阔混交林的D值出现上升趋势。草本层中,常绿阔叶林比毛竹-常绿阔叶林及针阔混交林较毛竹-针阔混交林得D值均有明显上升,毛竹-常绿阔叶林与常绿阔叶林的D值则较为接近。此外,毛竹纯林的乔木层、灌木层和草本层D值均为最低,毛竹生长对各层次多样性的影响较大。
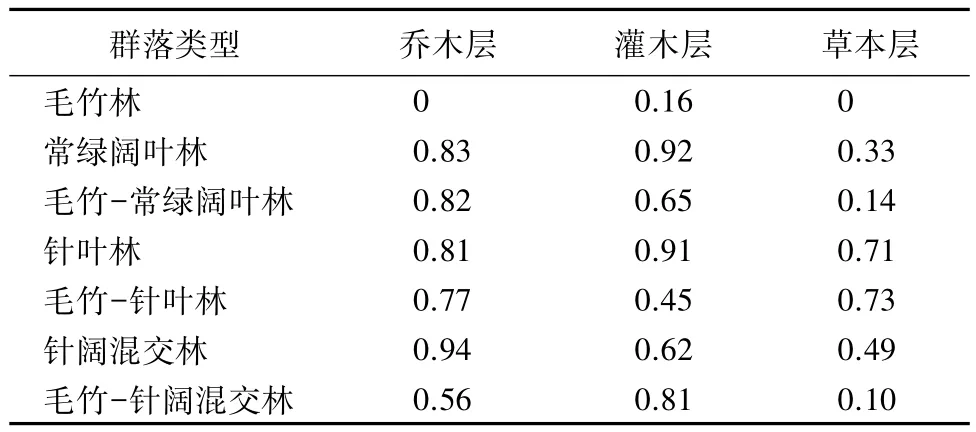
表7 不同群落的Simpson多样性指数
3.4.3 Shannon—Wienner多样性指数(H)
对比数据可得(见表8),乔木层、灌木层、草本层中毛竹的H值均为最低,与Simpson多样性指数(D)的计算结果一致。受到毛竹扩张影响的针阔混交林、常绿阔叶林和针叶林的H值,与未受到毛竹扩张影响的相应林分比较,毛竹扩张对针叶林的影响较为显著。灌木层中,毛竹-针阔混交林的H值最高,其值为1.88;毛竹-针叶林比针叶林的H值有所降低;常绿阔叶林和毛竹-常绿阔叶林的H值相差不大;毛竹-针阔混交林比针阔混交林H值高。在草本层中,毛竹-针叶林的H值最高,其值为1.35;毛竹-针阔混交林比针阔混交林的H值下降较大;而毛竹-常绿阔叶林与常绿阔叶林H值相差不大。
3.4.4 Pielou 均匀度指数(J)
从表9中可得,毛竹在各个层次的J值均为7个森林类型中最低。乔木层中,受毛竹扩张影响后的林分,J值均呈现下降趋势。在灌木层中,毛竹-常绿阔叶林与常绿阔叶林、毛竹-针叶林与针叶林相比,J值均有所降低。在草本层中,除毛竹-针叶林与针叶林的J值接近外,毛竹-常绿阔叶林比常绿阔叶林、毛竹-针阔混交林比针阔混交林的J值均有所下降。
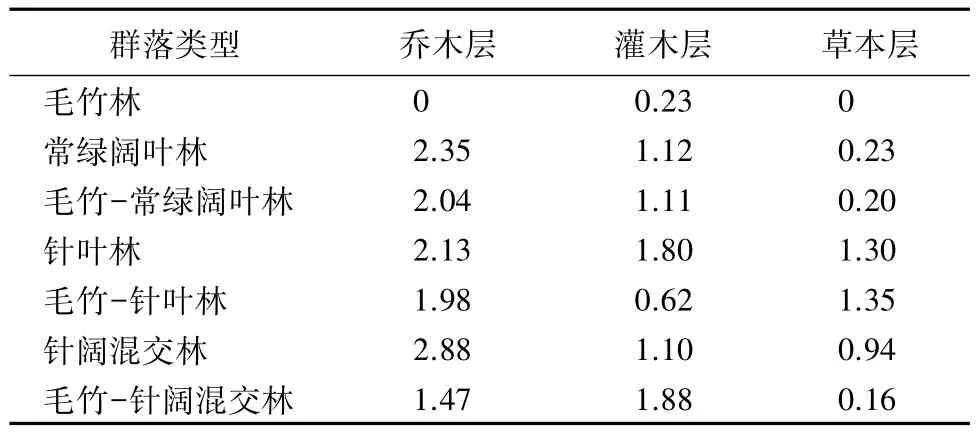
表8 不同群落的Shannon—Wienner多样性指数
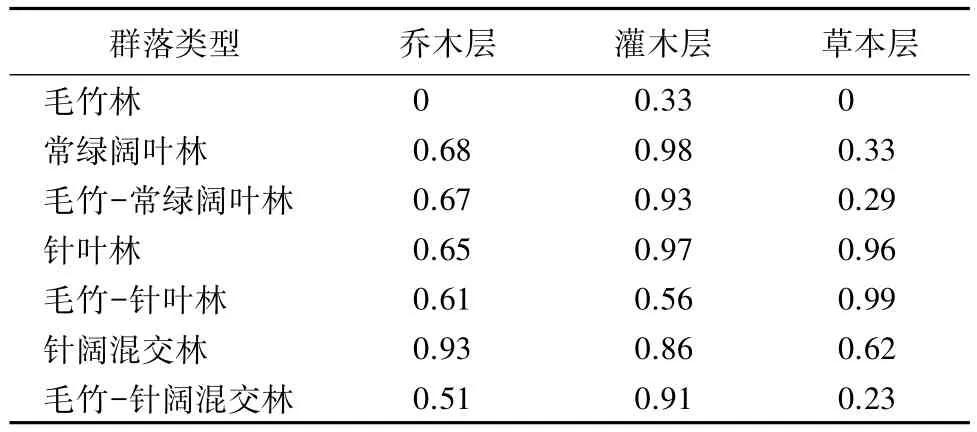
表9 不同群落的Pielou均匀度指数
4 结论与讨论
通过调查,天目山自然保护区毛竹林在多个群落类型中均有扩张现象。毛竹立竹度随着群落类型的改变而改变。在调查的7个群落中,毛竹的立竹度由大到小为:毛竹林、毛竹-针阔混交林、毛竹-常绿阔叶林、毛竹-针叶林,毛竹的入侵分别降低了相应的群落类型的总立木数。对于各群落类型的乔木层的多样性分析,毛竹的入侵分别降低了相应的群落类型的Patrick丰富度指数、Simpson多样性指数、Shannon—Wienner多样性指数和Pielou均匀度指数。对于灌木层,毛竹入侵使得对常绿阔叶林灌木层的R、D、H和J值影响不大,降低了针叶林的各多样性指数值,但却提高了针阔混交林相应的多样性指数值,变化趋势较复杂。对于草本层,毛竹入侵除对针阔混交林生物多样性指标影响较大外,对其他群落影响不大。综上所述,毛竹的入侵对群落乔木层的生物多样性影响较大;对灌木层和草本层影响较小,甚至在部分森林类型中其生物多样性有所增加。
杨怀等[11]对鸡公山自然保护区毛竹林扩张对植物物种组成和生物多样性的研究结果表明:物种的多样性和均匀度都随毛竹的出现呈现明显的下降趋势,反映出毛竹林在扩张过程中,对生物多样性的影响不仅表现在使物种丰富度的迅速下降和层次结构的逐渐简单化,而且使群落的物种组成也发生着显著变化。在天目山国家级自然保护区内,赵明水等[12]曾采用样带网格调查法,研究毛竹林向针阔混交林扩张的过程中对植物多样性的影响。研究表明毛竹蔓延使针阔混交林群落的植物多样性降低。其中,对乔木层植物多样性指数的影响最大,多样性呈现明显的下降趋势;灌木层植物多样性有增加的趋势;而草本层植物多样性在多数样带中,过度区相应较高。群落的均匀度指数大多随毛竹的出现而降低,反映出毛竹蔓延使群落中物种分配出现不均匀。本文通过对浙江省天目山国家级自然保护区7种不同群落类型的立木数、不同类型不同层次的物种丰富度、多样性指数及均匀度指数的调查及计算,得到毛竹林的扩张对针阔混交林、常绿落叶林、针叶林的物种多样性均有不同程度的影响。随着毛竹在不同林分中的不断扩张,各个群落类型的物种丰富度均有所降低,毛竹在所扩张林分中逐渐成为优势物种,并且毛竹的扩张有着将被扩张林分逐步演变为毛竹林的趋势,长期下去保护区最具特点的典型植被和其内丰富的生物多样性将消失。
因此,要正确认识毛竹林扩张对自然保护区内物种多样性的不利影响,在认真贯彻自然保护区的科学保护观,破除“绝对保护”的传统观点的基础上,可施行自然保护区核心区彻底清除毛竹林、实验区间伐毛竹林等人工干预措施,以控制自然保护区毛竹林的蔓延[5]。并制定自然保护区内资源可持续利用和管理的政策及其法规体系,从一草一木不能动转向发挥保护区多种价值,从单一管理类型转向多种管理类型[18]。保护区需加强毛竹林扩张的监测和控制,掌握其的种群变化动态[19],运用GPS等技术手段,对保护区毛竹分布点及范围进行定位,定期对毛竹扩张区域的种群数量变化进行监测,并建立相关数据库,适时对收集的数据进行分析和研究[20]。另外,保护区可借助自身科研实力优势,同时与相关科研单位合作,加强对保护区内外来入侵植物的调查研究,制定有效的防治技术与对策[21]。利用现有的手段阻止毛竹林在保护区的不断扩张,同时寻求更为合理的管理措施。
[1]兰思仁.武夷山国家级自然保护区植物物种多样性研究[J].林业科学,2003,39(1):36-43.
[2]蔡亮,张瑞霖,李春福,等.基于竹鞭状态分析的抑制毛竹林扩散的方法[J].东北林业大学学报,2003,31(5):68-70.
[3]吴家森,姜培坤,王祖良.天目山国家级自然保护区毛竹扩张对林地土壤肥力的影响[J].江西农业大学学报,2008,30(4):689-692.
[4]赵永艳,苏继申.竹阔混交林土壤性状与分析[J].南京林业大学学报:自然科学版,2007,31(1):81-84.
[5]杨淑贞,杜睛洲,陈建新,等.天目山毛竹林蔓延对鸟类多样性的影响研究[J].浙江林业科技,2008,28(4):43-46.
[6]詹敏,窦云鹏,郭培培,等.天目山不同森林类型林冠对酸雨的缓冲作用[J].浙江林业科技,2010,30(2):26-30.
[7]王兵,杨清培,郭起荣,等.大岗山毛竹林与常绿阔叶林碳储量及分配格局[J].广西植物,2011,31(3):342-348.
[8]丁丽霞,王祖良,周国模,等.天目山国家级自然保护区毛竹林扩张遥感监测[J].浙江林学院学报,2006,23(3):297-300.
[9]李燕华,白尚斌,周国模,等.自然保护区内毛竹竹鞭的动态生长研究[J].安徽农业科学,2010,38(18):9834-9835,9837.
[10]赵明水,刘亮,陆森宏,等.天目山织染保护区毛竹林扩张对植物多样性影响研究[C]//中国林学会,第二届中国林业学术大会:S8野生动物、湿地与自然保护区论文集.北京:中国林学会,2009:190-196.
[11]杨怀,李培学,戴慧堂,等.鸡公山毛竹扩张对植物多样性的影响及控制措施[J].信阳师范学院学报:自然科学版,2010,23(4):553-557.
[12]郑成洋,方精云,何建源.福建武夷山自然保护区地形对毛竹(Phyllostachys pubescens)林分布的影响[J].生物多样性,2004,12(1):75-81.
[13]杨清培,王兵,郭起荣,等.大岗山毛竹扩张对常绿阔叶林生态系统碳储特征的影响[J].江西农业大学学报,2011,33(3):529-536.
[14]朱长龙,上官林平.井冈山毛竹林扩边对生物多样性的影响初探[J].国土与自然资源研究,2009(3):45-46.
[15]马克平.生物群落多样性的测度方法I:α多样性的测度方法(上)[J].生物多样性,1994,2(3):162-168.
[16]马克平,刘玉明.生物群落多样性的测度方法I:α多样性的测度方法(下)[J].生物多样性,1994,2(4):231-239.
[17]白尚斌,周国模,王懿祥,等.天目山国家级自然保护区毛竹扩散过程的林分结构变化研究[J].西部林业科学,2012,41(1):77-82.
[18]韩念勇.中国自然保护区可持续管理政策研究[J].自然资源学报,2000,15(3):201-207.
[19]曹飞,宋小玲,何云核,等.惠州红树林自然保护区外来入侵植物调查[J].植物资源与环境学报,2007,16(4):61-66.
[20]刘峰,陶国达,王东升.纳板河自然保护区外来入侵植物状况调查及防范对策[J].林业调查规划,2008,33(6):112-114,117.
[21]宋小玲,曹飞,何云核,等.广东省鼎湖山国家级自然保护区外来入侵植物调查[J].浙江林学院学报,2009,26(4):538-543.
