不同氮水平对中美山杨幼苗生物量、氮浓度及其季节变化的影响1)
2014-09-14李海霞周志军张建瑛邢亚娟
李海霞,周志军,张建瑛,邢亚娟
(黑龙江省林业科学研究所,哈尔滨150081)
氮素既是植物生长最重要的结构物质,又是植物生理代谢中最活跃、无处不在的重要物质—酶的主要成分[1],所以氮素对植物生理代谢和生长有重要作用。它的供应量及其有效性制约着林木的生长速度及生长状态,比其他任何一种营养元素更能限制植物的生产力[2-3]。在氮养分胁迫条件下,植物能够通过自身调节使植物做出形态和生理上的适应性反应,以增加其在胁迫条件下的生存机会[4]。
中美山杨(PopulusdavidianaandP.tremuloides)是我国东北地区最重要的用材树种之一,本文以中美山杨幼苗为试验材料,研究不同供氮水平对中美山杨幼苗根、茎、叶生物量分配以及氮浓度随季节变化的影响,进一步明确不同供氮水平对中美山杨生长的影响机制。
1 材料与方法
1.1 材料来源
本试验所用苗木为两年生的中美山杨播种苗,试验在温室中进行。
1.2 试验设置
2012年4月将供试苗木根部用清水漂洗后,用0.5%高锰酸钾溶液消毒,再用蒸馏水漂洗3次。然后分别移至装有河砂的钵中栽培,每钵植苗1株。幼苗供给如下全营养液:4 mmol/L NH4NO3、1 mmol/L KH2PO4、1 mmol/L KCl、1 mmol/L CaCl2·6H2O、0.6 mmol/L MgSO4·7H2O、0.021 mmol/L FeCl3·6H2O、6μmol/L MnCl2·4H2O、0.016 mmol/L H3BO3、0.3μmol/L ZnCl2、0.3μmol/L CuCl2·2H2O、0.3 μmol/L Na2MoO4·2H2O,必要时用Ca(OH)2或H2SO4把pH值调整到5.5~6.0。5月末进行不同浓度的梯度处理。
在氮浓度处理中设置如下4个浓度,括号中为相应处理表示符号:1mmol/L(N1)、4 mmol/L (N4)、8 mmol/L (N8)、16 mmol/L (N16),通过调节NH4NO3浓度来实现,(其中N8为正常供氮浓度),每个处理均为5株。在进行氮浓度处理时,其他营养成分不变。每3天浇1次营养液,每次每钵浇50 mL,浇灌营养液在8:00~9:00进行。每日8:00~9:00和17:00~18:00分2次浇水,每次每钵约100 mL[5]。
1.3 测定指标与方法
对中美山杨幼苗进行季节性取样测定,7~9月份每月取样1次,共3次。每次每处理随机挑选3株苗木进行全株收获,测定根、茎和叶的鲜重,然后置于75℃烘箱里烘干至恒质量,分别测出干质量。将每株样品的根、茎和叶粉碎,3株作为3个重复,采用浓H2SO4-H2O2消煮,过滤,定容,用multiN/C2100s碳氮分析仪测定全氮含量。
2 结果与分析
2.1 氮浓度处理下中美山杨幼苗各部分生物量及其分配
生物量是光合产物积累的结果,氮的供应状况明显影响着植物对碳同化物质的分配格局。不同氮浓度处理明显影响中美山杨幼苗生物量的累积。由表1可见,7月份采样时,各个氮处理下幼苗根、茎和叶生物量均随N供给浓度的增加而增加,在高N供应下达最大,这三者导致了单株总生物量在高N供应下最大,为7.76g/株,8月份,各个N处理下的幼苗茎和叶生物量均随N供给浓度的增加而增加,在高N供应下达最大,由于根的生物量在N8下最大,导致单株总生物量在适量供氮下最大,为9.98g/株,超过N8水平总生物量稍有下降,为9.85g/株,下降了1.3%。9月份随着供N浓度的增加,中美山杨叶的生物量逐渐增加,供N浓度为8mmol/L时达到最大,为2.62g/株,比供N不足时(1.78 g/株)增加了47.1%。茎生物量在供N不足时(1mmol/L)值最小,为2.57g/株,并随着供N水平的增加而增加,在供N浓度为16mmol/L时达最大,为3.94g/株,增加了53.31%。叶和茎导致地上部分在供N为8mmol/L时最大,为6.15 g/株,但当超过正常供N时,地上生物量又变小(5.75 g/株),减少了6.5%。而根的变化有所不同,在供N浓度4mmol/L时达最大,为6.88 g/株。叶、茎和根这3个器官生物量的变化最终导致幼苗全株总生物量在1~16mmol/L的供N范围内随供N水平增加而增加,在16mmol/L时达最大,比低N供应增加了11.22%。
在不同季节,中美山杨各部分生物量分配比例不同。从7月份到9月份,分配到叶中的生物量比例逐渐降低,平均降低了12.5%,而根系的生物量比例N1和N4逐渐增加,N8和N16有所降低,总体上平均增加了3.75%。研究认为,根系和叶片的生长具有相关性[6],叶片作为碳的“源”,向根系提供碳和代谢所需的能量,而根系为地上部分提供养分和水分。

表1 不同N处理下山杨幼苗根、茎、叶和全株生物量及其分配 g
2.2 氮浓度处理下中美山杨根、茎、叶中的全氮浓度
2.2.1 氮浓度处理下中美山杨根系中的全氮浓度
氮是诸矿物质养分中限制植物生长最严重的大量元素之一,它直接限制着森林生产力。Jones[7]认为植物根系的氮吸收能力与根系内部含量负相关,当土壤养分含量非常丰富或者施肥量过高时,叶片和根系的氮含量与土壤养分供给量不成正比,但当土壤养分成为限制性资源时,根系养分含量和叶片养分含量也相应降低。
不同氮处理下中美山杨幼苗根系全氮浓度变化情况如图1。在整个生长阶段,随供氮水平提高,山杨幼苗根系中全氮浓度也明显提高,7、8、9月份不同处理间根系全氮浓度差异均呈显著水平(P值分别为0.00,0.00和0.00)。7月份和9月份在高N水平(16mmol/L)下根系氮浓度达到最大,分别为11.36mg/g和14.48mg/g,8月份则在正常供N下达最大为10.19mg/g。8月份不同处理下根系全氮浓度变幅为6.45~10.19 mg/g,高低相差1.5倍,9月份变幅为7.07~14.49mg/g,高低相差2.05倍。说明越到生长后期,不同处理之间差别越大。
从不同生长阶段来看,从7月份到9月份中美山杨根系氮浓度持续上升,在生长末期氮浓度平均达到生长初期的1.16倍,说明随着苗木的生长,根系中氮在不断的积累,浓度加大。根系是树木生长发育过程中最重要的营养器官,也是养分贮藏的重要器官,在生长末期大量的养分转移到根系中贮藏[8-9]。

图1 不同氮处理下中美山杨幼苗根系中全氮浓度
2.2.2 氮浓度处理下中美山杨茎中的全氮浓度
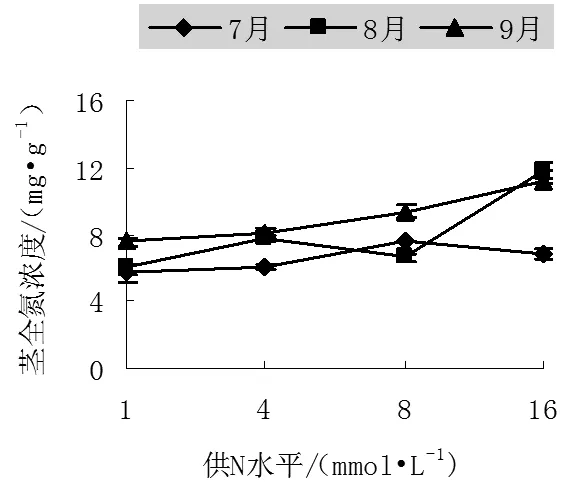
图2 不同氮处理下中美山杨幼苗茎中全氮浓度
不同季节山杨茎中全氮浓度表现为7月份不同处理之间变化不明显,8月份和9月份均是在高N处理下达最大,分别为11.7mg/g和11.21mg/g,比N1分别增加了95.01%和48.4%,N16是N1的1.98和1.48倍,这说明越到生长后期,不同处理间的差别越小,也就是说输送到地上部分的养分逐渐减少。
2.2.3 氮浓度处理下中美山杨叶片中的全氮浓度
叶片的光合作用是森林生产力的主要实现者。当养分供给速度降低时,单位重量叶片氮含量显著降低,并导致叶片光合能力显著降低[10]。增加土壤中的养分含量通常可改善叶片氮状态,并由此而提高叶片的光合作用。从试验的结果来看,不同供N浓度处理下山杨幼苗叶中全氮浓度差异显著,随着供N水平的增加,叶中全氮浓度增加,均在高N处理(N16)下达最大,其中7月份为18.32mg/g、8月份为17.22mg/g、9月份为20.17mg/g,比N1分别增加了39.73%、33.99%和45.38%。
不同季节叶片全氮浓度也有一定的变化。幼苗生长初期,N4处理下全氮浓度较高,8月份各个处理下全氮浓度相对于7月份有所下降,而到了生长末期(9月份)除了N4外其余各处理叶片中全氮浓度增加,N1、N8、N16比生长初期提高了1.06、1.05、1.1倍。
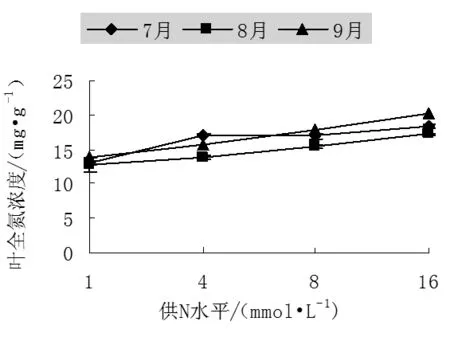
图3 不同氮处理下中美山杨幼苗叶中全氮浓度
3 讨论
3.1 氮素供应对幼苗生物量分配的影响
在自然条件下,土壤中的矿质营养有限或有效性较低,通常限制树木的生长发育。因此,在氮缺乏的土壤环境中,增加这些营养供应会促进树木生长。在本试验中,7月份和9月份幼苗总生物量均出现在高氮供应下,8月份幼苗生物量出现在适量供氮水平下。供N浓度增加能够提高山杨总生物量和各部分生物量,但各部分生物量比例会发生变化。
3.2 氮素供应对幼苗体内养分含量的影响
植物体内在一定环境下,植物的养分吸收量取决于根系的供应量和植物器官的生长和行使正常功能对养分的需要量[11]。Högberg等[12]认为,当土壤养分含量非常丰富或者施肥量过高时,叶片和根系的养分含量与土壤养分供给量不成正比。但是,当土壤养分成为限制性资源时,根系养分含量和叶片养分含量相应降低。
本试验发现,中美山杨体内养分浓度与外界养分供应浓度有着密切关系。总体来看,山杨根茎叶对外部养分供应状况反应都比较敏感,供氮浓度降低时,根、茎、叶中的全氮浓度也降低。随供氮水平的增加,叶片全氮浓度显著上升,8月份与氮水平呈正相关(r=0.992),整个生长季均是在高氮水平下最大。
[1]赵平,孙谷畴,彭少麟.植物氮素营养的生理生态学研究[J].生态科学, 1998(2).
[2]Cassman K G, Kropf M J, Gaunt J, et al. Nitrogen use efficiency of rice reconsidered: what are the key constraints? [J] Plant Soil, 1993, 155/156:359-362.
[3]Crawford N M, Glass A D M. Molecular and physiological aspects of nitrate uptake in plants [J]. Trends in plant Science, 1998, 3: 389-395.
[4]李春俭,张福锁.营养胁迫及其适应性反应的调节//冯锋,张福锁,杨新泉.植物营养研究-进展与展望[A].北京:中国农业大学出版社,2000.
[5]李海霞,李正华,戴伟男,等.氮磷水平对中美山杨幼苗碳氮积累与分配的影响[J].西南林业大学学报,2013,33(3):8-14.
[6]Hendricks JJ, Nadelhoffer KJ, Aber JD. Assessing the role of fine roots in carbon and nutrient cycling [J]. Trends Ecol Evol, 1993, 8:174-178.
[7]Jones, H.E., Hogberg, P., and Ohlsson, H.Nutrient assessment of a forest fertilization experiment in northern Sweden by root bioassays [J]. Forest Ecology and Management, 1991, 64:59-69.
[8]沈善敏,宇万太,张璐,等.杨树主要营养元素内循环及外循环研究Ⅰ.落叶前后各部位养分浓度及养分贮量变化[J].应用生态学报,1992,3(4):296-301.
[9]沈善敏,宇万太,张璐,等.杨树主要营养元素内循环及外循环研究Ⅱ.落叶前后各部位养分在植株体内外的迁移和循环[J].应用生态学报,1993,4(1):27-31.
[10]Carswell F. E., J.Grace, M.E.Lucas and Jarvis P.G. Interaction of nutrient limitation and elevated CO2 concentration on carbon assimilation of a tropical tree seedling (Cedrelaodorata) [J].Tree Physiology.2000, 20:977-986.
[11]Lauchli A, Bieleski RL.张礼中,毛知耘译.植物的无机营养[M].北京:农业出版社,1992:104-106.
[12]Peter Högberg, Lars Högbom, and Helga Schinkel.Nitrogen-related root variables of trees along an N-deposition gradient in Europe [J]. Tree Physiology, 1998, 18:823-828.
