BDNF对大鼠急性脊髓损伤后Bcl-2表达的影响
2014-09-12王岩峰秦光华张玉强杨明超
王岩峰 秦光华 张玉强 杨明超 王 喆
(中国医科大学附属第一医院骨科,辽宁 沈阳 110001)
脊髓原发性损伤后的继发性损伤扩大了损伤范围,加重了损伤程度,是脊髓损伤后影响脊髓修复的重要病理过程。脊髓继发性损伤的过程十分复杂,包括水肿、炎症反应、局部缺血、谷氨酸受体过度激活、脂质过氧化作用、钙离子超载等,但是目前尚无一个公认的关于脊髓损伤后继发伤的病理机制。脑源性神经营养因子(BDNF)对神经元的生存、分化、生长起重要作用。腺病毒介导BDNF基因移植可促进SCI 后红核脊髓神经元再生和修复,能激活再生相关基因表达〔1〕。SCI后,BDNF在损伤局部升高,可以促进脊髓神经元和轴突的再生〔2〕。Ikeda 等〔3〕发现,鞘内注射BDNF可提高SCI急性期内Cu/Zn超氧化物歧化酶(Cu/Zn SOD)和髓鞘碱性蛋白(MBP)在脊髓神经元和胶质细胞中的活性,从而对脊髓神经功能的恢复起积极作用。Bcl-2属于抑制细胞凋亡基因。潘磊等〔4〕研究发现,Bcl-2 蛋白不仅在Wistar成年大鼠神经元中提高表达;更多在胶质细胞中表达,同时降低Bax的表达;BBB评分表明,神经细胞凋亡明显减少,大鼠神经功能恢复明显。国外有学者证实MP疗法增强SCI 功能恢复的机制之一为提高Bcl-2 表达和抑制Bax 表达〔5〕。但是,外源性BDNF能否通过调节Bcl-2的表达参与急性脊髓损伤的发病机制尚未见文献报道。本研究旨在通过检测各组大鼠Bcl-2的表达,为治疗脊髓损伤提供实验依据。
1 材料与方法
1.1材料 羊抗大鼠Bcl-2、β-actin多克隆抗体(Santa- Cruz,CA);兔抗羊IgG、S-P免疫组化染色试剂盒、DAB显色试剂盒(福州迈新)。
1.2大鼠急性脊髓损伤动物模型的建立及分组 取健康成熟Wistar大鼠72只,雌雄不限,体重220~250 g,由中国医科大学实验动物中心提供。随机分为对照组,损伤组,BDNF组。模型制备:10%水合氯醛0.4 ml/100 g腹腔内注射麻醉,无菌原则下进行背正中切口,切除T8、T9椎板,暴露硬脊膜。按Nystrom法将35 g重物通过2.2 mm×5 mm弧形光滑金属垫片压迫该段脊髓后正中部5 min,分层缝合,碘伏消毒。脊髓损伤后分6、12 h,1、3、7 d 共5个检测时间点,每个时间点8只;BDNF组:损伤后,立即向硬脊膜下腔内注射BDNF,分为6、12 h,1、3、7 d共5个检测时间点,每个时间点8只;空白对照组8只,只做T9~T10椎板切除术。
1.3免疫组化 各组大鼠分别于脊髓损伤后各时间点取材,麻醉后剖胸,经左心室-主动脉插管,4%多聚甲醛灌流固定。以伤处为中心取1.5 cm脊髓组织,多聚甲醛浸泡固定。常规蜡块包埋,连续切片数张(纵切),片厚6 μm,每只大鼠取12张切片,按SP试剂盒说明书测定Bcl-2的表达:常规脱蜡至水,3%过氧化氢处理、高温修复、血清封闭、滴加一抗过夜(浓度均为1∶150),并同时用PBS代替一抗作为阴性对照,4 ℃孵育过夜、隔日滴加二抗孵育20 min、SABC 37℃孵育20 min,继而二氨基联苯胺(DAB)显色,镜下观察显色背景。以上各步骤间均用0.01 mol/L PBS漂洗3次,每次5 min,而后苏木素复染,蒸馏水冲洗,梯度酒精逐级脱水,二甲苯透明,中性树脂封片,光镜观察脊髓组织变化。
1.4Western 印迹免疫蛋白印迹 动物以10%水合氯醛0.4 ml/100 g腹腔内注射,麻醉后,各时间点分别取4只动物,直视下取出距损伤中心区上下各0.5 cm两段脊髓,置于裂解液中机械匀浆,4 ℃ 17 000 r/min离心1 h,吸出上清液,弃去沉淀。用考马斯亮蓝G250结合法,根据已知牛血清白蛋白溶液的浓度及其吸光度和样品吸光度计算出样品蛋白浓度和加样量。加入样品缓冲液,然后将样品置于沸水浴中加热5 min使蛋白质变性。将制备好的样品置于-20℃冰箱备用。制备12%分离胶,4%浓缩胶,用微量加样器将样品加在凝胶表面样品槽中。电泳后湿转至硝酸纤维素膜上。5%脱脂奶粉TTBS缓冲液4℃过夜。一抗Bcl-2、β-actin(1∶200)室温孵育2 h,洗膜,辣根酶标记羊抗兔IgG-HRP(1∶5 000,Santa Cruz)室温孵育1 h,洗膜后加入ECL试剂( Santa Cruz)反应1 min,暗室曝光显影后冲洗胶片。凝胶成像分析系统上摄像分析,测得目标带的IDV,进而计算出各组样品Bcl-2目标带的IDV与内参照β-actin IDV的比值。
2 结 果
2.1免疫组化结果 从免疫组化图片上可以看出,对照组Bcl-2蛋白阳性表达细胞数较少(11.06±0.65)。损伤组在手术后6 h开始出现大量的神经元和胶质细胞凋亡,与对照组相比,Bcl-2蛋白阳性表达的细胞数明显增加(P<0.05),并在3 d时达到峰值;而在之后逐渐减少,但在7 d时仍有一定数量的Bcl-2阳性表达的细胞。而相比于实验组的各时间点,BDNF组的大鼠脊髓Bcl-2阳性表达明显低于损伤组。见图1,表1。
2.2Western 印迹结果 与对照组相比(0.08±0.01),脊髓损伤组大鼠损伤后6、12 h,1、3、7 d的Bcl-2表达明显升高(P<0.05),且逐渐上升,3 d达高峰,而在之后逐渐减少,但在7 d时仍有大量表达;BDNF组6、12 h,1、3、7 d时间点的大鼠脊髓Bcl-2阳性表达明显低于损伤组(图2,表1)。

图1 免疫组化方法检测各组小鼠Bcl-2蛋白的表达(×200)
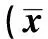
表1 各组小鼠Bcl-2蛋白光密度值及NF-κB灰度值比较±s,n=4)

图2 Western印迹方法检测各组小鼠Bcl-2和 β-actin蛋白表达
3 讨 论
传统观念认为,脊髓损伤后神经元和轴突不可再生,其主要原因是损伤局部环境缺乏营养物质和诱导轴突再生的蛋白因子〔6〕。但有研究发现,SCI后脊髓神经元和轴突仍具有一定的再生能力,但这种再生需要有神经营养因子(NTFs)的营养支持〔7〕。BDNF是NTFs家族中的成员之一,对神经元的生存、分化、生长起重要作用〔8〕。SCI后,BDNF在损伤局部的升高,可以促进脊髓神经元和轴突的再生〔9〕,目前已成为SCI 研究中的热点之一。
Bcl-2主要位于线粒体外膜、核被膜和内质网膜,可抑制细胞凋亡基因。有研究证实,米诺环素能提高SCI后神经功能恢复,其作用是增加Bcl-2 的表达,从而提高Bcl-2/ Bax 的比值,减少神经细胞走向凋亡,进而保护损伤的脊髓神经组织〔10〕。王金光等〔11〕证实β-七叶皂甙钠改善血循环,中止SCI 继发性损伤,抑制脊髓神经细胞凋亡和炎症表达的机制之一是上调Bcl-2 表达;还有研究发现,银杏叶提取物( EGb)能抑制SCI后细胞凋亡,在运动功能恢复、损伤脊髓组织保护上发挥其有益作用,其机制之一也是上调Bcl-2 和下调Bax 基因表达〔12~14〕。
本实验提示外源性BDNF很可能是通过上调Bcl-2的表达来参与急性脊髓损伤的发病机制的。
4 参考文献
1Kwon BK,Liu J,Lam C,etal.Brain-derived neurotrophic factor gene transfer with adreno-associated viral and lentiviral vectors prevent srubro spinal neuronal atrophy and stimulates regeneration-associated gene expression after cute cervical spinal cord injury 〔J〕.Spine,2007;32(11):1164 - 73.
2Gulino R,Lombardo SA,Casabona A,etal.Leve ls of brain-de rived neurotrophic factor and neurotrophin-4 in lumbar motoneurons after low-thoracic spinal cord hemisection 〔J〕.Bra in Res,2004;1013(2):174-81.
3Ikeda O,Murakami M,Ino H,etal.Effects of BDNF on compression induced spinal cord injury:BDNF attenuates down-regulation of superoxide dismutase expression and promotes upregulation of myelin basic protein expression〔J〕.Neuropathol Exp Neurol,2002;61:142-53.
4潘 磊,吕国华,雷 高,等.大剂量甲基强的松龙对大鼠脊髓损伤后神经细胞凋亡的影响〔J〕.郧阳医学院学报,2007;26(5):271-4.
5Kim YT,Caldwell JM,Bellamkonda RV.Nanoparticle mediated local delivery of methylprednisolone after spinal cord injury 〔J〕.Biomaterials,2009;30(13):2582 - 90.
6Weidner N,Grill RJ,Tuszynski MH.Elimination of basal lamina and the collagen “Scar” afte r spinal cord injury fails to augment c rticospinal tract regeneration 〔J〕.Exp Neuro,1999;160( 1):40- 50.
7Widenfalk J,Lundstromer K,Jubran M,etal.Neurotrophic factor and receptor in the immature and adult spinal cord after mechanical injury or kainic acid 〔J〕.J Neurosci,2001;21:3457-75.
8Lamballe F,Klein R,Barbcaid O.TrkC,a new member of the trk family of tyrosine protein kinases,is a receptor for neurotrophin〔J〕.Cell,1991;66(5):967-79.
9Gulino R,Lombardo SA,Casabona A,etal.Levels of brain-de rived neurotrophic factor and neurotrophin-4 in lumbar motoneurons after low-thoracic spinal cord hemisection〔J〕.Brain Res,2004;1013(2):174-81.
10Cheng G,Liang Q,Chen AM,etal.Effect of minocycline on expression of Bcl2,Bax and Bcl2xl in rats with spinal cord injury〔J〕 .Chin J Clin Rehabil,2006;10(24):181-3.
11王金光,郑启新,郭晓东,等.应用β-七叶皂甙钠治疗大鼠脊髓损伤的实验研究〔J〕.中国康复医学杂志, 2005;2(1):15-17,80.
12Ao Q,Sun XH,Wang AJ,etal.Protective effects of extract of Ginkgo biloba(EGb 761)on nerve cells after spinal cord injury in rats 〔J〕 .Spinal Cord,2006;44(11):662-7.
13张 烽,冯炜炜,周海涛.银杏叶提取物对大鼠急性脊髓损伤后神经功能恢复的作用及其机制〔J〕.中国脊柱脊髓杂志,2007;17(9):675-9.
14樊立宏,王坤正,程 斌.银杏叶提取物对兔脊髓缺血再灌注损伤的保护作用〔J〕.中西医结合学报,2006;4(2):181-4.
