秋水仙素注射大蒜花苞诱导四倍体的研究
2014-09-08薛小艳刘世琦郭会平孙亚丽于新会连海峰
薛小艳 刘世琦 郭会平 孙亚丽 于新会 连海峰
(山东农业大学园艺科学与工程学院,作物生物学国家重点实验室,农业部黄淮地区园艺作物生物学与种质创制重点实验室,山东泰安 271018)
秋水仙素注射大蒜花苞诱导四倍体的研究
薛小艳 刘世琦*郭会平 孙亚丽 于新会 连海峰
(山东农业大学园艺科学与工程学院,作物生物学国家重点实验室,农业部黄淮地区园艺作物生物学与种质创制重点实验室,山东泰安 271018)
为获得染色体加倍的大蒜植株,用0.05%、0.10%、0.15%和0.20%的秋水仙素注射苍山糙蒜和金蒜3号的花苞,并在不同生长调节剂组合的MS培养基上培养花苞内形成的气生鳞茎,当再生体系建立后,对根尖进行倍性鉴定。结果表明,MS基本培养基添加0.05 mg·L-1NAA和2.0 mg·L-1KT能诱导较多的气生鳞茎萌发,直径4~5 mm的气生鳞茎萌发率最高,且苍山糙蒜气生鳞茎的萌发率整体比金蒜3号高。秋水仙素注射花苞后,金蒜3号的花苞死亡率远远高于苍山糙蒜。随着秋水仙素浓度增大,花苞内形成气生鳞茎数不断降低,染色体加倍率升高。苍山糙蒜以0.20%秋水仙素浓度处理下四倍体诱导率最高,为7.69%;金蒜3号以0.15%秋水仙素浓度处理下四倍体诱导率最高,为11.76%。
大蒜花苞;气生鳞茎;组织培养;四倍体;秋水仙素
大蒜(Allium sativumL.)有性杂交困难,自然变异有限,多倍体育种成为无性繁殖大蒜作物改良品种的有效途径之一(郭瑞峰和咸丰,2011)。秋水仙素(colchicine)可通过抑制或破坏纺锤体的形成,使细胞不能分裂,但姐妹染色单体仍能最终分开,并进一步重组成一个染色体数目加倍的细胞核的途径诱导染色体加倍(周明龙,2012),因而作为较为理想的多倍体诱变剂广泛使用(Sun et al.,2009;Kulkarni & Borse,2010)。秋水仙素通常采用浸种、滴涂和注射生长点等方法诱导植物无性阶段产生多倍体(Takamura & Miyajima,1996;郭启高 等,2000)。
大蒜气生鳞茎是由蒜薹上花托的小鳞芽发育而成,通常每个花苞原基可形成10~40个气生鳞茎(张素芝和李纪蓉,2006;马雯 等,2011)。气生鳞茎体积小,数量多,带病毒程度低,易发育成独立的试管苗(张素芝和李纪蓉,2006)。相关研究表明,以气生鳞茎为外植体的大蒜愈伤组织途径或不定芽途径的组织培养再生体系均可获得染色体加倍效果,但不定芽途径的植株再生报道相对较少且存在技术障碍(马雯 等,2011;袁学军和李艳丽,2011)。
本试验以大蒜花苞为试验材料,通过秋水仙素注射花苞腔的方法,研究了秋水仙素对花苞及形成气生鳞茎数的影响,并采用不定芽途径离体组织培养法将花苞内形成的气生鳞茎培养成试管苗,以期获得四倍体大蒜植株,为大蒜多倍体诱导育种和新种质的创新提供新的途径。
1 材料与方法
1.1 试验材料
供试大蒜品种苍山糙蒜和金蒜3号。
1.2 试验方法
1.2.1 秋水仙素对田间大蒜花苞的注射处理 大蒜于2012年10月5日种植于山东农业大学园艺实验站,采用常规水肥管理。于2013年5月6日大蒜抽薹花苞初期,用秋水仙素对花苞腔进行注射处理:每个品种共处理80株花苞,分7:00、16:00两批,采用浓度为0.05%、0.10%、0.15%、0.20%的秋水仙素注射花苞腔内原基生长点,每次注射0.1 mL,每天1次,连续注射3次,对照用清水处理,3次重复。于2013年5月27日采收花苞内形成的气生鳞茎,于4℃贮藏15 d后进行大蒜气生鳞茎萌发的组织培养及再生植株的倍性鉴定。
1.2.2 气生鳞茎室内组织培养的植株再生 将用清水处理、4℃贮藏15 d的气生鳞茎剥皮,用75%酒精消毒30 s,无菌水冲洗3次,4%次氯酸钠消毒35 min,无菌水冲洗3次,将其接种于不同诱芽培养基中诱导芽萌发,计算气生鳞茎萌发率。
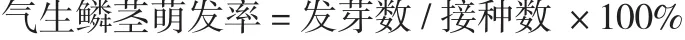
设置3种培养基进行诱芽培养基配方的筛选。1 号培养基:MS+NAA 0.05 mg·L-1+BA 0.1 mg·L-1(马雯 等,2011);2 号培养基:MS+NAA 2.0 mg·L-1+ BA 6.0 mg·L-1(张素芝和李纪蓉,2006);3号培养基:MS+NAA 0.05 mg·L-1+KT 2.0 mg·L-1。培养基添加30 g·L-1蔗糖和 7.5 g·L-1琼脂,pH 6.0,121℃灭菌 20 min。
将用秋水仙素处理、4℃贮藏15 d的气生鳞茎剥皮,将其接种于3号培养基中,研究气生鳞茎粒子大小对萌发率的影响。之后继续在3号培养基上培养形成再生植株。组培室培养条件为:(25±1)℃,光照强度 60 µmol·m-2·s-1,光照时间 16 h·d-1。
1.2.3 根尖染色体数目的鉴定 将组培苗培养60 d后,视组培苗的生根情况,取1~2 cm的根,参照文献(朱徽,1982;饶小珍 等,2003;孙奇 等,2011)进行预处理、固定、解离、染色。以及镜检的方法,经多次试验筛选出了适宜大蒜根尖染色体计数的直接压片鉴定法。
取气生鳞茎培养所得小苗的粗壮根系,每株取1~2个生长旺盛的根尖,在25℃饱和对二氯苯水溶液中预处理3 h,然后用现配制的卡诺固定液(无水乙醇∶冰乙酸=3∶1)在4℃下固定30 min,在95%酒精、85%酒精和70%的酒精中依次洗涤20 min后,放入70%的酒精中保存于4 ℃冰箱中待用;制片时,水洗净根尖后,用1 mol·L-1的盐酸在60℃条件下解离8 min,倒出盐酸,再加清水浸洗3次,最后放于清水中30 min后染色;染色时,切取根尖2~3 mm置于载玻片中,加1滴卡宝品红染液,染色5 min后,盖上盖玻片,压片,在荧光显微镜下观察染色体数目。
所有试验数据采用 DPS 6.55 和 Excel 2003 进行统计分析。
2 结果与分析
2.1 不同诱导培养基对气生鳞茎萌发率的影响
由表1可知,不同培养条件下气生鳞茎萌发率差异极显著。气生鳞茎在3号培养基(MS+NAA 0.05 mg·L-1+KT 2.0 mg·L-1)萌发率最高,苍山糙蒜萌发率高达75.91%,金蒜3号萌发率高达63.59%。

表1 不同诱导培养基对气生鳞茎萌发率的影响

图1 大蒜气生鳞茎在不同培养基上的发育状况
气生鳞茎在1号培养基上发育异常,从根部产生芽,不适宜气生鳞茎的正常萌发(图1-a、A);气生鳞茎在2号培养基上发育正常,部分根部产生芽,部分茎尖发芽,有部分气生鳞茎正常萌发(图1-b、B);气生鳞茎在3号培养基上发育正常,茎尖发芽,适于气生鳞茎的正常萌发(图1-c、C)。
2.2 粒子大小对气生鳞茎萌发率的影响
无菌条件下,将气生鳞茎培养于3号培养基上,结果发现,气生鳞茎萌发不仅受培养环境的影响,还受气生鳞茎粒子大小的影响。粒子较小和较大都不利于气生鳞茎的萌发,粒子直径4~5 mm时,气生鳞茎萌发率最高,且苍山糙蒜的气生鳞茎萌发率整体比金蒜3号高(图2)。
2.3 大蒜气生鳞茎组织培养条件下的发育变化
无菌条件下,将气生鳞茎接种于3号培养基(图3-a、A)上;培养 7~10 d 后茎尖转绿(图 3-b、B);约经过 20 d,茎尖发芽(图 3-c、C);60 d后生根成苗(图3-d、D)。
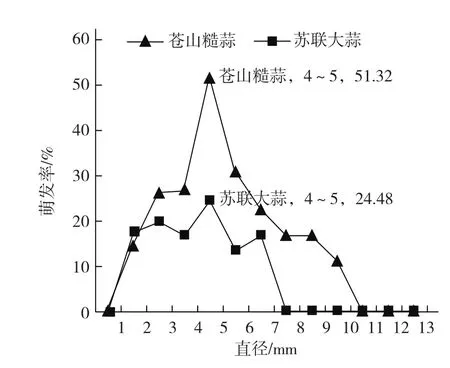
图2 粒子大小对气生鳞茎萌发率的影响

图3 大蒜气生鳞茎的植株再生
2.4 秋水仙素对花苞及形成气生鳞茎数的影响
由表2可知,秋水仙素注射花苞后,金蒜3号的花苞死亡率远远高于苍山糙蒜,且随着秋水仙素浓度的增大,花苞内形成的气生鳞茎数随之减少,直径4~5 mm苍山糙蒜气生鳞茎粒子率随之有所降低,直径4~5 mm金蒜3号气生鳞茎粒子率先升高后降低。可见秋水仙素通过影响粒子直径大小间接影响了气生鳞茎的萌发率。
2.5 秋水仙素对苍山糙蒜和金蒜3号的倍性诱导效应
由表3可以看出,0.15%秋水仙素注射苍山糙蒜花苞对染色体加倍的诱导率低于金蒜3号,不同浓度秋水仙素处理的气生鳞茎的倍性变化规律基本一致,即随着秋水仙素浓度增大,二倍体率降低,嵌合体率升高,四倍体率也升高(0.20%秋水仙素处理金蒜3号除外)。苍山糙蒜以0.20%秋水仙素浓度下四倍体诱导率最高,为7.69%;金蒜3号以0.15%秋水仙素浓度下四倍体诱导率最高,为11.76%。
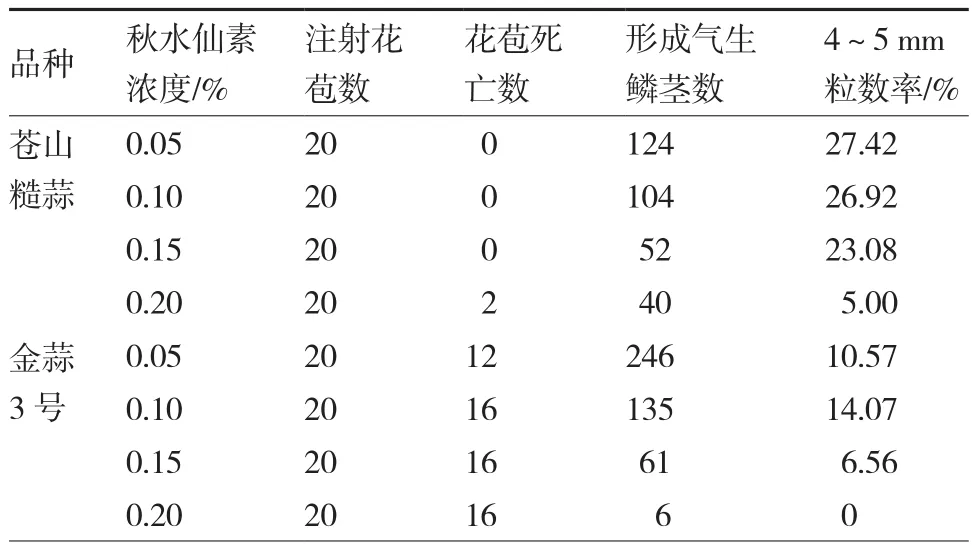
表2 秋水仙素对花苞及形成气生鳞茎的影响
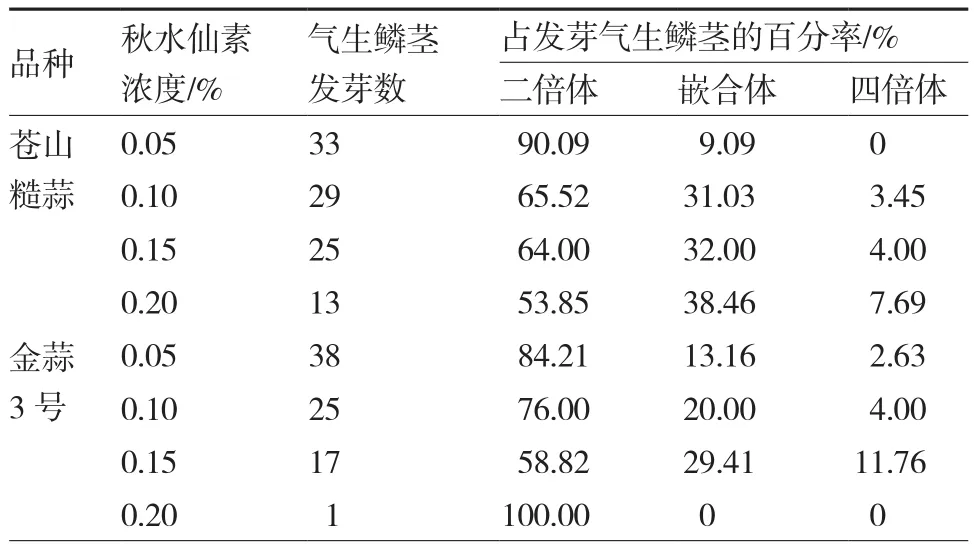
表3 秋水仙素对花苞内气生鳞茎的诱导变异
由图4可以看出用根尖压片法镜检观察染色体条数,气生鳞茎得到的小苗,不仅有16条染色体的二倍体植株(图4-a)和32条染色体的四倍体植株(图4-c),还有一些嵌合体(图4-b)。
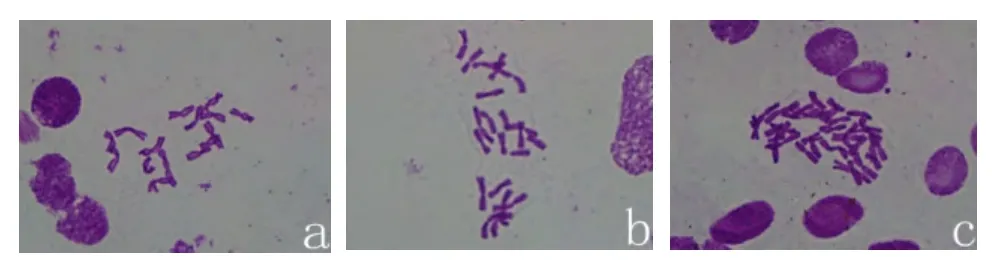
图4 二倍体、嵌合体和四倍体植株的染色体
3 结论与讨论
本试验通过不同培养基配方筛选发现,不同生长调节剂组合的MS培养基对气生鳞茎的萌发影响较大。气生鳞茎在1号培养基(MS+NAA 0.05 mg·L-1+BA 0.1 mg·L-1) 上 发 育 异 常, 从根部产生芽,不适宜气生鳞茎的正常萌发;气生鳞茎在 2 号培养基(MS+NAA 2.0 mg·L-1+BA 6.0 mg·L-1)上部分发育正常,茎尖发芽,但正常萌发率较低。这与张素芝和李纪蓉(2006)、马雯等(2011)的研究结果有所不同,可能是因为大蒜品种间有差异,对气生鳞茎的萌发造成不同的影响。气生鳞茎在 3号培养基(MS+NAA 0.05 mg·L-1+ KT 2.0 mg·L-1)培养条件上发育正常,茎尖发芽,正常萌发率最高,苍山糙蒜萌发率达75.91%,金蒜3号萌发率达63.5%。2个品种间萌发率的差异可能是由于品种间差异造成的,比如生理休眠期因素(陈沁滨 等,2007)。
大蒜气生鳞茎萌发不仅受培养环境的影响,还受粒子大小的影响。粒子较小和较大都不利于气生鳞茎的萌发,粒子直径为4~5 mm时,萌发率最高。不同大小的气生鳞茎萌发率差异很大,这与气生鳞茎的休眠情况不同有关。较小的气生鳞茎由于种皮障碍,成熟度差,休眠程度高,不易打破(杨期和 等,2003)。较大的气生鳞茎发育较早,进入了生理休眠期,需满足冷温需求量才能解除休眠(徐红艳 等,2013)。直径4~5 mm的气生鳞茎,未受到种皮障碍,也未进入生理休眠期,尚处于养分积累和鳞茎形成期,其体内物质代谢旺盛,极易萌发。
采用秋水仙素注射苍山糙蒜和金蒜3号花苞,二者均获得了一定频率的四倍体试管苗。经过比较发现,秋水仙素注射金蒜3号的花苞死亡率远远高于苍山糙蒜,这可能是因为金蒜3号花苞和苍山糙蒜花苞对秋水仙素的敏感程度不同所致。秋水仙素的毒害性较大,如果处理不当,不仅无法使植物细胞染色体加倍,还会对处理材料造成严重的毒害作用,甚至使处理材料死亡,对诱变效率有较大的影响(郑永强和徐坤,2003)。因此需要针对不同材料筛选合适的处理浓度。
苍山糙蒜以0.20%秋水仙素浓度下四倍体诱导率最高,为7.69%;金蒜3号以0.15%秋水仙素浓度下四倍体诱导率最高,为11.76%。此次试验获得了四倍体大蒜植株,为大蒜多倍体诱导育种和新种质创新提供了新的途径。
秋水仙素注射花苞,形成的气生鳞茎经组织培养获得试管苗,用根尖压片法镜检观察染色体条数,发现不仅有16条染色体的二倍体植株和32条染色体的四倍体植株,还有一些染色体重复加倍或有丝分裂不平衡造成的嵌合体。由于大蒜属于无性繁殖作物,有性繁殖困难,导致其遗传背景狭窄,新品种选育困难,获得的嵌合体材料可通过组织培养技术进行大量繁殖,为大蒜新品种选育提供新的途径(徐启江 等,2008)。
目前,大蒜多倍体诱导主要通过人工途径中的化学方法,包括活体诱导法和离体诱导法。与经离体诱导的多倍体相比,直接处理生长点使其萌发的活体诱导法具有操作简单且适于推广的优点,但其多倍体诱导率较离体诱导法低。采用直接注射花苞腔初期不定芽生长点与注射隐匿于贮藏叶中的生长点相比,诱导效果更好,但诱变剂的选取,处理材料,诱变剂浓度﹑剂量及处理技术环节上还有待于进一步探索与完善(温艳斌和程智慧,2012)。
陈沁滨,侯喜林,王建军,韩建明.2007.不同熟性洋葱休眠期生理生化的变化.园艺学报,34(1):221-224.
郭启高,张钟灵,周虹,梁国鲁,宋明.2000.秋水仙碱诱导生姜多倍体的研究.西南农业大学学报,22(5):400-402.
郭瑞峰,咸丰.2011.大蒜育种技术研究进展.安徽农业科学,39(5):2617- 2619.
马雯,李唯,李金娟.2011.贮藏时间和外源激素对大蒜气生鳞茎形成的影响.湖南农业科学,(19):41-44,56.
饶小珍,许友勤,陈寅山,高如承.2003.西施舌的核型分析.动物学杂志,38(2):2-5.
孙奇,刘世琦,张涛,陈娴,孟凡鲁.2011.贮藏时间对大蒜再生植株的影响.中国蔬菜,(4):72-77.
温艳斌,程智慧.2012.大蒜多倍体诱导及其研究进展.中国蔬 菜,(22):8-16.
徐启江,陈青奇,陈典.2008.利用茎尖离体嫁接获得大蒜体细胞嵌合体.园艺学报,35(1):65-70.
徐红艳,徐迎春,董凤祥,张清华,王彦辉.2013.夏果型树莓和黑莓品种需冷量及药剂打破休眠效果的研究.中国农学通报,29(16):119-124.
杨期和,叶万辉,宋松泉,殷寿华.2003.植物种子休眠的原因及休眠的多形性.西北植物学报,23(5):837-843.
袁学军,李艳丽.2011.大蒜气生鳞茎愈伤诱导和植株再生.山西农业大学学报:自然科学版,31(1):61- 65.
张素芝,李纪蓉.2006.影响大蒜气生鳞茎萌发的因素分析.四川农业大学学报,24(2):148 -151.
郑永强,徐坤.2003.秋水仙素在植物体细胞染色体加倍中的应用研究进展.中国农学通报,19(5):89-91.
周明龙.2012.秋水仙素诱导染色体数目加倍的机制.生物学教学,37(8):49-50.
朱徽.1982.植物染色体及染色体技术.北京:科学出版社:1-93.
Kulkarni M,Borse T.2010.Induced polyploidy with gigas expression for root traits inCapsicum annuum(L.).Plant Breeding,129(4):461-464.
Sun Q,Sun H,Li L,Bell R L.2009.In vitrocolchicine-induced polyploid plantlet production and regeneration from leaf explants of the diploid pear(Pyrus communisL.)cultivar‘Fertility’.The Journal of Horticultural Science & Biotechnology,84(5):548-552.
Takamura T,Miyajima I.1996.Colchicine induced tetraploids in yellow-flowered cyclamens and their characteristics.Scientia Horticulturae,65(4):305-312.
Studies on Tetraploid Induction of Garlic Bud by Colchicine Injection
XUE Xiao-yan,LIU Shi-qi*,GUO Hui-ping,SUN Ya-li,YU Xin-hui, LIAN Hai-feng
(CollegeofHorticultureScienceandEngineering,ShandongAgriculturalUniversity,StateKeyLaboratoryofCropBiology,KeyLaboratoryofBiologyandGeneticImprovementofHorticulturalCrop(HuanghuaiRegion),MinistryofAgriculture,Tai’an271018,Shandong,China)
In order to acquire tetraploid garlic(Allium sativumL.),diploid garlic buds were injected with 0.05%,0.10%,0.15% and 0.20% colchicine,respectively.Then the aerial bulbils in garlic buds were cultured in MS medium combined by different growth regulators.After a regeneration system was constructed,polyploid appraisal was conducted on the root tip.The result indicated that more aerial bulbils were able to germinate on the MS basic culture medium adding 0.05 mg·L-1NAA and 2.0 mg·L-1KT.The aerial bulbils with 4-5 mm diameter had the highest germination rate.The germination rate of ‘Cangshancaosuan’ bulbils was higher than that of ‘Jinsuan No.3’.When the garlic buds were injected with colchicines,the mortality of ‘Jinsuan No.3’ bud was much higher than that of ‘Cangshancaosuan’.The number and size of the formed aerial bulbils were decreased and the rate of doubled chromosome was increased along with the increase of colchicine concentration.The highest doubling rate of ‘Cangshancaosuan’ was obtained by the treatment of injecting the buds with colchicine concentration at 0.20%,and its induction rate was 7.69%.The highest doubling rate of ‘Jinsuan No.3’ was obtained by injecting the buds with colchicine concentration at 0.15%,then its induction rate could reach the highest - 11.76%.
Garlic bud;Aerial bulbil;Tissue culture;Tetraploid;Colchicine
薛小艳,硕士研究生,专业方向:蔬菜栽培生理与分子生物学,E-mail:xuelinyan@163.com
*通讯作者(Corresponding author):刘世琦,教授,博士生导师,专业方向:蔬菜栽培生理与分子生物学,E-mail:liusq99@sdau.edu.cn
2013-10-17;接受日期:2013-12-03
“十二五”农村领域国家科技计划项目(2013BAD01B04)
