不同淡水水位对桐花树幼苗生长的影响
2014-08-28刁俊明刘惠娜曾宪录陈桂珠
刁俊明, 刘惠娜, 曾宪录, 陈桂珠
(1. 嘉应学院生命科学学院,梅州 514015; 2. 中山大学环境科学与工程学院,广州 510275)
红树林作为热带、亚热带海岸和河口潮间带的木本植物群落,由它组成的红树林湿地生态系统具有巨大生态效益[1].作为红树林重要物种之一,桐花树(Aegicerascorniculatum)(紫金牛科蜡烛果属)在我国广泛分布于东南沿海地区[2].近年来,诸多学者在桐花树林的动植物群落特征、抗性生理[3]、化学成份[4]及其对重金属、污水、富营养物等污染环境的适应、治理和各影响因素对红树林湿地系统中红树植物的生理生态特性影响等开展了研究[5-7].不同的红树林植物具有不同的耐淹水性[8],桐花树抗水淹能力强于秋茄(Kandeliacandel)、木榄(Bruguieragymnorrhiza)但不及白骨壤(Avicenniamarina)[9],潮汐浸淹时间是影响红树林生长的重要限制因子[10].廖宝文等[11]在野外(或模拟)海水潮汐浸淹条件下,发现桐花树忍耐海水淹浸能力强,树体的1/ 3浸入水中达3年仍可正常生长.作者的前期研究表明桐花树对淡水环境也具有较强的适应性[12-14].然而对桐花树在不同淡水水位条件下的生长情况了解尚少.本文采用不同淡水水位处理2月龄桐花树幼苗1年,探讨其在淡水淹浸胁迫下的生理生态响应,为其在淡水河流、湖泊和水库的消落带中应用提供科学依据.
1 材料与方法
1.1 试验材料
2009年10月22日,采集广东省梅州市嘉应学院生命科学学院红树林植物种植基地桐花树母树周围的2月龄幼苗,选取株高9~10 cm,节数1~2个,叶片数2~3片的幼苗为试验材料.
1.2 试验方法
幼苗种植在5个水池中,水池为10 m×2.5 m×0.8 m的长方形,土壤为35 cm厚的塘泥.每个水池的一端安装进水水龙头,相对的另一端安装出水水龙头,使水池的水位保持恒定.每个水池种植30株幼苗,共种植150株幼苗.每个水池内淹水位以淹浸茎高计算,分别设置0、1/4、2/4、3/4、4/4(即整株淹没)水位.其中,0水位为土壤表面不积水的处理.在雨天的时候及时排放和晴天时通过控制进出水水量,使水位保持设置的高度.淹水的水位设定是根据植物的生长高度变化而调整,每90 d调整1次.每60 d对土壤进行施肥1次,用0.5%复合肥溶液施入水体中.设定每个水位10株标样苗供定期测定生长指标之用.在全光照自然条件下淡水种植桐花树幼苗1 年.
1.3 幼苗生长测定
在试验0 d和90 d测定5个水位组10株标样苗的茎高、茎径、胚轴高、胚轴径、分枝数、叶片数;采用剪纸法随机测定第 3节叶片的叶面积.
在0 d 和360 d,5个水位组分别随机选取10株幼苗测定根、胚轴、茎、叶的干质量及总干质量,观察植株生物量结构的变化情况.
每90 d记录1次幼苗存活株数和死亡株数及死亡症状等,并计算出存活率.
1.4 几种生理指标的测定
每90 d采取桐花树标样苗成熟叶片,剪去粗大的叶脉并剪成碎块等前期处理后,参照文献[5]的方法测定过氧化物酶(POD)活性,四氮唑蓝法测定超氧化物歧化酶(SOD).参照张志良的方法[15],用蒽酮比色法测定可溶性糖含量.根系活力的测定,采用α-萘胺法.每个处理重复3次.
1.5 数据统计分析
采用Excel和SPSS 13.0软件对所得数据进行单因素方差分析(one-way ANOVA),若差异达到显著,则进行Duncan多重比较,P<0.05 为差异显著.
2 结果与分析
2.1 淡水淹水位对桐花树幼苗生长的影响
2.1.1 对桐花树幼苗存活率的影响 淡水淹水位试验180 d时,与0水位相比,1/4、2/4水位的幼苗成活率(图1)差异不显著,但均显著地大于3/4和4/4水位的成活率;360 d时,1/4、2/4水位的幼苗成活率显著大于0水位,同时大于3/4和4/4水位.
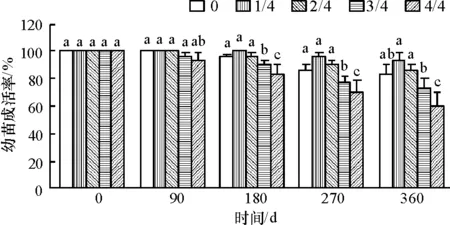
图1 不同淡水淹水位下桐花树幼苗的存活率
2.1.2 对桐花树幼苗茎生长的影响 1/4、2/4、3/4水位幼苗的茎高年增量(表1)显著增高,整株淹没的茎生长降低.1/4、2/4水位均对2月龄桐花树幼苗纵向生长和横向加粗生长均有促进作用,其中,1/4水位的茎高年增量和茎径年增量均显著大于0水位的.2/4水位的茎径年增量与0水位之间不存在显著差异.而3/4和4/4水位则对幼苗径的纵向生长和横向加粗生长均有抑制作用,4/4水位的抑制显著.
1/4 和 2/4水位在各个时期的茎高和茎径增量没有差异(图2和图3),从180 d起其茎高和茎径均显著大于0、3/4和4/4水位的茎高和茎径降低,说明随着淹浸时间的延长,高水位处理不利于茎的生长.1/4和2/4水位适合桐花树幼苗的生长.
2.1.3 对桐花树幼苗胚轴生长的影响 不同淡水水位处理360 d时,与0水位相比,1/4、2/4水位幼苗的胚轴高年增量和胚轴直径年增量差异不显著,但它们均显著地大于3/4和4/4水位.可见,3/4和4/4水位抑制胚轴生长(表2).
2.1.4 对桐花树幼苗分枝数、叶片生长的影响 不同淡水淹水位处理360 d后,1/4、2/4水位的平均每株分枝数与0水位差异不显著,显著地大于3/4和4/4水位的(表3),其叶片数叶长、叶宽和叶面积增加显著,其中叶面积达到极显著差异.可见,在高水位3/4、4/4淹浸下桐花树幼苗反应最强烈,叶片数减少.

表1 不同淡水淹水位下桐花树幼苗茎高、茎径年增长量
注:表中数据为平均值±标准差,同列不同小写字母间表示差异显著(P<0.05),反之则差异不显著,下表同.
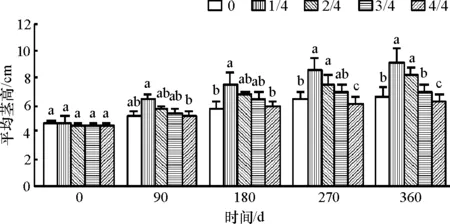
图2 不同淡水淹水位对桐花树幼苗茎高的影响

图3 不同淡水淹水位对桐花树幼苗茎径的影响
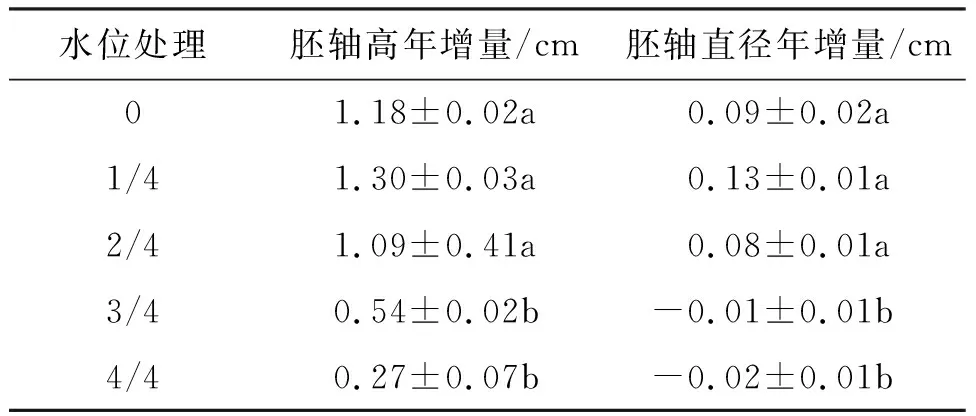
表2 不同淡水淹水位下桐花树幼苗胚轴高、胚轴直径年增长量
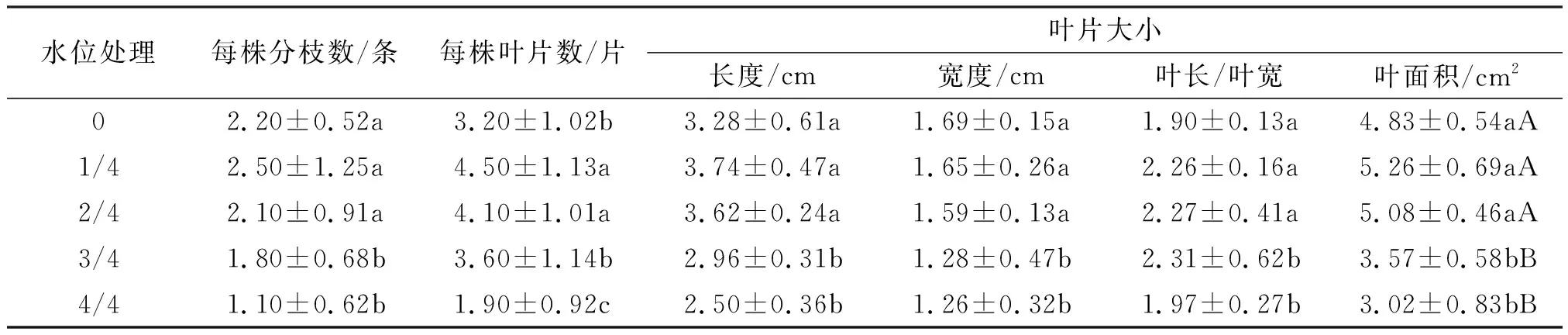
表3 不同淡水淹水位处理对桐花树幼苗分枝数和叶片生长的影响(360 d)Table 3 Effect of fresh water level on branch number, and leaf growth of Aegiceras corniculatum seedlings(360 d)
2.1.5 对桐花树幼苗生物量的影响 不同淡水淹水位组幼苗的根、茎、叶和胚轴生物量大小一致地表现为:叶>胚轴>茎>根(表4).从表4可知,360 d时,根生物量表现为1/4、2/4、3/4水位的均显著大于0和4/4水位的;而1/4、2/4、3/4水位的茎、胚轴、叶和单株的生物量增加,1/4和2/4水位的单株生物量大于0水位,3/4和4/4水位降低单株生物量.4/4高水位淹浸使生物量显著下降.
2.2 淡水淹水位对2月龄桐花树幼苗生理的影响
2.2.1 对桐花树幼苗过氧化物酶和超氧化物歧化酶活性的影响 叶片的过氧化物酶(POD)和超氧化物歧化酶(SOD)活性对不同淹水位的反应规律一致(表5),均表现为随淹水位增大而呈显著升高的趋势.3/4、4/4水位的POD和SOD酶活性均极显著大于0、1/4、2/4水位的,且4/4水位的POD和SOD酶活性分别是1/4水位的2.60倍和3.76倍.可见,高水位淹浸可使幼苗叶片的POD和SOD活性显著升高.
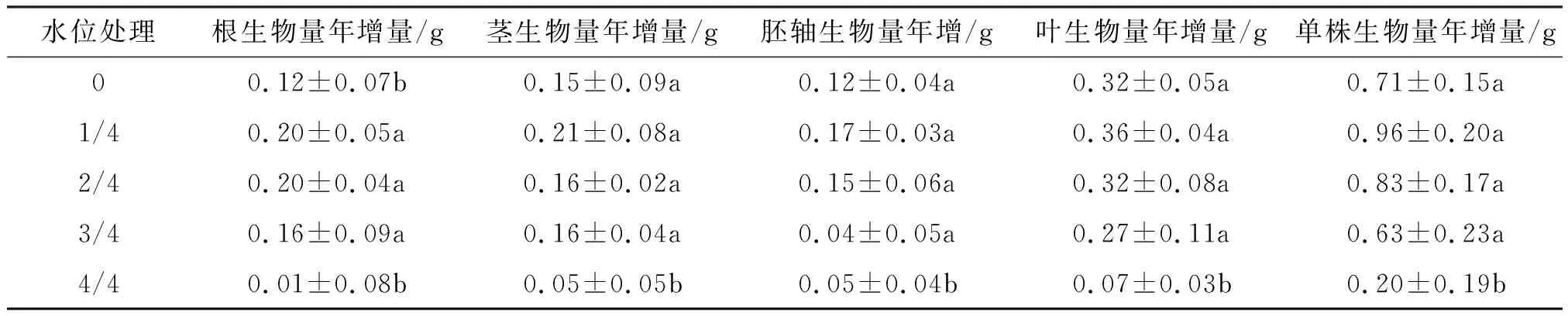
表4 不同淡水淹水位处理对桐花树幼苗生物量的影响
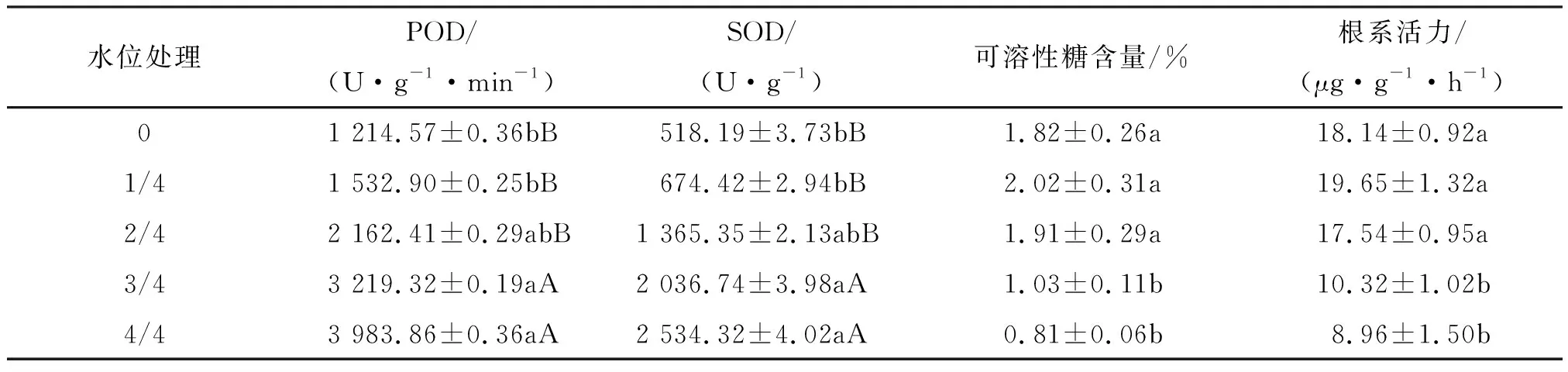
表5 不同淡水淹水位处理下幼苗叶片的过氧化物酶、超氧化物歧化酶活性、可溶性糖含量和根系活力的变化
2.2.2 桐花树幼苗叶片可溶性糖含量和根系活力的影响 叶片可溶性糖含量和根系活力均表现为2/4以下中低水位组显著地高于高水位组(表5).3/4、4/4水位的可溶性糖含量低于0水位,比1/4和2/4水位分别显著地降低.根系活力表现为1/4水位最大,与0和2/4水位差异不显著.3/4、4/4水位的根系活力显著地降低说明高水位的淹浸胁迫可使根系活力显著下降,抑制幼苗根系的生长,影响桐花树光合产物的积累.
3 讨论
本试验以0水位作为对照,淹浸水位的适宜升高(1/4和2/4水位)对2月龄桐花树幼苗的生长具有促进作用,但淹浸水位达到3/4以上不利于幼苗的生长,并且随淹浸时间的延长其起抑制作用更加显著,4/4水位的成活率显著下降.
由于红树植物被潮汐周期性浸淹,高长度茎可降低缺氧危害的程度;同时,加快茎高生长可延长叶片光照时间,增加光合作用有机产物[16].高水位促进大红树(Rhizophoramangle)幼苗茎高的增长[17];最低潮区滩涂上的大红树幼苗早期茎生长最快[18-19].而正红树(R.apicula-ta)的茎高生长随高程增加而减小[10].本试验结果发现1/4和2/4水位淹浸处理桐花树幼苗短时间(3~6个月),对茎生长有促进作用,随着淹浸时间的延长则起抑制作用;而中低水位处理有利于茎的生长.
红树植物的叶对淹水胁迫的反应敏感.Pezeshki等[20]发现,在表土渍水环境中的萌芽白骨壤、拉关木(Lagunculariaracemasa)和大红树植株的叶片生长均受到显著抑制,总叶面积减少. Ye等[21]对比研究发现,高水位处理的木榄幼苗总叶面积显著地小于低水位的.本研究表明,桐花树幼苗的叶对淹水胁迫的反应较茎和根更为强烈,高水位严重地抑制叶的生长,叶面积显著变小,叶片保存率降低.叶在各器官中受害程度最大,光合器官的损害和缺失降低幼苗生长速度和成活率.高水位的幼苗生物量显著地小于低水位的.
逆境下(如干旱、盐害和重金属胁迫等)植物根系活力下降[28].Ye等[22]研究表明红树植物受潮汐浸淹胁迫时根系活力显著降低.本研究结果与上述的研究相吻合,即高水位淡水生境下桐花树幼苗的根系活力显著下降,从而抑制桐花树幼苗根系的生长.可溶性糖是光合作用的重要产物,本研究发现水位增高会影响桐花树光合产物的积累,其原因是水位增高使植物的叶片数量和叶面积减少,造成光合能力下降.叶片光合产物积累减少,最终减缓植株生长.
桐花树幼苗叶片的SOD及POD酶活性在较高水位时显著升高,对桐花树叶片产生的一种生理响应.
红树植物长期适应于水淹生境,具有特殊的形态特征和生理响应等.但超过一定淹水深度和淹浸时间就会影响植株的生长发育,甚至枯死[23].试验还表明:2月龄桐花树幼苗在树体的2/4淹浸入淡水有较强的耐淹水能力.本研究结果可为桐花树在淡水河流、湖泊和水库的消落带中种植淹水位置的确定提供科学依据.
参考文献:
[1] 高桂娟,李志丹,韩瑞宏,等.桐花树研究进展[J].热带农业科学,2009,29(7):76-81.
Gao G J, Li Z D, Han R H, et al. Research progress of mangrove speciesAegicerascorniculatum[J].Chinese Journal of Tropical Agriculture, 2009, 29(7):76-81.
[2] 张忠华,胡刚,梁士楚.广西红树林资源与保护[J].海洋环境科学,2007,26(3):275-279,282.
Zhang Z H, Hu G, Liang S C. Mangrove resources and conservation in Guangxi[J]. Marine Environmental Science, 2007, 26(3):275-279,282.
[3] 杨盛昌,林鹏.红树植物秋茄和桐花树抗寒力的越冬变化[J].应用生态学报,1997,8(6):561-565.
Yang S C, Lin P. Cold-resistance ability of two mangrove speciesKandeliacandelandAegicerascorniculatumduring their overwintering period[J]. Chinese Journal of Applied Ecology, 1997,8(6):561-565.
[4] 王继栋,董美玲,张文,等.红树林植物桐花树的化学成分[J].中国天然药物,2006,4(4):275-277.
Wang J D, Dong M L, Zhang W, et al. Chemical constituents of mangrove plantAegicerascorniculatum[J].Chinese Journal of Natural Medicines,2006,4(4):275-277.
[5] 刘亚云,孙红斌,陈桂珠.氯联苯对桐花树幼苗生长及膜保护酶系统的影响[J].应用生态学报,2007,18(1):123-128.
Liu Y Y, Sun H B, Chen G Z. Effects of PCBs onAegicerascorniculatumseedlings growth and membrane protective enzyme system[J].Chinese Journal of Applied Ecology, 2007,18(1):123-128.
[6] 刁俊明,曾宪录,钟福生.不同浓度铅处理对桐花树叶片生理指标的影响[J].嘉应学院学报:自然科学版,2012,30(5):61-66.
Diao J M, Zeng X L,Zhong F S. The effect of lead exposure on the physiological characteristics ofAegicerasCorniculatumLeaf[J].Jouranl of Jiaying University:Natural Science,2012,30(5):61-66.
[7] 章金鸿,李玫,陈桂珠.红树林湿地对榨糖废水中N、P的吸收和净化的可能性[J].重庆环境科学, 1999,21(6):39-41.
Zhang J H, Li M, Chen G Z. The absorption and the potential capability in purificating N & P in sugar wastewater by mangrove wetland[J].China Chongqing Environment Science,1999,21(6):39-41.
[8] 何斌源,赖廷和,陈剑锋,等.两种红树植物白骨壤(Avicenniamarina)和桐花树(Aegicerascorniculatum)的耐淹性[J].生态学报,2007,27(3):1130-1138.
He B Y, Lai T H, Chen J F, et al. Studies of the tolerance ofAvicenniamarinaandAegicerascorniculatumto seawater immersion in Guangxi, China[J]. Acta Ecologica Sinica,2007,27(3):1130-1138.
[9] 罗美娟,张守攻,崔丽娟,等.桐花树幼苗生长与生物量分配对淹水胁迫的响应[J].浙江林业科技,2012,32(4):15-19.
Luo M J, Zhang S G,Cui L J, et al.Response of growth and biomass allocation ofAegicerascorniculatumto waterlogging stress[J]. Journal of Zhejiang Forestry Science and Technology, 2012,32(4):15-19.
[10] Kitaya Y,Jintana V,Piriyayotha S, et al. Early growth of seven mangrove species planted at different elevations in a Thai estuary[J].Trees-Structure and Function, 2002,16:150-154.
[11] 廖宝文,郑德璋,郑松发,等.红树植物桐花树育苗造林技术的研究[J].林业科学研究, 1998,11(5):474-480.
Liao B W, Zheng D Z, Zheng S F, et al. The studies on seedling nursery and afforestation techniques ofAegicerascorniculatumof mangroves[J].Forest Research,1998,11(5):474-480.
[12] 刁俊明,陈桂珠.盆栽桐花树对不同遮光度的生理生态响应[J].生态学杂志,2011,30(4):656-663.
Diao J M, Chen G Z.The eco-physiological responses of pot-plantedAegicerascorniculatumto different shade conditionss[J].Chinese Journal of Ecology,2011,30(4):656-663.
[13] 刁俊明,邹嫣娉,陈桂珠.淡水培养对桐花树幼苗生长发育的影响[J].林业科学研究, 2010,23(3):387-392.
Diao J M, Zou Y P, Chen G Z. Effect of fresh water cultivation on the growth and development ofAegicerascorniculatumseedlings[J].Forest Research,2010,23(3):387-392.
[14] 刁俊明,孙卿,陈桂珠.淡水驯化对桐花树光合生理特性的影响[J].植物研究,2010,30(3):416-423.
Diao J M, Sun Q, Chen G Z.The photosynthetic characteristics ofAegicerascomiculatumseedlings under fresh water adaptation[J].Bulletin of Botanical Research,2010,30(3):416-423.
[15] 张志良.植物生理实验指导[M].3版.北京:高等教育出版社,2003,41-165.
[16] 叶勇,卢昌义,郑逢中,等.模拟海平面上升对红树植物秋茄的影响[J].生态学报, 2004,24(10):2238-2244.
Ye Y, Lu C Y, Zheng F Z, et al. Effects of simulated sea level rise on the mangroveKandeliacandel[J]. Acta Ecologica Sinica,2004,24(10),2238-2244.
[17] Ellison A M,Farnsworth E J. Simulated sea level change alters anatomy, physiology, growth, and reproduction of red mangrove (RhizophoramangleL.)[J].Oceanographic Literature Review, 1998, 45(6):1003-1004.
[18] Ellison A M, Farnsworth E J. Seedling survivorship, growth,and response to disturbance inBelizeanmangal[J].American Journal of Botany, 1993,80:1137-1145.
[19] Farnsworth E J, Ellison A M. Sun/shade adaptability of the red mangrove,Rhizophoramangle(Rhizophoraceae): Changes through ontogeny at several levels of biological or-ganization[J].American Journal of Botany, 1996,83:1131-1143.
[20] Pezeshki S R, Delaune R D,Patrick W H Jr. Differential response of selected mangroves to soil flooding and salinity: Gas exchanges and biomass partitioning[J].Canadian Journal of Forest Research,1989,20:869-874.
[21] Ye Y, Tam N F Y,Wong Y S,et al. Does sea level rise influence propagule establishment, early growth and physic ology ofKandeliacandelandBruguieragymnorrhiza[J].Journal of Experimental Marine Biology and Ecology, 2004,306(2):197-215.
[22] Ye Y, Tam N F Y, Wong Y S,et al. Growth and physiological responses of two mangrove species (BruguieragymnorrhizaandKandeliacandel) to waterlogging[J]. Environmental and Experimental Botany, 2003,49:209-221.
[23] 赖廷和,何斌源.木榄幼苗对淹水胁迫的生长和生理反应[J].生态学杂志,2007,26(5):650-656.
Lai T H, HE B Y.Growth and physiological responses ofBruguieragymnorrhizaseedlings to waterlogging stress[J].Chinese Journal of Ecology,2007,26(5):650-656.
