蒙古沙冬青AM真菌物种多样性空间异质性
2014-08-15王晓乾贺学礼程春泉陈严严姜桥
王晓乾,贺学礼,程春泉,陈严严,姜桥
(河北大学 生命科学学院,河北 保定 071002)
AM(arbuscular mycorrhiza)真菌根外菌丝构成菌丝网络系统及其分泌的球囊霉素等代谢产物,将不同植物根系连接起来,这对保持荒漠植物多样性和稳定性,生态系统不同组分之间的生物演化与分布,能量、信息传递,物质交换等具有重要意义[1-3],在植物营养和水分状况改善,提高植物抗逆性和适应性方面也有重要作用.研究表明,荒漠环境AM真菌物种多样性丰富,不同宿主植物AM真菌群落组成和丰富度存在差异[4-6].因此,中国西部地区是受荒漠化危害较严重的地区,菌根生物技术研究的主要方向是通过AM菌剂-植物联合修复逆境.由于环境条件和土壤类型的复杂性和多样性,只有系统研究目标植物AM真菌在特定环境下物种多样性和群落组成,才能提高宿主植物适宜度,强化AM真菌应用效果,是植被在特殊条件下恢复获得成功的重要影响因素.
沙冬青属(Ammopiptanthus)仅含蒙古沙冬青(A.mongolicus)和新疆沙冬青(A.nanus),主要分布于中国西北荒漠地区,属于超旱生常绿阔叶灌木树种,是国家重点保护的珍稀濒危植物[7].蒙古沙冬青具有抗寒抗冻、耐高温、耐旱等特性,是荒漠地区用于防风固沙的优良灌木[8-9].近年来,有关沙冬青生理生态、迁地保护、区系分布与濒危原因等方面以及内蒙古地区AM真菌多样性研究[10]时有报道,人们从不同角度开展AM真菌物种多样性研究[11-12]、AM真菌与球囊霉素及环境影响因子的研究[13],对于荒漠环境AM真菌的研究多集中在菌根侵染及其结构类型等方面[14-15],涉及沙冬青AM真菌共生关系、生态分布规律和物种多样性的研究甚少.
本研究选取内蒙古不同荒漠样地,通过野外采样和室内分析,对蒙古沙冬青AM真菌群落组成和物种多样性变化规律进行研究,以便为筛选优良AM真菌菌种,促进荒漠植被恢复以及蒙古沙冬青生长提供依据.
1 材料与方法
1.1 样地概况
磴口县(N 40°29′,E 106°30′),海拔1 030 m,平坦沙地,年均降水量149.2 mm,年均蒸发量2 380.6 mm,主要伴生植物有骆驼刺(Alhagisparsifolia)和白刺(Nitrariatangutorum),沙冬青高大粗壮直立,节间大,叶子较乌拉特后旗的狭长.
乌拉特后旗(N 41°45′,E 107°157′),海拔1 295 m,平坦石质滩,年均降水量138.5 mm,年均蒸发量2 752.2 mm,主要伴生植物有沙蒿(Artemisiadesterorum)和驼驼蓬(Peganumharmala),沙冬青低矮,叶片浓密.
阿拉善左旗(N 41°45′,E 107°57′),海拔1 295 m,以平坦风蚀沙地为主,年均降水量112 mm,年均蒸发量3 217.7 mm,主要伴生植物有花棒(Hedysarumscoparium)和四合木(Tetraenamongolica),沙冬青低矮,冠幅大,叶片浓密.
乌海(N 39°45′,E 106°57′),海拔1 163 m,以洪水冲积沟滩为主,个别有大块石头堆积,土质松软,年均降水量159.8 mm,年均蒸发量3 289 mm,主要伴生植物有蒙古扁桃(Amygdalusmongolica)和白刺(N.tangutorum),沙冬青数量密集,叶片细长且稀疏.
1.2 样品采集
2013年7月,分别从所选样地随机选取生长良好的沙冬青植株各5株,去除其枯枝落叶层后,贴近植株根颈部挖土壤剖面,从0~50 cm土层采集土样,将土样装入塑料袋密封编号后带回实验室,土样阴干过2 mm筛4 ℃冷藏备用.
1.3 AM真菌分离鉴定
分离AM真菌孢子用湿筛倾析-蔗糖离心法[16],在体视显微镜下挑选大小相近、颜色相同的孢子置于乳酸、水等浮载剂中制片,观察孢子分类学特征,根据Schenck 等[17-18]的“VA 菌根鉴定手册”和国际AM真菌保藏中心(INVAM)在http://invam.caf.wvu.edu上提供的最新分类描述及图片等资料进行分类鉴定.
1.4 AM真菌多样性
孢子密度(spore density,SD):每20 g土样中含有的孢子数;种丰度 ( species richness,SR) :每20 g土样中含有AM 真菌种的数目,即 SR=AM 真菌总种次数 / 土样数;分离频度(frequency,F):AM 真菌某属或种出现次数/土样数×100%;相对多度(relative abundance,RA):该采样点AM 真菌某属或种的孢子数/该采样点AM真菌总孢子数×100%;重要值( importance value,I):频度和相对多度之和的平均值,即I=(F+RA) /2;Shannon多样性指数公式:H=-∑(Pi×lnPi);Simpson多样性指数公式:D=1-∑(Pi×Pi);均匀度指数:J=H/lnS,式中:Pi为某样地种i的孢子数(Ni)和该样地孢子总数(N)之比,即Pi=Ni/N,S为某样地AM真菌的种类总数.Sφrensen相似性系数公式:Cs= 2j/(a+b) ,j为AM真菌在A和B2个样地共有种数,a为A样地AM真菌全部物种数,b为B样地AM真菌全部物种数.
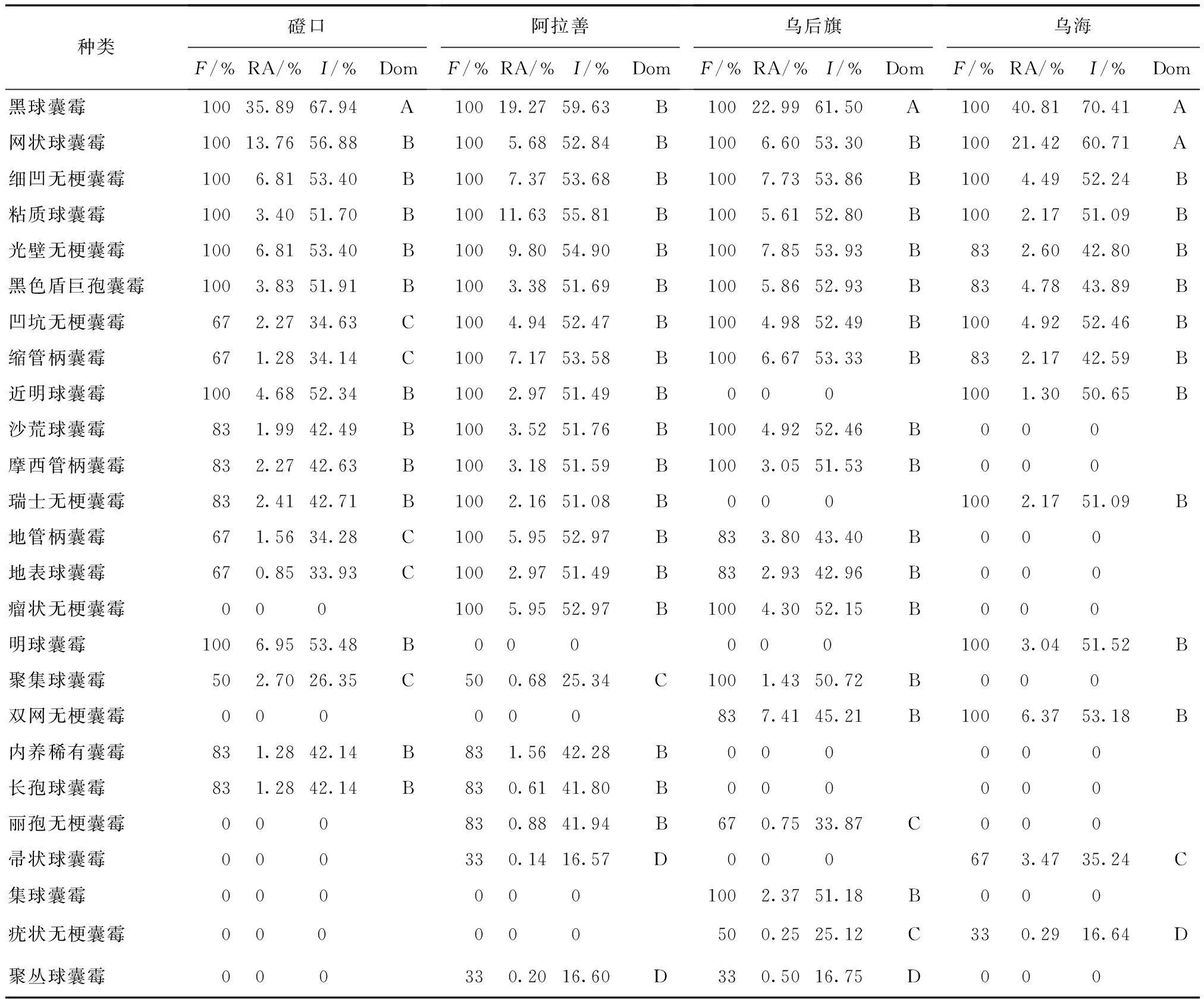
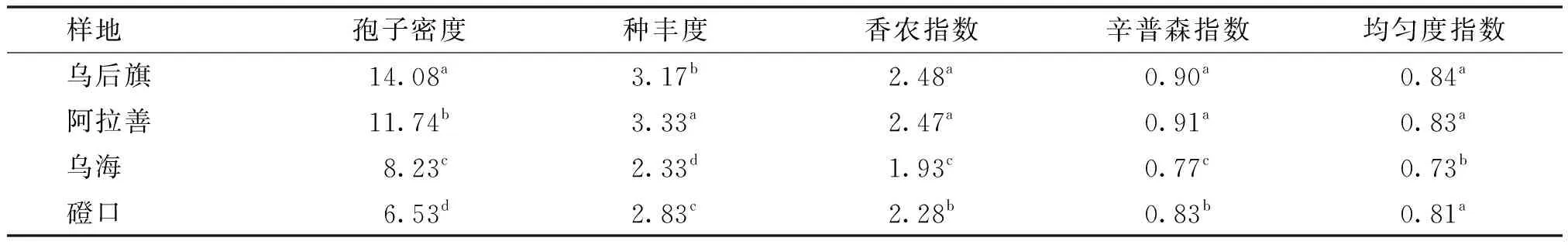
将 AM 真菌优势度按重要值(I) 划分为 4 个等级,即I>60% 为优势属(种) ,40% 采用SPSS19.0生物统计软件对实验数据进行单因素方差分析. 本实验共分离鉴定出AM真菌5属25种(图1),包括球囊霉属(Glomus)12种,无梗囊霉属 (Acaulospora)8种,管柄囊霉属(Funneliformis)3种,盾巨孢囊霉属(Scutellospora)和内养囊霉属(Entrophospora)各1种,其中黑球囊霉、粘质球囊霉、网状球囊霉、凹坑无梗囊霉、细凹无梗囊霉、光壁无梗囊霉和黑色盾巨孢囊霉等7种为4样地共有种. 磴口样地AM真菌有18种,阿拉善有21种,乌拉特后旗有19种,乌海有14种.球囊霉属和无梗囊霉属在4样地均有分布,盾巨孢囊霉属仅在阿拉善、乌拉特后旗和乌海出现,内养囊霉属只在阿拉善样地出现.不同属种AM真菌孢子形态特征差异明显,主要表现在孢子形态、颜色、大小、孢壁、表面纹饰和连孢菌丝等方面. a.地表球囊霉(Glomus versiforme);b.帚状球囊霉(G.coremioides);c.明球囊霉(G.clarum);d.沙荒球囊霉(G.deserticola);e.粘质球囊霉(G.viscosum);f.聚丛球囊霉(G.aggregatum);g.聚集球囊霉(G. glomerulatum);h.近明球囊霉(G.claroideum);i.长孢球囊霉(G.dolichosporum);j.集球囊霉(G.fasciculatum);k.黑球囊霉(G.melanosporum);l.网状球囊霉(G.reticulatum);m.丽孢无梗囊霉(Acaulospora elegans);n.光壁无梗囊霉(A.laevis);o.凹坑无梗囊霉(A.excavata);p.双网无梗囊霉(A.bireticulata);q.瑞士无梗囊霉(A.rehmii);r.疣状无梗囊霉(A.tuberculata);s.瘤状无梗囊霉(A.tuberculata);t.细凹无梗囊霉(A.scrobiculata);u.地管柄囊霉(Funneliformis geosporum );v.摩西管柄囊霉(F.mosseae);w.缩管柄囊霉(F.constrictum);x.黑色盾巨孢囊霉(Scutellospora nigra);y.稀有内养囊霉(Entrophospora infrequens).图1 5属25种AM真菌孢子形态 Fig.1 Spore morphology of 25 AM fungal species(×400) 由表1可知,球囊霉属和无梗囊霉属分离频度、相对多度、重要值在4样地较大,是共同优势属,内养囊霉属仅在磴口和阿拉善出现,与管柄囊霉属、盾巨孢囊霉属构成磴口和阿拉善的亚优势属,乌拉特后旗和乌海的亚优势属是管柄囊霉属和盾巨孢囊霉属. 表1 4样地AM真菌各属分离频度(F)、相对多度(RA)和重要值(I) 由表2可见,25种AM真菌中,黑球囊霉分离频度、相对多度和重要值最高,为4样地共同优势种.光壁无梗囊霉、细凹无梗囊霉、粘质球囊霉和黑色盾巨孢囊霉是4样地共同亚优势种.集球囊霉仅分布在乌拉特后旗,是亚优势种. 表2 4样地AM真菌种分离频度(F)、相对多度(RA)、重要值(I)和优势度(Dom) AM真菌分布具有明显空间差异,不仅样地AM真菌属种构成存在差异,而且阿拉善和乌拉特后旗AM真菌孢子密度、种丰度、香农指数和辛普森指数明显高于其他样地,均匀度指数乌拉特后旗≈阿拉善>磴口>乌海(表3). 表3 4样地AM真菌孢子密度、种丰度和物种多样性指标 同一列数据中不同上标字母表示在P<0.05水平上差异显著,孢子密度为20 g土样中含有的孢子数. 通过对2个样地间Sφrensen相似性系数计算得出,磴口和阿拉善相似性系数最大为0.87,阿拉善与乌拉特后旗相似性系数次之,为0.80,乌海与阿拉善和乌拉特后旗的相似性系数分别为0.63和0.55.统计分析表明,乌海与阿拉善和乌拉特后旗AM真菌组成差异显著. AM真菌与植物属于共同进化,其存在和活跃性受宿主和土壤类型显著影响[19].张淑容等[20]研究表明,蒙古沙冬青的须根以及荒漠环境下土壤透气性好,为AM真菌生长发育提供了侵染条件,使菌丝侵入到根组织细胞内,与蒙古沙冬青形成良好共生关系,而共生关系的形成有利于宿主植物生长和对极端荒漠环境的生态适应. 本实验共分离鉴定出5属25种AM真菌,多于沙坡头蒙古沙冬青根围的3属12种[11],与陈烝[21]等在塞北荒漠柠条锦鸡儿根围分离的4属24种AM真菌相接近,少于宁夏和甘肃荒漠地区蒙古沙冬青的4属36种[12].同时,球囊霉属和无梗囊霉属在4样地占比例最大.说明球囊霉属和无梗囊霉属是荒漠环境的广谱生态型[22]. 由于样地间土壤类型、年均降雨量及蒸发量等不同,使得不同样地蒙古沙冬青生长状态存在差异,同时影响AM真菌群落的种属组成、孢子密度、种丰度及多样性指数.内蒙古样地AM真菌孢子密度和种丰度与刘春卯[12]研究的宁夏、甘肃样地存在明显差异,磴口、乌海样地孢子密度和种丰度与宁夏相近,阿拉善与乌拉特后旗孢子密度和种丰度接近,甘肃孢子密度和种丰度明显高于其他样地.说明随着蒙古沙冬青生态地理变化,导致与之共生的AM真菌物种多样性发生了空间演替. 25种AM真菌中,有7种为4样地共有种.这7种共有种虽在不同样地均有分布,但7种AM真菌生态适应性不尽相同,主要体现在分离频度、相对多度、重要值、优势度,以及孢子颜色、表面纹饰等方面,共有种在不同样地间的差异,可能与土壤理化性质以及蒙古沙冬青生长状态有关. AM真菌属于专性共生真菌,适宜环境条件有利于AM真菌生长发育,生态环境(土壤类型、理化性质、植被类型和气候条件)差异对AM真菌多样性有重要影响[23].同时,AM真菌在长期进化过程中,也产生了相应的生态适应性.这对于进一步筛选优良AM真菌菌种,促进蒙古沙冬青生长和资源保护有重要意义. 参 考 文 献: [1] SMITH S E, READ D J. Mycorrhizal symbiosis[M].Cambridge, U K: Academic Press, 2008. [2] HE Xueli, LI Yingpeng, ZHAO Lili.Dynamics of arbuscular mycorrhizal fungi and glomalin in the rhizosphere ofArtemisiaordosicaKrasch.in Mu Us sandland,China[J]. Soil Biology and Biochemistry, 2010,42:1313-1319. [3] SANCHEZ-CASTRO I, NURIA F, PABLO C.Temporal dynamics of arbuscular mycorrhizal fungi colonizing roots of representative shrub species in a semi-arid Mediterranean ecosystem[J]. Mycorrhiza, 2012, 22(6):449-460. [4] 石兆勇,高双成,王发园. 荒漠生态系统中丛枝菌根真菌多样性[J].干旱区研究,2008,25(8):783-789. SHI Zhaoyong,GAO Shuangcheng,WANG Fayuan. Biodiversity of arbuscular mycorrhizal fungi in desert ecosystems[J].Arid Zone Research,2008,25(8):783-789. [5] 杨静,贺学礼,赵丽莉.内蒙古荒漠沙柳AM真菌物种多样性[J].生物多样性,2011,19(3):377-385. YANG Jing,HE Xueli,ZHAO Lili. Species diversity of arbuscular mycorrhizal fungi in the rhizosphere ofSalixpsammophilainInner Mongolia desert[J].Acta Ecologica Sinica,2011,19(3):377-385. [6] CHEN Zheng, HE Xueli, GUO Huijuan. Diversity of arbuscular mycorrhizal fungi in the rhizosphere of three host plants in the farming-pastoral zone, north China[J]. Symbiosis, 2012, 7(3): 149-160. [7] 王庆锁,李勇,张灵芝. 珍稀濒危植物沙冬青研究概况[J]. 生物多样性,1995,3(3):153-156. WANG Qingsuo,LI Yong,ZHANG Lingzhi.Rare and endangered plant research survey[J]. Biodiversity Science, 1995,3(3):153-156. [8] 蔚秋实,王继和,李昌龙.不同生境条件下沙冬青种群分布格局与特征的初步研究[J]. 植物生态学报,2005,29(4):591-598. WEI Qiushi,WANG Jihe,LI Changlong.A preliminary study on the distribution patterns and characeristics ofAmmopiptanthusmongolicuspopulations in different desert environments[J].Acta Phytoecologica Sinica, 2005,29(4):591-598. [9] 何恒斌,贾昆峰,贾桂霞. 沙冬青根瘤菌的抗逆性[J].植物生态学报,2006,30(1):140-146. HE Hengbin,JIA Kunfeng,JIA Guixia.A preliminary study on the stress resistance of rhizobia isolated fromAmmopiptanthusmongolicus[J].Journal of Plant Ecology, 2006,30(1):140-146. [10] 贺学礼,郭辉娟,王银银. 内蒙古农牧交错区沙蒿根围AM真菌物种多样性[J]. 河北大学学报:自然科学版,2012,32(5):506-514. HE Xueli,GUO Huijuan,WANG Yinyin.Species diversity of arbuscular mycorrhizal fungi in the rhizosphere ofArtemisiasphaerocephalain Inner Mongolia[J].Journal of Hebei University:Natural Science Edition, 2012,32(5):506-514. [11] 贺学礼,刘雪伟,李英鹏.沙坡头地区沙冬青AM真菌的时空分布[J].生态学报,2010,30(2):370-376. HE Xueli,LIU Xuewei,LI Yingpeng.The spatio-temporal distribution of arbuscular mycorrhizal fungi in the rhizosphere ofAmmopiptanthusmongolicusfrom Shapotou[J].Acta Ecologica Sinica, 2010,30(2):370-376. [12] 刘春卯,贺学礼,徐浩博. 蒙古沙冬青 AM 真菌物种多样性研究[J]. 生态环境学报,2013, 22(7): 1148-1152. LIU Chunmao,HE Xueli,XU Haobo. Species diversity of arbuscular mycorrhizal fungi in the rhizosphere ofAmmopiptanthusmonglicus[J].Ecology and Environmental Sciences,2013, 22(7): 1148-1152. [13] 贺学礼,陈程,何博. 北方两省农牧交错带沙棘根围AM真菌与球囊霉素空间分布[J]. 生态学报,2011, 31(6): 1653-1661. HE Xueli,CHEN Cheng,HE Bo. Spatial distribution of arbuscular mycorrhizal fungi and glomalin ofHippophaerhamnoidesL in farming-pastoral zone from the two northern provinces of China [J]. Acta Ecologica Sinica,2011, 31(6): 1653-1661. [14] 姜桥,贺学礼,陈伟燕.新疆沙冬青 AM 和 DSE 真菌的空间分布[J].生态学报,2014, 34(11):2929-2937 JIANG Qiao, HE Xueli,CHEN Weiyan.Spatial distribution of AM and DSE fungi in the rhizosphere ofAmmopiptanthusnanus[J].Acta Ecologica Sinica, 2014, 34(11):2929-2937. [15] 贺学礼,侯晓飞. 荒漠植物油蒿根围AM真菌的时空分布[J].植物生态学报,2008, 32(6): 1373-1377. HE Xueli,HOU Xiaofei. Spatio-temporal distribution of arbuscular mycorrhizal fungi fromArtemisiaOrdosicain Yulin city of Shaanxi Province,CHINA[J].Journal of Plant Ecology,2008, 32(6): 1373-1377. [16] IANSON D C, ALLEN M F.The effects of soil texture on extraction of vesicular arbuscular mycorrhizal spores from arid soils[J]. Mycologia, 1986,78:164-168. [17] SCHENCK N C, PEREZ Y. Manual for the identification of VA mycorrhizal fungi[M]. 2nd ed. Florida: FNVAM Gainesville, 1988: 91-97. [18] SCHÜβLER A, WALKER C. The glomeromycota: a species list with new families and genera[M]. Gloucester, England:Reatespace, 2010. [19] 山宝琴,贺学礼,白春明.荒漠油蒿根围AM真菌分布与土壤酶活性[J].生态学报,2009,29(6):3044-3051. SHAN Baoqin,HE Xueli,BAI Chunming.Distribution of arbuscular mycorrhizal fungi and soil enzyme activities in the rhizosphere ofArtemisiaordosicain desert[J]. Acta Ecologica Sinica, 2009,29(6):3044-3051. [20] 张淑容,贺学礼,徐浩博. 蒙古沙冬青根围AM和DSE真菌与土壤因子的相关性研究[J]. 西北植物学报,2013,33(9):1891-1897. ZHANG Shurong,HE Xueli,XU Haobo.Correlation study of AM and DSE fungi and soil factors in the rhizosphere ofAmmopiptanthusmongolicus[J].Acta Bot.Boreal.Occodent.Sin., 2013,33(9):1891-1897. [21] 贺学礼,陈烝,郭辉娟.荒漠柠条锦鸡儿AM真菌多样性[J].生态学报,2012,32(10) :3041-3049. HE Xueli,CHEN Zheng,GUO Huijuan.Diversity of arbuscular mycorrhizal fungi in the rhizosphere ofCaraganakorshinskiiKom.in desert zone[J].Acta Ecologica Sinica, 2012,32(10):3041-3049. [22] 杨秀丽,闫 伟,包玉英.大兴安岭落叶松林丛枝菌根真菌多样性[J]. 生态学报,2010,30(3): 504-510. YANG Xiuli,YAN Wei,BAO Yuying.Diversity of arbuscular mycorrhizal fungi in Dahurian larch forests in Da Hinggan Ling Mountains[J]. Acta Ecologica Sinica, 2010,30(3): 504-510. [23] SYKOROVA Z, WIEMKEN A, REDECKER D. CooccurringGentianavernaand Gentiana acaulis and their neighboring plants in two Swiss upper montane meadows harbor distinct arbuscular mycorrhizal fungal communities[J]. Applied Environmental Microbiology, 2007, 73(17): 5426-5434.1.5 数据处理
2 结果与分析
2.1 AM真菌群落组成

2.2 AM真菌属分离频度、相对多度和重要值

2.3 AM真菌种分离频度、相对多度和重要值
2.4 AM真菌物种多样性空间差异
2.5 AM真菌组成的相似性
3 讨论
