油菜田看麦娘对精喹禾灵的抗性水平及抗性机理研究
2014-08-10祝玮玮刘艺妩刘祥英柏连阳
祝玮玮, 刘艺妩, 刘祥英, 罗 坤, 柏连阳,2*
(1. 湖南农业大学农药研究所, 长沙 410128; 2. 湖南省农业科学院, 长沙 410128)
油菜田看麦娘对精喹禾灵的抗性水平及抗性机理研究
祝玮玮1, 刘艺妩1, 刘祥英1, 罗 坤1, 柏连阳1,2*
(1. 湖南农业大学农药研究所, 长沙 410128; 2. 湖南省农业科学院, 长沙 410128)
采用培养皿种子萌发法和盆栽法测定了湖南省境内长沙市、永州市、常德市、益阳市、浏阳市、岳阳市、娄底市7个市州10个地区油菜田看麦娘潜在抗药性种群对精喹禾灵的抗性水平,测定了常德市桃源地区看麦娘潜在抗药性种群以及敏感种群谷胱甘肽-S-转移酶(GSTs)对精喹禾灵的敏感性,对桃源地区看麦娘抗性品系及敏感品系ACCase基因片段进行扩增和测序,比较了两种生物型的基因序列。培养皿种子萌发法测定结果表明:常德市桃源地区看麦娘抗药性生物型对精喹禾灵的抗性水平最高,抗性倍数为10.50倍,其他地区看麦娘抗性倍数在2.01~7.09倍之间,抗性水平不明显;盆栽法测定结果表明:桃源地区看麦娘抗性倍数最高,为25.30倍,其他地区看麦娘抗性倍数在2.43~9.47倍之间,尚未产生明显抗药性。经精喹禾灵处理后,看麦娘抗药性生物型的GSTs活力在第5天明显高于敏感生物型,表明GSTs的活性是引起看麦娘对精喹禾灵抗性的重要因子。通过靶标基因片段扩增与DNA测序比对发现,抗药性生物型氨基酸序列第93位比敏感生物型多出一个丙氨酸,抗药性的产生与靶标基因的突变是否相关需要进行进一步验证。
油菜田; 看麦娘; 精喹禾灵; 抗药性
看麦娘(AlopecurusaequalisSobol.)为我国常见田间恶性杂草,属禾本科(Poaceae)禾亚科(Agrostidoideae)看麦娘属(Alopecurus),多危害小麦、油菜等夏收作物[1-2]。目前用于防治油菜田看麦娘的茎叶处理剂以乙酰辅酶A羧化酶ACCase抑制剂类为主,其主要包括两大类:芳氧苯氧丙酸类AOPP和环己烯酮类CHD。ACCase抑制剂类除草剂主要是抑制乙酰辅酶A羧化成丙二酰辅酶A,导致植物内部的脂肪酸无法合成,细胞膜结构遭到破坏,进而达到破坏植物代谢使其快速死亡的效果[3-4]。
由于ACCase抑制剂对双子叶植物的高度安全和对禾本科杂草的高效抑制,因此被广泛用于油菜田、小麦田等田间杂草的防除[5]。长期以来频繁和广泛使用除草剂使得油菜田杂草抗药性问题频发。2001年Heap就报道已有11种抗性生物型杂草在油菜田发现,其中最严重的是对ACCase抑制剂产生抗性的野燕麦和狗尾草[6]。我国最早对油菜田杂草看麦娘抗药性进行报道的是黄世霞,其通过培养皿种子萌发法研究发现部分油菜田看麦娘已对高效氟吡甲禾灵产生抗性[7]。王信群研究结果表明油菜田看麦娘已对高效氟吡甲禾灵产生一定抗性,同时对精喹禾灵产生交互抗性,但对环己烯酮类的快捕净无交互抗性[8]。
杂草对除草剂产生抗药性主要有三大原因[9-10]:一是作用靶标的改变;二是代谢作用的加强;三是除草作用的屏蔽。部分抗ACCase抑制剂类除草剂的杂草通过改变修饰体内的ACCase来达到降低对除草剂敏感性的效果,如Brown[11]等研究发现,ACCase的异亮氨酸突变为亮氨酸导致看麦娘对环己烯酮类除草剂烯禾啶产生抗性;Délye[12]等报道,ACCase在1 781位将异亮氨酸编码成亮氨酸的突变是看麦娘抗芳氧苯氧基丙酸类除草剂的主要原因。谷胱甘肽-S-转移酶(GSTs)是一个同工酶家族,其可受多种因素诱导,对一些有毒物质,包括农药、除草剂、致癌物和诱变剂等起到解毒作用[13],郇志博[14]研究黑龙江省大豆田稗草对精喹禾灵的抗性机理发现,低水平抗性与GSTs活动增强相关。
油菜田看麦娘对ACCase抑制剂产生抗性的问题已在部分地区发现并报道,但目前湖南省境内仍未见相关研究报道,鉴于此,笔者从湖南省内7个市10个地区的油菜田采集到了11个看麦娘潜在抗药性生物型,并从南京农业大学获得了相应的敏感生物型(未使用过除草剂),运用培养皿种子萌发法和盆栽法测定了其对精喹禾灵的抗性水平,测定了常德市桃源地区看麦娘潜在抗药性生物型以及由南京农业大学提供的看麦娘敏感生物型谷胱甘肽-S-转移酶的活性,并通过PCR扩增技术与DNA测序比对研究其抗药性产生的分子基础,为科学合理使用该药提供理论指导。
1 材料与方法
1.1 供试材料
药剂: 94%精喹禾灵原药,京博农化科技有限公司;15.8%精喹禾灵乳油,上海艾科思生物药业有限公司。
主要试剂:硝酸钾、无水乙醇、丙酮、吐温-80,天津恒兴化学试剂有限公司;PVP40,Sigma公司;Tris、L-还原型谷胱甘肽,Ameco公司;考马斯亮蓝、蛋白标准液,南京建成生物工程研究所;Plant Genomic DNA Kit(离心柱型),TIANGEN公司;10×EasyTaq buffer、dNTP mixture、EasyTaq DNAPloymerase,Trans公司;巯基乙醇、氯仿、Tris平衡酚,Solarbio公司。
主要仪器:智能型人工气候箱,宁波海曙赛福实验仪器厂;电子天平,赛多利斯科学仪器(北京)有限公司;WM-2天然气压缩机,天津仪器;Allegra X-22R 离心机,BECKMAN COULTER; 5804R离心机,德国Eppendorf公司;UV-1240紫外分光光度计,岛津公司;PCR扩增仪,Applied Biosystems公司;DYY-6 c型电泳仪,北京市六一仪器厂;凝胶成像分析系统,培清科技。
杂草种子:于2013年4-5月在湖南省7个市10个地区油菜田采集看麦娘潜在抗药性生物型,敏感种子由南京农业大学植物保护学院杂草研究室提供,采集地点与用药历史见表1。
表1看麦娘采集地点与精喹禾灵用药历史
Table1ThecollectingsitesofAlopecurusaequalisandthebackgroundofquizalofop-P-ethyl
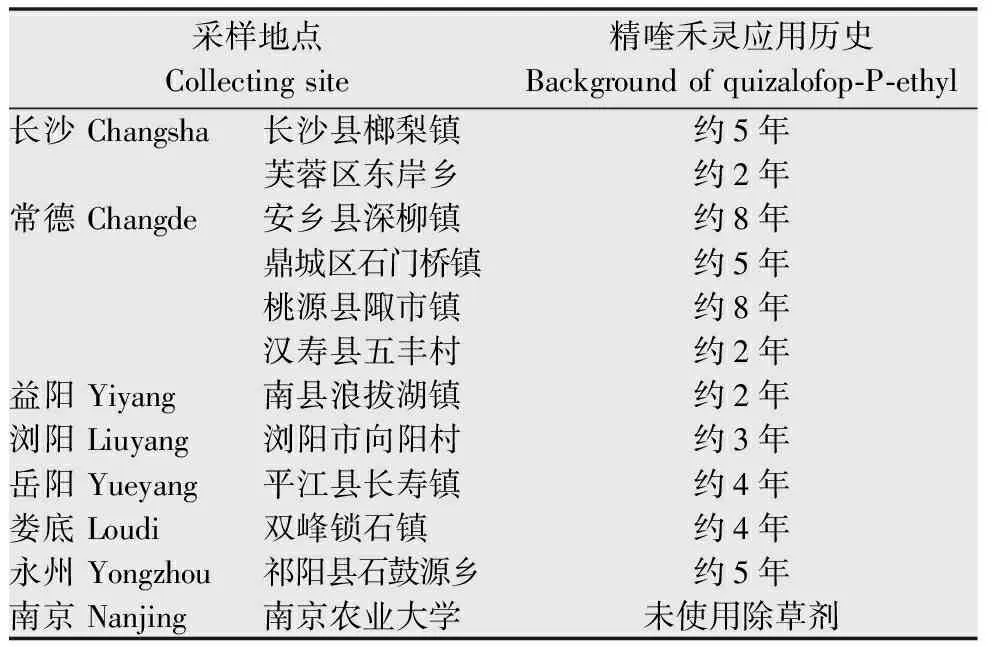
采样地点Collectingsite精喹禾灵应用历史Backgroundofquizalofop⁃P⁃ethyl长沙Changsha长沙县榔梨镇约5年芙蓉区东岸乡约2年常德Changde安乡县深柳镇约8年鼎城区石门桥镇约5年桃源县陬市镇约8年汉寿县五丰村约2年益阳Yiyang南县浪拔湖镇约2年浏阳Liuyang浏阳市向阳村约3年岳阳Yueyang平江县长寿镇约4年娄底Loudi双峰锁石镇约4年永州Yongzhou祁阳县石鼓源乡约5年南京Nanjing南京农业大学未使用除草剂
1.2 方法
1.2.1 看麦娘的抗药性测定
采用培养皿种子萌发法和盆栽法测定看麦娘对精喹禾灵的抗性水平[15]。培养皿种子萌发法:称取53.19 mg的94%精喹禾灵原粉,先用少量丙酮溶解后再用0.1%的吐温-80水溶液稀释成系列质量浓度药液,供试。
在直径9 cm的培养皿中进行剂量反应试验。看麦娘种子用质量分数为0.2%的硝酸钾(KNO3)于4 ℃冰箱中低温处理72 h后,继续用去离子水培养至露白,挑选长势一致的种子各40粒分别置于含有5 mL不同质量浓度药液和铺有两张滤纸的培养皿内,加盖确保种子处于湿润状态。精喹禾灵的有效成分含量分别为0.2、0.4、0.8、1.6、3.2、6.4 mg/L,以去离子水处理作为空白对照,每个处理重复4次。培养皿置于光照培养箱内培养,温度为白天24 ℃,晚上18 ℃,每天光照12 h。处理7 d后测量芽长,计算抑制率,求出回归方程。所有数据使用DPS软件处理。
盆栽法:称取0.03 mL的15.8%精喹禾灵乳油,用去离子水稀释成系列质量浓度药液,供试。
在直径为9 cm的塑料盆钵内,装入风干、过筛并与营养土混合后的土壤。每盆播种经0.2%硝酸钾处理72 h后培养至露白的看麦娘种子20粒。置于光照培养箱中培养,温度为白天24 ℃,晚上18 ℃,每天光照12 h。出苗后,每盆定苗10株,待看麦娘长至3~4叶期进行茎叶喷雾,喷液量为450 L/hm2。通过预试验设定精喹禾灵的有效处理剂量为236.88、118.44、59.22、29.61、14.81和7.40 g/hm2,以去离子水溶液处理作为空白对照,每个处理设4次重复。于喷药后21 d剪取看麦娘地上部分,称鲜重,计算各处理的鲜重抑制率,使用DPS软件求出回归方程。
1.2.2 谷胱甘肽-S-转移酶(GSTs)的活性测定[16]
谷胱甘肽-S-转移酶(GSTs)的提取:于施用精喹禾灵后1~8 d取样。取看麦娘地上部分0.5 g,剪碎放入预冷的研钵中,加入5 mL Tris-HCl缓冲液(0.1 mol/L,pH 8.0,含还原型谷胱甘肽25 mmol/L,5%PVP),冰浴匀浆,4 000 r/min离心10 min,取上清液,12 000 r/min离心5 min,取上清液作为酶提取液,以上操作均在-4 ℃条件下进行。
谷胱甘肽-S-转移酶(GSTs)的活性测定:于3 mL Tris-HCL缓冲液(0.1 mol/L,pH 8.0)中加入0.1 mL酶液,25 ℃保温10 min,加入0.1 mL无水乙醇配制的13 mmol/L CDNB,反应10 min 后,于 340 nm 处测得A值。以 1 mg/L 蛋白 10 min 催化反应使吸光值改变 0.001为一个酶活力单位,以1 mg/L可溶性蛋白的酶活力单位数为酶活力。以施药酶活力与空白对照酶活力的比值为相对活力。各处理重复3次,使用SPSS软件进行差异显著性分析。
1.2.3 ACCase的作用位点PCR扩增、DNA测序及序列分析
选取南京农业大学敏感品系和常德市桃源地区潜在抗性品系看麦娘作为供试材料,培养方法同盆栽法,出芽后每盆移种看麦娘20株,出苗后,每盆定苗16株,待各株看麦娘长至3~4叶期即使用Plant Genomic DNA Kit提取DNA。参考黄世霞[17]合成本试验引物(表2),用于扩增ACCase的CT区序列。
表2本试验中所用引物1)
Table2Theprimersusedinthisstudy

引物名称Primername引物序列Primersequence目的区域PositionACVRG15′CTGCAAACATTGGTG⁃GACCTCTTCCTATTAC3′5816~6872ACVRG1R5′CAGTCGGTGCTTCCTGC⁃TGCAGCTG3′
1) 目的区域指位于大穗看麦娘(Alopecurusmyosuroides)基因序列(GenBank No.AJ310767)中的位置。
PCR扩增体系包含:Template 2.4 μL,Primer ACVRG1、ACVRG1R各1 μL,10×EasyTaq Buffer 1 μL,dNTP Mixture和EasyTaq(5U/μL)各1 μL,Sterile Ultra-pure Water 36.6 μL。扩增程序如下:94 ℃ 10 min;94 ℃ 45 s,63 ℃ 50 s,72 ℃ 90 s,35个循环;72 ℃ 10 min。
PCR产物回收、测序均由博尚生物技术(上海)有限公司完成。测序结果由生物软件Vector NTI进行比对分析。同源性比对在基因数据库NCBI中进行BLAST分析。
2 结果与分析
2.1 看麦娘对精喹禾灵的抗性水平
通过培养皿种子萌发法测定看麦娘的抗药性水平(表3)结果表明,与敏感株系相比,常德桃源采集点的看麦娘对精喹禾灵的抗性水平最高,其抗性倍数为10.50倍,EC50为2.246 mg/L。其他采集点的抗性倍数在2.01~7.09之间,抗性水平不明显。
表3培养皿法测定不同看麦娘生物型对精喹禾灵的抗药性水平
Table3TheresistancelevelsofdifferentAlopecurusaequalisbiotypestoquizalofop-P-ethyldeterminedbyseedbioassayexperiments
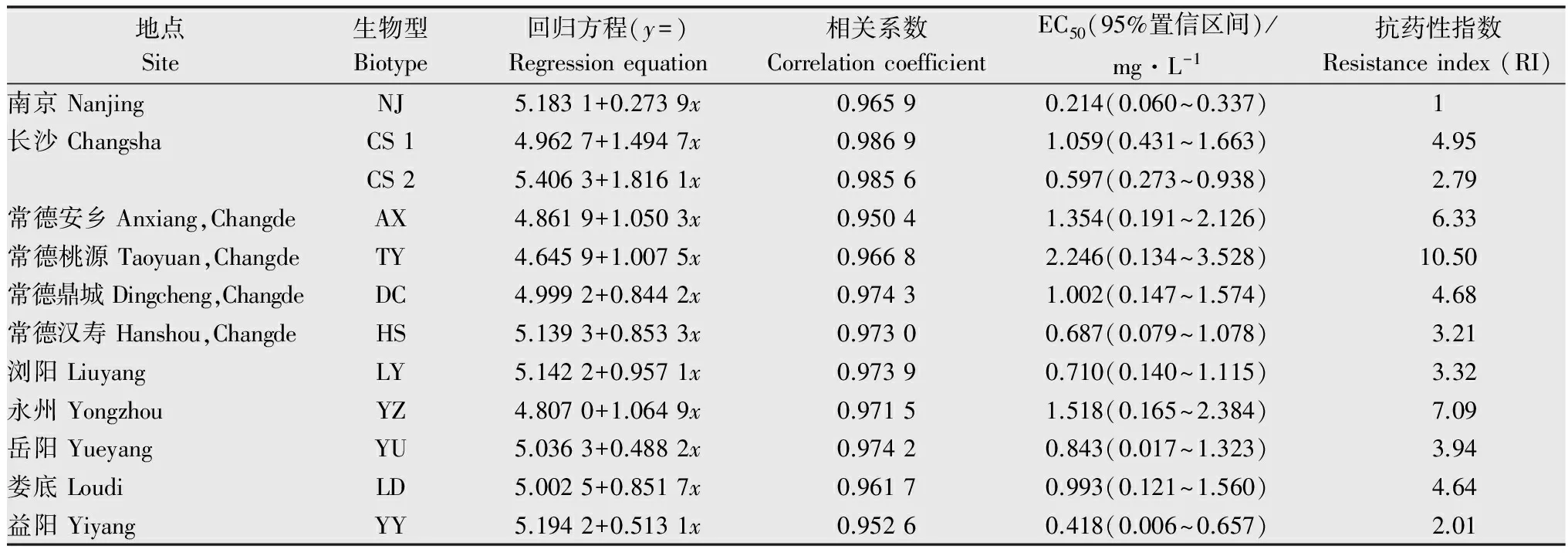
地点Site生物型Biotype回归方程(y=)Regressionequation相关系数CorrelationcoefficientEC50(95%置信区间)/mg·L-1抗药性指数Resistanceindex(RI)南京NanjingNJ5.1831+0.2739x0.96590.214(0.060~0.337)1长沙ChangshaCS14.9627+1.4947x0.98691.059(0.431~1.663)4.95CS25.4063+1.8161x0.98560.597(0.273~0.938) 2.79常德安乡Anxiang,ChangdeAX4.8619+1.0503x0.95041.354(0.191~2.126)6.33常德桃源Taoyuan,ChangdeTY4.6459+1.0075x0.96682.246(0.134~3.528)10.50 常德鼎城Dingcheng,ChangdeDC4.9992+0.8442x0.97431.002(0.147~1.574)4.68常德汉寿Hanshou,ChangdeHS5.1393+0.8533x0.97300.687(0.079~1.078)3.21浏阳LiuyangLY5.1422+0.9571x0.97390.710(0.140~1.115)3.32永州YongzhouYZ4.8070+1.0649x0.97151.518(0.165~2.384)7.09岳阳YueyangYU5.0363+0.4882x0.97420.843(0.017~1.323)3.94娄底LoudiLD5.0025+0.8517x0.96170.993(0.121~1.560)4.64益阳YiyangYY5.1942+0.5131x0.95260.418(0.006~0.657)2.01
盆栽法结果(表4)表明,常德桃源采集点的看麦娘对精喹禾灵的抗性水平较高,其抗性倍数为25.30倍,EC50为81.801 g/hm2,其他采集点的抗性倍数在2.43~9.47倍,尚未产生明显抗性。
表4整株法测定不同看麦娘生物型对精喹禾灵的抗药性水平
Table4TheresistancelevelsofdifferentAlopecurusaequalisbiotypestoquizalofop-P-ethylbywhole-plantbioassay
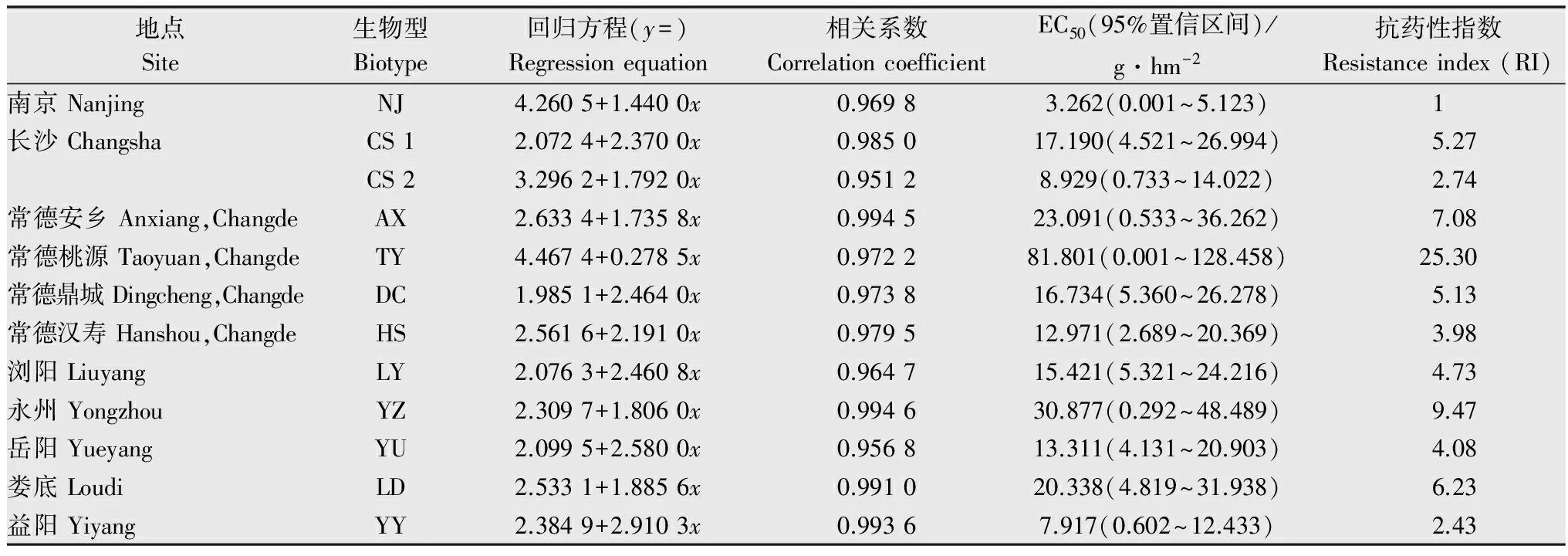
地点Site生物型Biotype回归方程(y=)Regressionequation相关系数CorrelationcoefficientEC50(95%置信区间)/g·hm-2抗药性指数Resistanceindex(RI)南京NanjingNJ4.2605+1.4400x0.96983.262(0.001~5.123)1长沙ChangshaCS12.0724+2.3700x0.985017.190(4.521~26.994)5.27CS23.2962+1.7920x0.95128.929(0.733~14.022)2.74常德安乡Anxiang,ChangdeAX2.6334+1.7358x0.994523.091(0.533~36.262)7.08常德桃源Taoyuan,ChangdeTY4.4674+0.2785x0.972281.801(0.001~128.458)25.30常德鼎城Dingcheng,ChangdeDC1.9851+2.4640x0.973816.734(5.360~26.278)5.13常德汉寿Hanshou,ChangdeHS2.5616+2.1910x0.979512.971(2.689~20.369)3.98浏阳LiuyangLY2.0763+2.4608x0.964715.421(5.321~24.216)4.73永州YongzhouYZ2.3097+1.8060x0.994630.877(0.292~48.489)9.47岳阳YueyangYU2.0995+2.5800x0.956813.311(4.131~20.903)4.08娄底LoudiLD2.5331+1.8856x0.991020.338(4.819~31.938)6.23益阳YiyangYY2.3849+2.9103x0.99367.917(0.602~12.433)2.43
2.2 精喹禾灵对抗性和敏感生物型看麦娘GSTs活力的影响
经精喹禾灵处理后,抗药性生物型看麦娘的GSTs活力即开始上升,至第5天达到最高值,为同期对照的1.27倍,随后GSTs活力开始下降,至第8天回落至同期对照水平;敏感生物型看麦娘GSTs活力在施药后开始下降,至第3天开始回升,第6天达到最高水平,为同期对照的1.12倍,第7天基本回落至对照水平,第8天降至对照水平以下。经方差分析发现,抗药性生物型GSTs活力在第5天显著高于敏感生物型GSTs活力,第1~4天及第6~8天两者并未发现显著性差异(图1)。
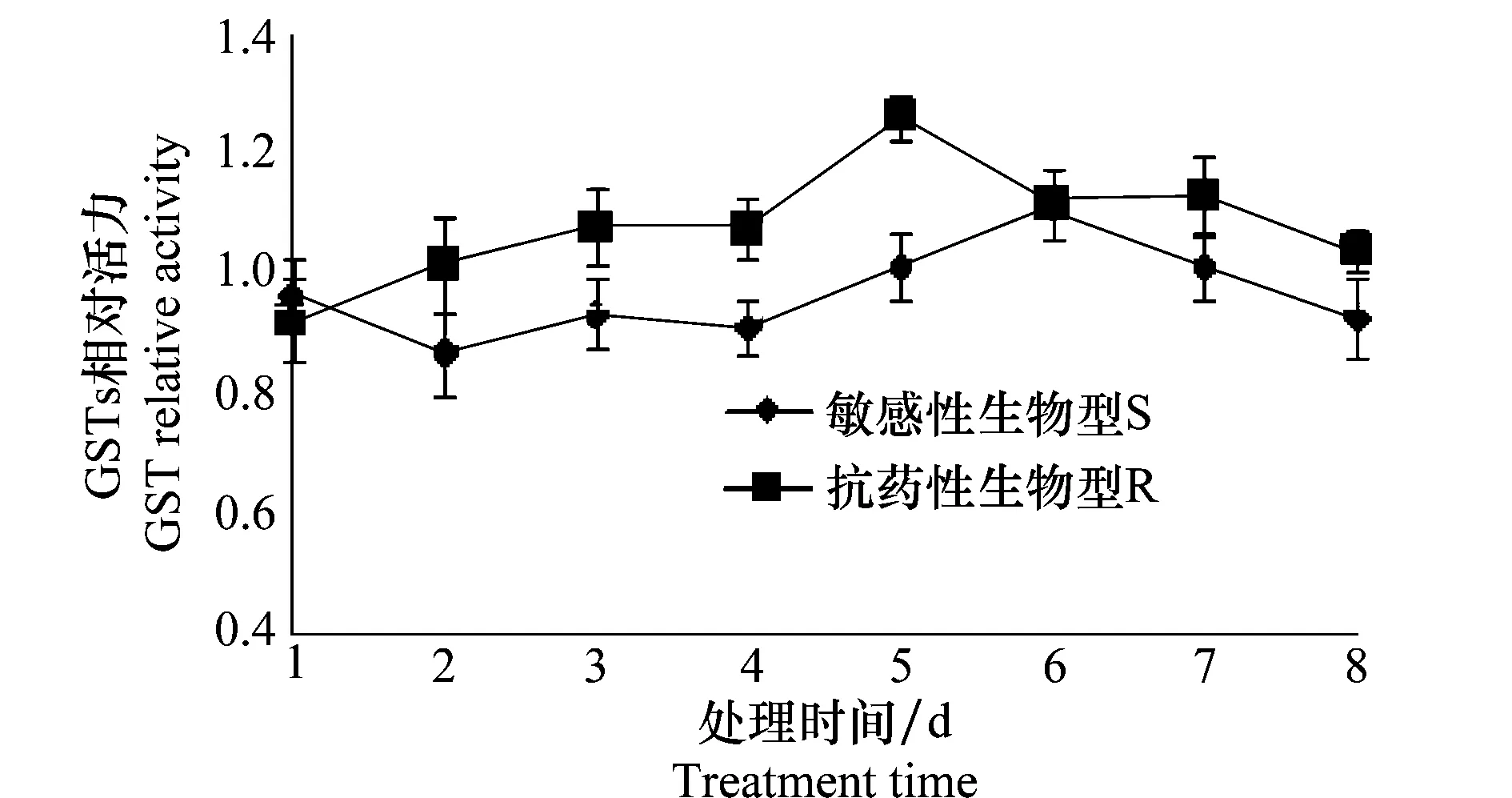
图1 精喹禾灵对看麦娘GSTs的影响
2.3 ACCase作用位点的序列分析
利用引物ACVRG1、ACVRG1R进行扩增,得到南农敏感生物型和桃源抗药性生物型看麦娘ACCase基因CT区的部分序列,扩增片段大小约为1 053 bp。在基因数据库NCBI中经BLAST分析比较,发现该序列与大穗看麦娘ACCase基因(GenBank No. AJ310767)序列同源性达90%,确定该扩增序列为看麦娘的ACCase基因部分序列。将扩增所得抗性及敏感生物型看麦娘氨基酸序列用Vector NTI11.5比对发现,抗药性生物型氨基酸序列在第93位比敏感生物型多一个丙氨酸(图2)。

图2 抗性及敏感生物型看麦娘靶标位点氨基酸序列比较(用引物ACVRG1、ACVRG1R扩增所得)
3 讨论
本研究采用培养皿种子萌发法和盆栽法测定了湖南省境内7个市10个地区采集点看麦娘对精喹禾灵的抗药性水平,两种测定方法结果均显示常德市桃源地区的抗性水平达到中等程度,其他地区采集点看麦娘对精喹禾灵尚未产生明显抗性,这一结果可能与桃源地区用药时间长于其他地区有关。明确各地油菜田看麦娘对精喹禾灵的抗药性水平,对制定化学防除方案及合理施用药剂有重要意义。此外,本研究中盆栽法抗性倍数普遍高于培养皿种子萌发法抗性倍数,其原因可能是培养皿法是在杂草幼苗期施药,而温室栽培法是在杂草3~5叶期施药,随着杂草自身的生长,其解毒代谢能力不断增强,故后者的抗性倍数要高于前者。
谷胱甘肽-S-转移酶(GSTs)是一类重要的解毒酶,它可以催化谷胱甘肽与多种疏水化合物的亲电子基团相连接,这种连接方式是生物体进行脱毒和排毒的重要方式[18]。研究结果显示,桃源地区看麦娘GSTs活力在第5天显著高于敏感品系看麦娘,表明其对精喹禾灵产生中等抗药性的原因之一为其代谢酶GSTs代谢能力的增强,这与其他相关禾本科杂草对ACCase抑制剂产生抗药性的研究结论基本一致[19-20]。Li Lingxu[21]等研究发现,第1 781位异亮氨酸突变为亮氨酸是导致麦田罔草对精恶唑禾草灵产生抗药性的主要原因;Tang Huaiwu[22]等报道,日本看麦娘对高效氟吡甲禾灵产生抗药性主要可能是由4个核苷酸突变导致的,包括甘氨酸取代1 734位精氨酸,丝氨酸取代1 739位苏氨酸,亮氨酸取代1 738位蛋氨酸,以及天冬酰胺取代2 041位异亮氨酸;汤怀武[23]报道,抗性和敏感生物型日本看麦娘ACCase基因序列存在11处差异。本研究结果显示,抗药性生物型植株的氨基酸序列在第93位比敏感生物型多一个丙氨酸,但考虑到抗药性生物型与敏感生物型来自不同生态区,遗传背景可能不同,因此其氨基酸序列的突变是由于其遗传背景不同导致还是抗性水平差异造成仍需进行进一步验证。
本试验从精喹禾灵对看麦娘GSTs活性影响和靶标位点基因的改变两个角度来阐述抗性产生的机理,但抗性的产生是否涉及其他酶系的作用以及相关靶标基因表达量的改变还有待进一步研究。
[1]孙丙耀. 麦田看麦娘属杂草种群消长原因及其抗性鉴定的研究[D]. 南京:南京农业大学,1996.
[2]黄世霞, 王庆亚, 张守栋. 看麦娘生物学特性研究[J]. 江苏农业科学,2006(4):58-60.
[3]Sasaki Y, Nagano Y. Plant acetyl-CoA carboxylase: structure, biosynthesis, regulation, and gene manipulation for plant breeding[J]. Bioscience, Biotechnology, and Biochemistry,2004,68(6):1175-1184.
[4]郭峰, 张朝贤, 黄红娟, 等. 杂草对ACCase抑制剂的抗性[J]. 杂草科学, 2011,29(3):1-6.
[5]黄世霞, 王庆亚, 董立尧, 等. 乙酰辅酶A羧化酶抑制剂类除草剂与杂草的抗药性[J]. 杂草科学, 2003(2): 1-5.
[6]Heap I, LeBaron H, Powles S B, et al. Introduction and overview of resistance[M]∥Powles S B, Shaner D L. Herbicide Resistance and World Grains. CRC Press, 2001: 1-22.
[7]黄世霞, 王庆亚, 张守栋. 油菜田看麦娘对高效盖草能产生抗药性的现状研究[J]. 安徽农业科学,2006,34(9):1913-1914.
[8]王信群, 黄世霞, 李楠. 油菜田看麦娘对10.8%高效盖草能抗性及对几种除草剂交互抗性的研究[J]. 安徽农业科学, 2006,34(16): 4022-4023.
[9]Heap I M. The occurrence of herbicide-resistant weeds worldwide[J]. Pesticide Science, 1997, 51(3): 235-243.
[10]张朝贤, 倪汉文, 魏守辉, 等. 杂草抗药性研究进展[J]. 中国农业科学, 2009,42(4):1274-1289.
[11]Brown A C, Moss S R, Wilson Z A, et al. An isoleucine to leucine substitution in the ACCase ofAlopecurusmyosuroides(black-grass) is associated with resistance to the herbicide sethoxydim[J]. Pesticide Biochemistry and Physiology,2002,72(3):160-168.
[12]Délye C, Wang T, Darmency H. An isoleucine-leucine substitution in chloroplastic acetyl-CoA carboxylase from green foxtail (SetariaviridisL. Beauv.) is responsible for resistance to the cyclohexanedione herbicide sethoxydim[J]. Planta,2002,214(3):421-427.
[13]郭玉莲, 陶波, 郑铁军, 等. 植物谷胱甘肽-S-转移酶 (GSTs) 及除草剂解毒剂的诱导作用[J]. 东北农业大学学报,2008,39(7):136-139.
[14]郇志博. 黑龙江省大豆田稗草对精喹禾灵的抗性研究[D]. 山东:山东农业大学,2011.
[15]陈宏州, 杨敬辉, 朱桂梅, 等. 油菜田日本看麦娘对高效氟吡甲禾灵的抗药性检测[J]. 西南农业学报, 2012,25(2):502-506.
[16]吴进才, 许俊峰, 冯绪猛, 等. 稻田常用农药对水稻3个品种生理生化的影响[J]. 中国农业科学, 2003,36(5):536-541.
[17]黄世霞, 何金铃, 王庆亚, 等. 看麦娘对稀禾啶和高效氟吡甲禾灵产生抗药性的分子基础[J]. 激光生物学报, 2010,19(6):832-837.
[18]Cobb A H, Kirkwood R C. Herbicides and their mechanisms of action[M]. England:Sheffield Academic Press,2000.
[19]韩瑞娟, 董立尧, 李俊, 等. 日本看麦娘对高效氟吡甲禾灵代谢抗性的初步研究[J]. 杂草科学,2010(1):3-7.
[20]Reade J P H, Milner L J, Cobb A H. A role for glutathione S-transferases in resistance to herbicides in grasses[J]. Weed Science,2004,52:468-474.
[21]Li Lingxu, Bi Yaling, Liu Weitang, et al. Molecular basis for resistance to fenoxaprop-p-ethyl in American sloughgrass (BeckmanniasyzigachneSteud.)[J]. Pesticide Biochemistry and Physiology, 2013,105(2):118-121.
[22]Tang Huaiwu, Li Jun, Dong Liyao, et al. Molecular bases for resistance to acetyl-coenzyme A carboxylase inhibitor in Japanese foxtail (Alopecurusjaponicus)[J]. Pest Management Science,2012,68(9):1241-1247.
[23]汤怀武. 日本看麦娘(Aloecurusjaponicus)对高效氟吡甲禾灵靶标酶与代谢酶抗性机理研究[D]. 南京:南京农业大学, 2012.
ResistancelevelofAlopecurusaequalistoquizalofop-P-ethylinrapeinHunanProvince
Zhu Weiwei1, Liu Yiwu1, Liu Xiangying1, Luo Kun1, Bai Lianyang1, 2
(1.InstituteofPesticides,HunanAgriculturalUniversity,Changsha410128,China;2.HunanAcademyofAgriculturalSciences,Changsha410128,China)
In order to investigate the resistance level and mechanism ofAlopecurusaequalisSobol. to quizalofop-P-ethyl in rape, 10 potential resistant biotypes were collected from 7 cities in Hunan Province and the susceptible biotype was obtained from Nanjing Agricultural University. The resistance level ofA.aequalisto quizalofop-P-ethyl was assessed by two methods: seed bioassay and whole-plant bioassay. The enzymology mechanism of Taoyuan Town potential resistant biotype and Nanjing Agricultural University susceptible biotype was studied with the important metabolic enzyme glutathione-s-transferase(GSTs), and the resistance mechanism at the molecular level was detected by using PCR technology and comparing the amino acid sequences of resistant and sensitive plants. The results of seed bioassay showed that the relative resistance factor of Taoyuan Town in Changde was 10.50, which was the highest. The resistance factor of all of the other potential resistant biotypes ranged from 2.01 to 7.09, which was sensitive to quizalofop-P-ethyl. The results of whole-plant bioassay showed that the relative resistance factor of the biotype from Taoyuan Town in Changde was 25.30, while the factors of other potential resistant biotypes were from 2.43 to 9.47. The GST activity of resistant biotypes was obviously higher than that of the susceptible biotype on the fifth day, indicating that the difference in the metabolic ability of GSTs to quizalofop-P-ethyl was an important reason for the resistance to quizalofop-P-ethyl. The comparison of ACCase sequences between the resistant and sensitive biotypes showed that the amino acid sequence of resistant biotype had one additional Ala, but further verification is needed to figure out the relationships between resistance level and target gene’s mutation.
rape field;Alopecurusaequalis; quizalofop-P-ethyl; herbicide resistance
2014-01-08
:2014-03-25
公益性行业(农业)科研专项(201303031)
S 481.4
:ADOI:10.3969/j.issn.0529-1542.2014.05.014
* 通信作者 E-mail: bailianyang2005@aliyun.com
