福寿螺对稻田水生植物群落结构的影响
2014-08-08赵本良章家恩戴晓燕黄伟均
赵本良,章家恩,*,戴晓燕,彭 莉,彭 卉,黄伟均
(1. 华南农业大学农学院生态学系, 广州 510642; 2. 农业部华南热带农业环境重点开放实验室, 广州 510642;3. 广东省高等学校农业生态与农村环境重点实验室, 广州 510642)
福寿螺对稻田水生植物群落结构的影响
赵本良1,2,3,章家恩1,2,3,*,戴晓燕1,彭 莉1,彭 卉1,黄伟均1
(1. 华南农业大学农学院生态学系, 广州 510642; 2. 农业部华南热带农业环境重点开放实验室, 广州 510642;3. 广东省高等学校农业生态与农村环境重点实验室, 广州 510642)
福寿螺是在我国南方地区危害水稻生产的一种外来入侵软体动物,已被世界自然保护联盟入侵生物专家组列为100种危害最为严重的外来入侵物种之一。在水域生态系统中福寿螺主要以摄食各种水生植物而生存。以水稻、大薸、水葫芦、粉绿狐尾藻、水花生和鸭舌草构成的稻田水生植物群落为对象,研究了低密度(4 只/m2)、中密度(8 只/m2)和高密度(12 只/m2)福寿螺处理对稻田水生植物群落的影响,同时结合福寿螺的食物选择性实验分析其相关行为特性。结果表明:1)低密度、中密度和高密度福寿螺处理1周内水稻密度均急剧下降,高密度福寿螺造成水稻密度下降超过70%,2周后福寿螺危害水稻秧苗的程度减轻;2)各密度处理福寿螺均抑制了水花生和鸭舌草扩散,第8周时高密度福寿螺处理中水花生和鸭舌草的密度仅为对照的30%和25%。同时,低密度福寿螺处理中的水花生和鸭舌草的密度显著高于中密度和高密度处理。不同密度福寿螺处理对水葫芦、大薸和粉绿狐尾藻影响不明显;3)水稻的日均密度增长率为负值,低密度、中密度和高密度福寿螺处理间存在明显差异,且均显著低于对照。水花生和鸭舌草的日均密度增长率虽为正值,但显著低于对照,水葫芦、大薸和粉绿狐尾藻的日均密度增长率和对照无差异。福寿螺日均密度增长迅速,各密度处理无显著差异。福寿螺对水稻、水花生、鸭舌草的选择性显著高于水葫芦、粉绿狐尾藻和大薸;4) 福寿螺引入8周后稻田水生植物群落物种多样性Simpson指数、Shannon指数和均匀度指数均显著降低。Simpson指数的大小顺序为:对照>低密度>中密度、高密度,Shannon指数和均匀度指数的大小顺序为:对照>低密度>中密度>高密度。说明福寿螺对供试水生植物有明显的选择性,其入侵会严重危害稻田水生植物群落的物种多样性。
福寿螺;入侵;水生植物;生物多样性;摄食偏好
外来物种入侵是威胁入侵生境生物多样性的一个重要因素[1]。由于各种生境的物种数量和组成比例存在较大差异,外来物种的入侵过程往往发生在具有不同“背景”生物多样性的生境中。实际上,入侵地“背景”生物多样性和外来生物入侵造成的生物多样性变化效应之间存在密切联系。具体来说,在入侵地处于低水平生物多样性的情况下,外来生物入侵可能会造成入侵地物种多样性的多向变化,如仅压制优势物种的增长,而对其他物种影响不明显,导致生物多样性出现升高或维持原态不变,或对所有物种均有一定抑制效应,生物多样性进一步降低等,因此,入侵生境的“背景”生物多样性可能是影响外来生物影响入侵地生物多样性的一个重要因素。
福寿螺(Pomaceacanaliculata(Lamarck))原产于南美洲,属中腹足目(Mesogastropoda)、瓶螺科(Ampullariidae)、瓶螺属(Pomacea)[2]。20 世纪80年代初作为食品引入我国,后因管理不善逃逸至各类水体,目前已经广泛分布于我国南方地区。福寿螺是杂食性动物,摄食对象广泛,包括大型水生植物、浮游藻类、无机及有机碎屑等[3],其中以摄食水生植物为主,目前报道福寿螺可以摄食的水生植物种类不低于50种[4- 6],据2008年不完全统计,我国广东16.7万 hm2左右的水稻遭受福寿螺不同程度的危害,且福寿螺在广西、海南、福建、湖南等省份也已有大面积的分布[7]。福寿螺已被列为国家环保部首批公布的16种重要外来入侵物种“黑名单”,也是世界100种恶性入侵生物之一[8- 9]。福寿螺除了严重危害水稻以外,还对水生生态系统造成一定风险,其入侵自然湿地后改变了自然湿地水生植物群落的结构,削弱了自然湿地的生态系统服务功能[10- 11]。福寿螺以摄食水生植物为主,入侵后影响稻田水生植物群落的组成和结构,而稻田生态系统的水生植物群落以水稻为主,其水生植物多样性在稻田重建阶段往往低于自然湿地。因此,分析福寿螺入侵稻田后对水生植物多样性的影响过程将有助于进一步理解生物入侵后入侵生境低水平生物多样性“背景”下生物多样性的变化过程。然而,目前亦未见福寿螺影响稻田水生植物群落物种多样性方面的研究报道。因此,本研究试图在特定植物组成的稻田水生植物群落基础上,以入侵生物福寿螺对稻田水生植物多样性的影响为研究实例,结合其摄食特性差异,深入分析入侵生物对稻田水生植物的影响,尤其是稻田水生植物群落物种多样性的变化规律,旨在为入侵生物的生态风险评价提供参考。
1 材料与方法
1.1 供试福寿螺和水生植物
福寿螺采集于华南农业大学农场(23°14′N, 113°38′E),放置于室内饲养1周,实验前饥饿24 h。根据文献分类方法[12],选择活动活跃的幼螺(重量0—6.5 g)和成螺(重量6.5—12 g)作为供试材料。幼螺为未成年螺,性特征不明显,未设置雌雄比;成年福寿螺的雌雄比设为1∶1。稻田水生植物品种为水稻(OryzasativaL. cv. Huanghuazhan)、大薸(PistiastratiotesL.)、水葫芦(Eichhorniacrassipes)、粉绿狐尾藻(Myriophyllumaquaticum)、水花生(Alternantheraphiloxeroides)和鸭舌草(Monochoriavaginalis),其中水稻秧龄3周,其他水生植物每株根长3—8 cm,鲜重2—5 g。
1.2 实验设计及采样方法
实验共计12个小区,小区面积4.5 m2(1.5 m×3 m),分为3个不同密度福寿螺处理和1个对照,均重复3次(图1),各小区间采用塑料薄膜隔开,以防止福寿螺混杂。参考相关福寿螺密度的研究报道[13],分别设置为低密度、中密度和高密度福寿螺处理,其中低密度处理投放幼螺10只和成螺8只,福寿螺密度为4 只/m2;中密度处理投放幼螺20只和成螺16只,福寿螺密度为8 只/m2;高密度处理投放幼螺30只和成螺24只,福寿螺密度12 只/m2。水稻每小区125株,密度28 株/m2。其他水生植物每小区4株,密度约1 株/m2。具体操作方法为:先将秧苗移栽至小区,而后标记小区,将其等分为50 cm×50 cm的假想方格。而后随机选择4个方格,供试的水生植物栽植于方格中水稻株行对角线正中(9 cm×10 cm),重复操作直至移栽完每种植物(图2)。实验时分别于第0、1、2、4和8周调查,并记录各小区内各种水生植物数量;于第8周时采用50 cm×50 cm的采样框摸查福寿螺的数量,统计后获得各小区内福寿螺的总数。
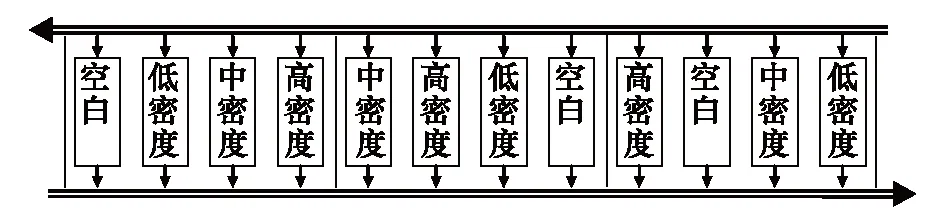
图1 小区田间排列示意图Fig.1 Field plots arrangement in experiment
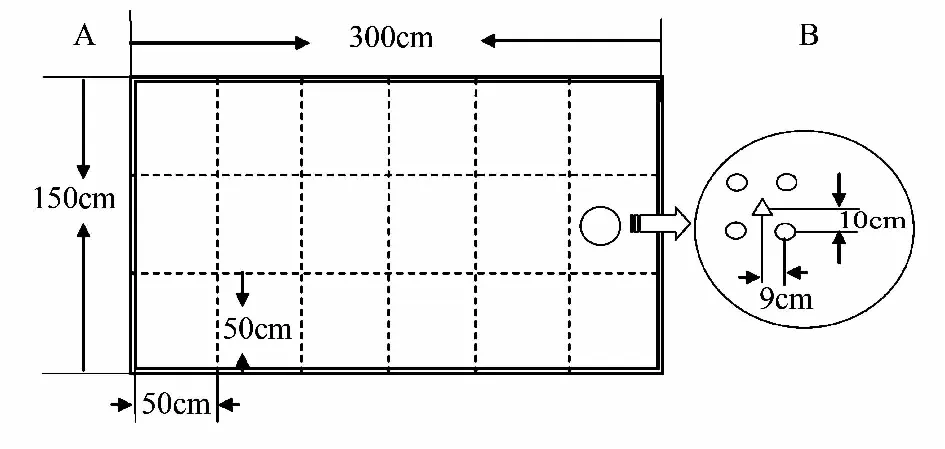
图2 稻田水生植物的种植位置示意图Fig.2 Planting location of aquatic macrophytes in field plotsA为小区布置,B为水生植物位置的局部放大,○为水稻,△为大薸、水葫芦、粉绿狐尾藻、水花生和鸭舌草的一种A: whole schema of one field plot; B: partial enlarged planting location of aquatic macrophytes; ○ represents rice, △ represents one kind of macrophyte among Pistia stratiotes L. ,Eichhornia crassipes, Myriophyllum Aquaticum, Alternanthera philoxeroides and Monochoria vaginalis
1.3 多样性指数和日均密度增长率的计算
实验中采用Simpson指数(D)、Shannon指数(H)和Pielou均匀度指数(E)分析福寿螺引入后稻田水生植物群落物种多样性的变化[14]:
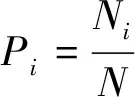
日均密度增长率计算公式为:
式中,Rf为第8周时的水生植物(株/m2)或福寿螺(只/m2)的密度,R0为初始水生植物(株/m2)和福寿螺(只/m2)密度,t为8周(56 d)。
1.4 福寿螺摄食偏好实验
摄食偏好实验根据文献改进[15- 16],采用分成6个方位的塑料盆于室内(25±1)℃进行,实验时每次取16只活跃福寿螺,幼螺8只,成螺8只(其雌雄比例为1∶1),置于塑料盆中0号位置,其他方位(1—6)的分别放置水稻、大薸、水葫芦、粉绿狐尾藻、水花生和鸭舌草等水生植物,重复6次(图3),1 h后观察并记录福寿螺达到的各个位置。
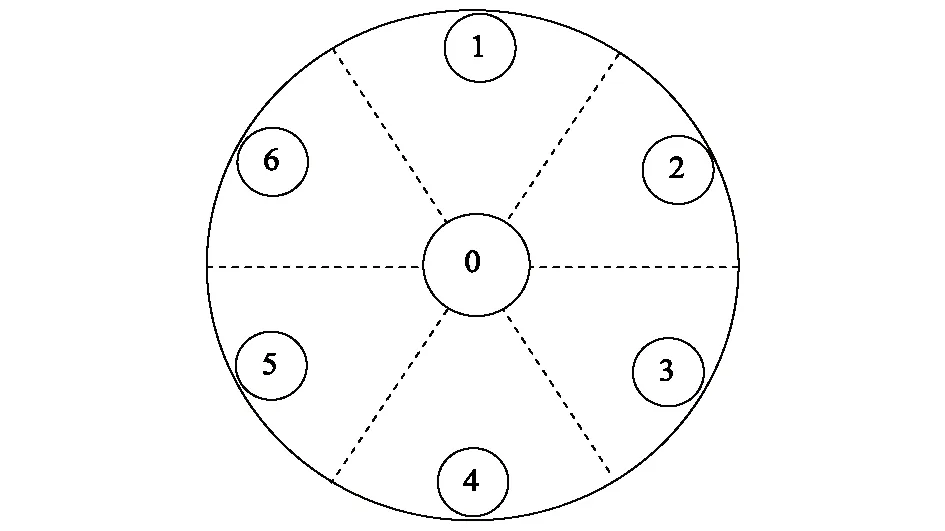
图3 摄食偏好实验中水生植物和福寿螺的位置布局示意图Fig.3 The layout of macrophytes and Pomacea canaliculata in the feeding preference experiment
1.5 数据统计分析
所有数据均用Microsoft Office Excel 2003和Origin Pro 7.5 整理绘图,数据经SPSS 13.0计算后以平均值±标准误的形式表示,差异显著性测验采用SPSS 软件的邓肯新复极差法。
2 结果与分析
2.1 福寿螺对稻田不同水生植物密度的影响
水稻受害状况随着福寿螺密度的增加而加重(图4)。和对照相比,1周时各密度福寿螺处理中的水稻密度下降均超过了40%,且高密度福寿螺处理中水稻密度下降超过70%。第2周时低密度和中密度福寿螺处理下水稻密度仍有约2%的轻微下降,高密度处理中水稻密度则下降了约18%。第2周至第8周,各处理的水稻密度保持不变。这表明福寿螺摄食水稻会持续一定时间,能够在移栽1—2周内对水稻造成严重危害。
福寿螺引入造成鸭舌草的生长繁殖受到明显抑制(图4)。第1周时,福寿螺处理组的鸭舌草密度显著低于对照(P<0.01)。第2周,高密度的福寿螺处理的鸭舌草密度显著低于中、低密度,亦显著低于对照(P<0.01),而中、低密度福寿螺处理间的鸭舌草密度差异不显著。第4周,福寿螺处理的鸭舌草密度和对照差距进一步扩大,而3个密度福寿螺处理间的鸭舌草密度接近。第8周时,鸭舌草密度大小为:中、高密度福寿螺处理<低密度处理<对照。
福寿螺引入对水花生密度增长具有较大的负效应(图4)。第1周,低、中和高密度福寿螺处理的水花生密度均大幅低于对照(P<0.01)。第2周,福寿螺处理组的水花生密度均显著低于对照,且福寿螺各密度处理间水花生密度差异不显著。第4周,中、高密度福寿螺处理组的水花生密度呈现不同程度的下降,第8周时水花生密度在福寿螺处理组和对照间的差异达到极显著水平(P<0.01),低密度福寿螺处理下的水花生密度亦显著高于中密度和高密度处理。
福寿螺引入对水葫芦密度影响不明显(图4)。第2周前,水葫芦的密度上升迅速,第2周至第8周,低密度福寿螺处理和对照的水葫芦密度逐步稳定,而中密度和高密度福寿螺处理仍在上升,至第8周时各处理和对照的水葫芦密度均没有显著差异。
福寿螺引入对大薸生长繁殖无明显影响(图4)。实验中各福寿螺处理和对照的大薸密度增加十分迅速。第1周时,福寿螺处理和对照的大薸密度十分接近;第2周后,低密度福寿螺处理中的大薸密度呈稳定状态,而在中密度和高密度福寿螺处理中仍在缓慢上升。至第8周时,福寿螺处理和对照的大薸密度无显著差异。
福寿螺引入并未影响粉绿狐尾藻生长密度的变化(图4),各福寿螺密度处理和对照的粉绿狐尾藻密度一直处于上升状态。第1周时,各处理和对照的粉绿狐尾藻密度差异不明显;第2周时,中密度福寿螺处理的粉绿狐尾藻密度高于对照,其他无显著差异。第4周和第8周时,福寿螺处理和对照间的粉绿狐尾藻密度差异不显著。

图4 福寿螺引入后稻田不同水生植物密度的变化Fig.4 Density change of five aquatic macrophytes after Pomacea canaliculata was introduced into the paddy fields
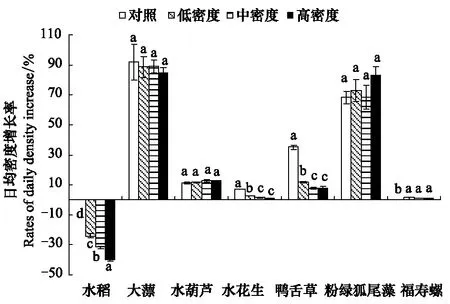
图5 福寿螺和水生植物的日均密度增长率变化Fig.5 Change of the rates of daily density increase of Pomacea canaliculata and macrophytes
2.2 福寿螺和水生植物日均密度增长率的变化
不同福寿螺密度处理中的福寿螺和6种水生植物的日均密度增长率变化存在差异(图5)。各密度处理中福寿螺的密度上升迅速,各处理间较接近。福寿螺造成水稻秧苗的日均密度增长率为负值,表明水稻生长受到严重危害,且随福寿螺密度升高,危害效应增强,高密度福寿螺处理危害最大(P<0.01)。水花生和鸭舌草的日均密度增长率虽为正值,但相比对照,其日均密度增长率显著下降,且随着福寿螺密度上升,下降趋势更大(P<0.01),表明两种植物的生长均受到福寿螺的抑制。大薸、粉绿狐尾藻和水葫芦的日均密度增长率在各处理和对照间无显著差异,表明福寿螺引入并不影响其生长。
2.3 福寿螺对稻田水生植物多样性的影响
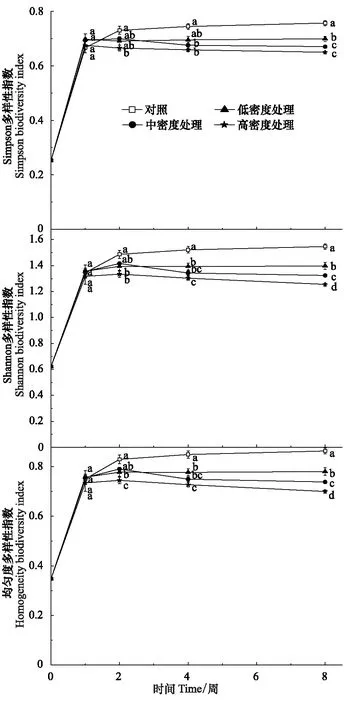
图6 福寿螺引入后水生植物Simpson、Shannon多样性指数和均匀度的变化Fig.6 Change of Simpson, Shannon and Homogeneity indices of aquatic macrophyte community after Pomacea canaliculata was introduced into the paddy fields
福寿螺引入1周后稻田水生植物的Simpson指数呈现缓慢下降趋势(图6)。第1周时,不同密度福寿螺处理和对照之间无显著性差异;第2周时,Simpson多样性指数分化,高密度福寿螺处理的Simpson指数显著低于对照(P<0.05)。第4周时,不同密度福寿螺处理的Simpson指数继续下降,中、高密度处理已显著低于对照(P<0.01)。第8周,Simpson指数的大小顺序为:对照>低密度>中密度、高密度,对照和福寿螺处理的Simpson指数达到极显著差异(P<0.01)。低密度福寿螺引入即可造成稻田水生植物群落的Simpson指数下降,中高密度福寿螺处理对Simpson指数的负面影响更甚,高密度福寿螺处理比对照下降了约14%,整体上不同密度福寿螺处理后稻田水生植物群落Simpson多样性指数明显降低,说明福寿螺引入会严重危害稻田水生植物群落的物种多样性。
除了对照以外,不同福寿螺密度处理的Shannon指数呈现先上升后下降的趋势(图6)。第1周时,各福寿螺处理和对照的Shannon指数无显著差异。第2周至第8周,不同密度处理的Shannon指数均在缓慢下降,第4周不同密度福寿螺处理均显著低于对照(P<0.01)。至第8周,高密度福寿螺处理的Shannon指数已经低于对照19%,且不同密度福寿螺处理间也存在显著差异(P<0.01),随着福寿螺密度增大,Shannon多样性指数降低愈大。福寿螺引入对稻田水生植物群落物种多样性有明显的负面效应。
与对照相比,不同密度福寿螺处理的水生植物群落的均匀度指数的变化趋势是先上升后逐渐下降(图6)。第1周时,各密度福寿螺处理间、且其和对照间,均匀度指数均无显著差异。第2周和第4周,不同密度福寿螺处理的均匀度指数下降,逐渐低于对照;至第8周,各密度福寿螺处理之间的均匀度指数差异极显著(P<0.01),其顺序为:低密度>中密度>高密度,三者均极显著低于对照(P<0.01),其中高密度处理的均匀度指数相比对照降低了约19%,表明不同密度福寿螺引入会造成稻田水生植物群落均匀度指数降低。
2.4 福寿螺对稻田水生植物的摄食偏好分析
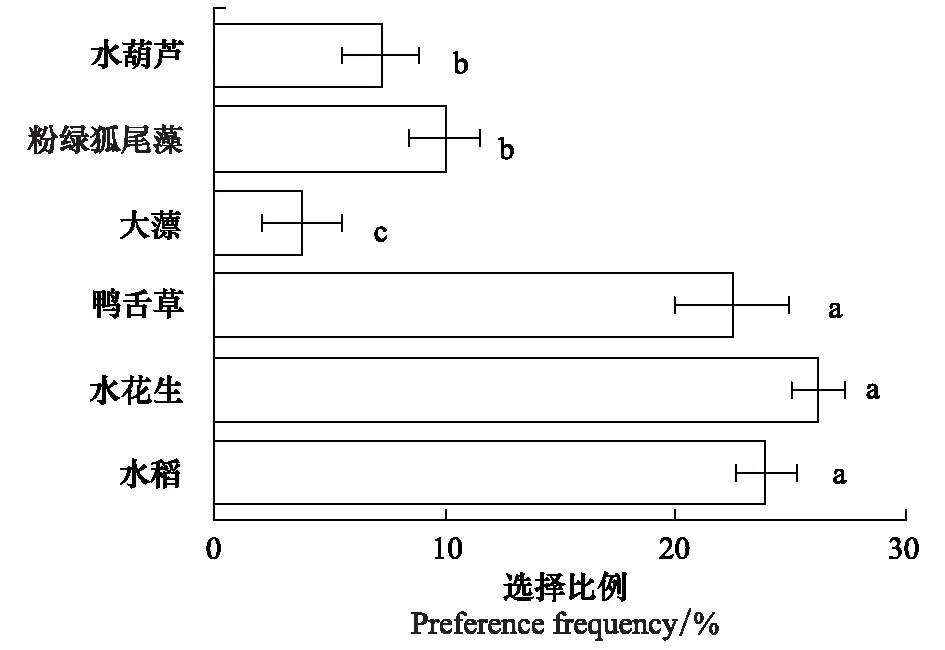
图7 福寿螺对不同水生植物的摄食选择频率Fig.7 Food preference frequency of Pomacea canaliculata on different macrophytes
福寿螺对供试水生植物的摄食选择存在较大差异(图7)。其中,福寿螺对水稻、水花生和鸭舌草的选择比例均超过20%,而对大薸、粉绿狐尾藻类和水葫芦的选择比例明显较低。福寿螺对水稻、水花生和鸭舌草的选择比例之间无显著差异,表明在实验条件下福寿螺对这3种植物的喜好程度大致相当。福寿螺对粉绿狐尾藻和水葫芦的选择比例无显著差异,对大薸的选择性最低,表明福寿螺并不喜好粉绿狐尾藻、大薸和水葫芦。总之,福寿螺更偏向于取食水稻、水花生和鸭舌草。
3 讨论
3.1 福寿螺密度和稻田水生植物密度变化趋势
福寿螺引入对各种供试植物生长密度变化趋势的影响可以分为3种情况,一种是前期降低后期稳定,如水稻。第2种是前期降低而后升高稳定,如水花生和鸭舌草;第3种是一直上升,如大薸、水葫芦和粉绿狐尾藻。水生植物生长密度变化趋势受到福寿螺摄食、植物生长特性、资源限制和竞争等因素的影响。首先,福寿螺喜欢取食秧苗,因为其鲜嫩的茎和柔软的叶片非常适合福寿螺进食,而对返青后茎杆逐渐硬化的水稻植株取食性下降,即其危害主要表现在苗期[17]。本实验中第2周时福寿螺各密度处理中的水稻秧苗密度仍有下降,特别是高密度处理,可能的原因是第1至2周时福寿螺仍在摄食水稻秧苗,或第1周受福寿螺危害的秧苗无法正常生长而死亡,而第2周后水稻密度保持稳定,因此,有些研究认为可以通过延长水稻移栽苗龄的措施来降低福寿螺的危害[18- 19]。对水花生和鸭舌草,室内实验结果表明水花生和鸭舌草均是福寿螺喜食的植物,其生长初期(第1周)被福寿螺取食导致密度降低,然而这两种植物生长繁殖迅速,后期在福寿螺的摄食压力下密度保持稳定。大薸、水葫芦和粉绿狐尾藻均属于福寿螺不喜食植物,故其生长密度一直处于上升状态。同时,各密度福寿螺处理中其自身的日均密度增长率间没有显著差异,表明福寿螺种群的扩张在实验中未受到其自身密度制约,福寿螺种群密度的增加具备较大的潜力。
3.2 福寿螺对6种水生植物生长的影响
福寿螺对6种供试植物中的水稻秧苗、水花生和鸭舌草的摄食性高,导致水稻秧苗、水花生和鸭舌草的日均密度增长率下降,尤其是水稻秧苗的日均密度增长率为负值,这也与初期水稻秧苗占主要地位有关。本课题组前期研究发现,在单一被动摄食条件下福寿螺摄食偏好为:水花生>鸭舌草>水稻[20],该研究和本实验的复合主动摄食实验方法有所不同。本实验中3种植物对福寿螺的取食偏好差异不明显,这表明在多种植物共存时,福寿螺对其喜食水生植物的选择行为受到了干扰,其取食结果则由对多种食源的喜食性及其多寡对比(即资源易获取性)而决定,因此本研究结果在一定程度上更贴近自然状态(即多种食物源共存)。有研究者试图将福寿螺做为“生物除草剂”[21],但本研究认为,福寿螺并非“见青即食”,实际上福寿螺的这种偏食性可能使某些其喜食的杂草减少,同时使得部分其他稻田水生植物(如杂草)更加泛滥。无论是主动还是被动摄食实验,福寿螺对水花生、鸭舌草和水稻秧苗的摄食性均高于其他植物,研究认为福寿螺在摄食时更偏向于摄食有利于其快速生长的水生植物[11],其原因可能是这3种植物更易于被福寿螺消化吸收。
本研究发现福寿螺对水生植物存在摄食偏好,这点和前期关于福寿螺摄食特性研究类似[22- 23],表明在田间使用福寿螺喜食的其他植物对其进行诱控是可行的,即将这些植物作为 “陷阱”植物,从而减少福寿螺对水稻的直接危害。同时本研究发现,福寿螺主要在引入稻田的1—2周内危害水稻,因此,可以考虑在目前的防治措施基础上,通过在水稻秧苗移栽期,向稻田中投放或增加一些福寿螺喜食的其他植物,以进一步降低福寿螺对秧苗的危害。
本研究中,大薸、粉绿狐尾藻和水葫芦3种入侵植物之间也存在复杂的竞争关系。大薸和粉绿狐尾藻的日均密度增长率不受福寿螺的影响,且显著高于水葫芦的日均增长率,在稻田水生植物群落中逐渐占据主要地位,在高密度福寿螺处理中二者的密度甚至达到水稻密度的近10倍。虽然有研究认为水葫芦比大薸具有较高的光能利用率[24],具有潜在的竞争优势,但在本实验中大薸、粉绿狐尾藻和水葫芦3种入侵植物同时竞争稻田资源,3种植物竞争生长的结果可能不仅仅受光能利用率的影响,而且可能还有更深层的原因,这方面有待进一步探讨。
3.3 福寿螺对稻田水生植物多样性的影响
群落结构的改变会造成生物多样性的变化,实验中影响稻田水生植物多样性的因素是福寿螺的选择性摄食和稻田非秧苗类水生植物的扩张,二者在不同时期影响不同。第1周时福寿螺各密度处理和对照间的Simpson指数、Shannon指数和均匀度指数无显著性差异,其原因在于此时期秧苗在水生植物群落中占主要位置,福寿螺大量摄食秧苗,减少了水稻数量,提高了多样性;而对照中则是稻田非秧苗类其他水生植物繁殖迅速,水稻数量不变,多样性提高。这使得二者的多样性指数接近,两种因素的影响程度相当。第2周至第8周时,水稻数量稳定,其他水生植物被福寿螺摄食状态和其扩张速度成为影响多样性指数变化的因素。实验8周后,稻田水生植物的Simpson指数、Shannon指数、均匀度均出现显著下降,可能的原因是福寿螺选择性摄食使得群落结构失调,水花生和鸭舌草密度受压制,而大薸、水葫芦和粉绿狐尾藻过度繁殖,最终导致稻田水生植物群落的物种多样性显著降低,且随着福寿螺密度增加,其摄食量变大,水生植物下降幅度愈来愈明显。对自然湿地调查研究发现,福寿螺引入后一些湿地水生植物开始消失,其摄食行为改变了湿地生态系统结构,会导致生物多样性下降,影响自然湿地生态系统服务功能[10]。在稻田人工湿地系统中,由于水稻在稻田内的主导性、福寿螺摄食秧苗的特性以及其他水生植物的迅速生长多方面的影响,在水稻种植前期,福寿螺并不会造成稻田水生植物群落的物种多样性下降,但随着时间的推移,福寿螺对稻田水生植物群落物种多样性的危害逐渐显现,即福寿螺引入稻田,在水稻生长后期会导致水生植物群落物种多样性下降,这和自然湿地的规律有所不同。
4 结论
综上所述,福寿螺是一种恶性入侵动物,其对稻田水生植物群落的影响体现在以下方面:
(1)福寿螺严重威胁水稻生产,危害性随水稻的生长逐渐降低。福寿螺引入1周内会造成秧苗密度的急剧下降,高密度福寿螺处理中水稻密度下降超过70%,到2周后水稻秧苗逐渐长大,福寿螺对其危害减轻,水稻植株密度保持稳定,各密度处理的福寿螺密度变化差异不明显。
(2)福寿螺对稻田水生植物密度的影响因种类而不同。福寿螺对水花生和鸭舌草有抑制效应,对水葫芦、大薸和粉绿狐尾藻影响不明显。福寿螺对不同水生植物的摄食选择性存在差异,其对水稻、水花生、鸭舌草的喜食性明显高于水葫芦、粉绿狐尾藻和大薸。
(3)福寿螺引入稻田后,前期水生植物群落物种多样性Simpson指数、Shannon指数和均匀度指数未发生变化,后期造成稻田水生植物群落的物种多样性逐渐降低,最终显著低于对照,福寿螺会严重影响稻田水生植物群落的物种多样性。
[1] Pimentel D, Lach L, Zuniga R, Morrison D. Environmental and economic costs of nonindigenous species in the United States. BioScience, 2000, 50(1): 53- 65.
[2] Hayes K A, Joshi R C, Thiengo S C, Cowie R H. Out of South America: multiple origins of non-native apple snails in Asia. Diversity and Distributions, 2008, 14(4): 701- 712.
[3] Carlsson N O L, Brönmark C. Size-dependent effects of an invasive herbivorous snail (Pomaceacanaliculata) on macrophytes and periphyton in Asian wetlands. Freshwater Biology, 2006, 51(4): 695- 704.
[4] Lach L, Britton D K, Rundell R J, Cowie R H. Food preference and reproductive plasticity in an invasive freshwater snail. Biological Invasions, 2000, 2(4): 279- 288.
[5] Wong P K, Liang Y, Liu N Y, Qiu J W. Palatability of macrophytes to the invasive freshwater snailPomaceacanaliculata: differential effects of multiple plant traits. Freshwater Biology, 2010, 55(10): 2023- 2031.
[6] Ye J R, Feng Y B, Lin X W, Wang H D, Zhu Z R. Host plants and their effects on body mass in the golden apple snail,PomaceacanaliculataLamarck. Journal of Biosafety, 2011, 20(2): 124- 131.
[7] Zhang J E, Fang L. Ecological issues of research for biological invasion ofAmpullariagigasSpix in China. Chinese Journal of Eco-Agriculture, 2008, 16(6): 1585- 1589.
[8] Lowe S J, Browne M, Boudjelas S. 100 of the World′s Worst Invasive Alien Species. New Zealand: the IUCN/SSC Invasive Species Specialist Group (ISSG), 2000: 6- 7.
[9] Yang Y X, Hu Y C, Li X H, Wang X J, Mou X D, Song H M, Wang P X, Liu C, Luo J R. Historical invasion, expansion process and harm investigation ofPomaceacanaliculatein China. Chinese Agricultural Science Bulletin, 2010, 26(5): 245- 250.
[10] Carlsson N O L, Brönmark C, Hansson L A. Invading herbivory: The golden apple snail alters ecosystem functioning in Asian wetlands. Ecology, 2004, 85(6): 1575- 1580.
[11] Carlsson N O L, Lacoursiere J O. Herbivory on aquatic vascular plants by the introduced golden apple snail (Pomaceacanaliculata) in Lao PDR. Biological Invasions, 2005, 7(2): 233- 241.
[12] Fu X Y, Wang H Q. Studies on breeding ecology ofampullariaGigas. Journal of Zhejiang Ocean University: Natural Science, 2000, 19(1): 37- 41.
[13] Ye J R, Li Y M. The population dynamics and distribution of golden apple snail in paddy fields. Chinese Agricultural Science Bulletin, 2009, 25(3): 185- 188.
[14] Luo S M. General Ecology. Beijing: China Agricultural Press, 2005: 116- 120.
[15] Pinowska A. Effects of snail grazing and nutrient release on growth of the macrophytesCeratophyllumdemersumandElodeacanadensisand the filamentous green algaCladophorasp. Hydrobiologia, 2002, 479(1/3): 83- 94.
[16] Li K Y, Liu Z W, Gu B H. Density-dependent effects of snail grazing on the growth of a submerged macrophyte,Vallisneriaspiralis. Ecological Complexity, 2009, 6(4): 438- 442.
[17] Wan P, Chu S H, Wu H H, Huang M S, Jiang G M. Study on occurrence, damage and control of golden apple snail,Pomaceacanaliculata(Lamarck) in paddy fields of Hubei Province. Hubei Agricultural Sciences, 2010, 49(12): 3072- 3075.
[18] Sin T S. Damage potential of the golden apple snailPomaceacanaliculata(Lamarck) in irrigated rice and its control by cultural approaches. International Journal of Pest Management, 2003, 49(1): 49- 55.
[19] Sanico A L, Peng S, Laza R C, Visperas R M. Effect of seedling age and seedling number per hill on snail damage in irrigated rice. Crop Protection, 2002, 21(2): 137- 143.
[20] Zhao B L, Dai W, Zhang J E, Cheng C G, Li G. Characteristics of feeding preference and nutrients utilization of golden apple snail (Pomaceacanaliculata) on macrophytes in paddy fields. Advance Journal of Food Science and Technology, 2012, 4(5): 316- 321.
[21] Joshi R C, Martin E C, Wada T, Sebastian L S. Role of golden apple snails in organic rice cultivation and weed management // Joshi R C, Sebastian L S, eds. Global Advances in Ecology and Management of Golden Apple Snails. Los Baos: Philippine Rice Research Institute, 2006: 483- 488.
[22] Zeng K Y, Hu F, Chen Y F, Chen J J, Kong C H. Effects of four invasive plants with the same origin area ofAmpullariagigas(golden snail) on it behaviors and killing activity. Acta Ecologica Sinica, 2008, 28(1): 260- 266.
[23] Xu W B, Zhong Q H, Li L F, Guo R R, Zhang J E, Luo M Z. The expulsive and attractive effects of 16 species of plants on golden apple snails (AmpullariagigasSpix). Guangdong Agricultural Sciences, 2010, 37(11): 149- 152.
[24] Zhu H, Ma R J. Comparison of photosynthetic characteristics between two hydrophytic invasive plants. Journal of Northwest A & F University: Natural Science Edition, 2010, 38(5): 193- 198.
参考文献:
[6] 叶建人, 冯永斌, 林贤文, 王华弟, 祝增荣. 福寿螺的寄主植物及其对福寿螺体重的影响. 生物安全学报, 2011, 20(2): 124- 131.
[7] 章家恩, 方丽. 关于我国农田福寿螺生物入侵需要加以研究的生态学问题. 中国生态农业学报, 2008, 16(6): 1585- 1589.
[9] 杨叶欣, 胡隐昌, 李小慧, 汪学杰, 牟希东, 宋红梅, 王培欣, 刘超, 罗建仁. 福寿螺在中国的入侵历史、扩散规律和危害的调查分析. 中国农学通报, 2010, 26(5): 245- 250.
[12] 傅先元, 王洪全. 大瓶螺繁殖生态学研究. 浙江海洋学院学报: 自然科学版, 2000, 19(1): 37- 41.
[13] 叶建人, 李云明. 福寿螺在水稻田的消长规律及其田间分布动态. 中国农学通报, 2009, 25(3): 185- 188.
[14] 骆世明. 普通生态学. 北京: 中国农业出版社, 2005: 116- 120.
[17] 万鹏, 褚世海, 武怀恒, 黄民松, 姜干明. 湖北省稻田福寿螺的发生规律、危害及防控研究. 湖北农业科学, 2010, 49(12): 3072- 3075.
[22] 曾坤玉, 胡飞, 陈玉芬, 陈建军, 孔垂华. 四种与福寿螺 (Ampullariagigas) 同源地入侵植物的杀螺效果. 生态学报, 2008, 28(1): 260- 266.
[23] 徐武兵, 钟秋华, 李林峰, 郭荣荣, 章家恩, 罗明珠. 16种植物对福寿螺的趋避效果研究. 广东农业科学, 2010, 37(11): 149- 152.
[24] 朱慧, 马瑞君. 2种水生入侵植物光合特性的比较. 西北农林科技大学学报: 自然科学版, 2010, 38(5): 193- 198.
Effects ofPomaceacanaliculataon aquatic macrophyte community structure in paddy fields
ZHAO Benliang1,2,3, ZHANG Jiaen1,2,3,*, DAI Xiaoyan1, PENG Li1, PENG Hui1, HUANG Weijun1
1DepartmentofEcology,CollegeofAgriculture,SouthChinaAgriculturalUniversity,Guangdong,Guangzhou510642,China2KeyLaboratoryofAgro-EnvironmentintheTropics,MinistryofAgricultureoftheP.R.China,Guangzhou510642,China3KeyLaboratoryofAgro-ecologyandRuralEnvironmentofGuangdongRegularHigherEducationInstitutions,SouthChinaAgriculturalUniversity,Guangzhou510642,China
Pomaceacanaliculata(Lamarck) is an invasive aquatic gastropod endangering rice production in Southern China, with a feeding preference for rice seedlings in paddy fields.P.canaliculatais listed as one of the top 100 invasive species by the Invasive Species Group of the World Conservation Union.P.canaliculatamainly feeds on a diverse range of macrophyte species. We investigated the invasive effects ofP.canaliculataon the aquatic macrophyte plant community in paddy fields. Our experimental macrophyte community consisted ofOryzasativaL.,PistiastratiotesL.,Eichhorniacrassipes,Myriophyllumaquaticum,Alternantheraphiloxeroides,Monochoriavaginalis. The control and three treatments used differentP.canaliculatadensities: control (P.canaliculata-free); low (4 individuals/m2); medium (8 individuals/m2) and high (12 individuals/m2). We analyzed the effect thatP.canaliculatahad on the macrophyte community using the compound food preference test. Our results showed: 1)O.sativadensities in the three treatments decreased rapidly one week afterP.canaliculatawas introduced. In herbivore, in theP.canaliculatahigh-density treatment,O.sativadensity declined approximately 70%. However, two weeks afterP.canaliculatawas introduced, the herbivore effect onO.sativadensity decreased. 2) After eight weeksP.canaliculatareducedA.philoxeroidesandM.vaginalisdensities (30% and 25%, respectively) in the high-density herbivore treatment and inhibited the growth ofA.philoxeroidesandM.vaginalis. The plant densities ofA.philoxeroidesandM.vaginaliswere higher in the low-density herbivore treatment compared with those in the medium and high-density herbivore treatments. However,P.canaliculatashowed no significant effect on the plant densities ofE.crassipes,P.stratiotesandM.aquaticum. 3) The daily density rate of increase was negative forO.sativa, and significantly different in the low, medium and high-density herbivore treatments. The daily density rate of increase forO.sativashowed a significant decrease compared with theP.canaliculata-free control. The daily density rate of increase forA.philoxeroidesandM.vaginaliswas positive, where the rate was significantly lower compared with theP.canaliculata-free control. Concurrently, the daily density rates of increase were not significantly influenced byP.canaliculataforP.stratiotes,E.crassipesandM.aquaticum. We found thatP.canaliculatapopulations increased rapidly. The daily density rate of increase forP.canaliculatapopulations in the three different density treatments were all high, but not significantly different. The food preference frequencies ofP.canaliculataonO.sativa,A.philoxeroidesandM.vaginaliswere all significantly higher compared withE.crassipes,P.stratiotesandM.aquaticum. 4) Eight weeks after the introduction ofP.canaliculatathe Simpson, Shannon and Homogeneity biodiversity indices showed a significant decline in the aquatic macrophyte community. The Simpson index values showed the decline in order of: the control > low- > medium-and high-densities. The Shannon and Homogeneity indices showed the decline in order of: the control > low- > medium- > high-densities. We conclude thatP.canaliculatashowed significantly different feeding preference on macrophyte plants species. The invasion ofP.canaliculatawould significantly endanger aquatic macrophyte community biodiversity in the paddy fields of China.
Pomaceacanaliculata; invasion; macrophyte; biodiversity; food preference
国家自然科学基金资助项目(U1131006,30900187,30770403); 广东省科技计划资助项目(2007B020709007,2011B020309009); 广东省高等学校高层次人才资助项目(粤教师函2010[79 号]); 广东省引进国(境)外高层次人才智力资助项目(粤外专[2010]51 号); 华南农业大学校级创新训练资助项目(SCX12104); 华南农业大学校长科学基金资助项目(2009K029)
2012- 04- 03;
2013- 01- 10
10.5846/stxb201304030601
*通讯作者Corresponding author.E-mail: jeanzh@scau.edu.cn
赵本良,章家恩,戴晓燕,彭莉,彭卉,黄伟均.福寿螺对稻田水生植物群落结构的影响.生态学报,2014,34(4):907- 915.
Zhao B L, Zhang J E, Dai X Y, Peng L, Peng H, Huang W J.Effects ofPomaceacanaliculataon aquatic macrophyte community structure in paddy fields.Acta Ecologica Sinica,2014,34(4):907- 915.
