凉水自然保护区不同林型地表甲虫物种多样性1)
2014-08-02刘哲强王利东
顾 伟 马 玲 刘哲强 焦 玥 王利东 张 琛
(东北林业大学,哈尔滨,150040)
凉水自然保护区不同林型地表甲虫物种多样性1)
顾 伟 马 玲 刘哲强 焦 玥 王利东 张 琛
(东北林业大学,哈尔滨,150040)
采用巴氏罐诱法,对凉水自然保护区5种主要林型地表甲虫组成和数量进行调查,分析了该地区地表甲虫多样性的变化趋势及主要气候环境因子对地表甲虫群落的影响。结果表明:调查共诱捕地表甲虫28种,隶属7个科,埋葬甲科和步甲科为优势类群;不同林型多样性指数除人工红松林外,差异均不大;聚类分析表明,原始红松林天然演替系列林型地表甲虫群落的组成结构较人工林存在较大区别;地表甲虫群落时间动态反映出,原始红松林多样性指数、均匀度指数和丰富度指数变化趋势在整个生长季均较为稳定,而人工红松林波动较大,时间序列上各生境地表甲虫多样性和丰富度变化趋势保持一致;土壤温度对各生境地表甲虫群落均产生较大影响,降雨量对天然植被演替生境中地表甲虫群落有重要调控作用。
地表甲虫;群落结构;多样性;生境类型;聚类分析;凉水自然保护区
生物多样性是人类生存和发展的基础,是生态系统可持续性的前提[1]。森林是生物多样性保护和保存的重要场所,人为活动对森林的生物多样性产生直接和间接的影响。地表甲虫是昆虫类群中种类最丰富的鞘翅目甲虫的一部分,其物种的组成和数量变化可以作为监测环境变化的重要指标。近年来,对此类甲虫的研究已成为生物多样性研究的重要内容,主要集中在对森林、湿地、荒漠等区域不同植被生境类型地表甲虫群落的研究[2-8],以及对特定区域不同功能区变化格局的探讨[9-10]和不同林龄比较[11]。凉水国家级自然保护区位于小兴安岭南端,是以保护原始阔叶红松林生态系统为主的森林和野生动物类型的保护区,区内保存有较大面积的原始阔叶红松林,同时也分布有不同类型的天然次生林和人工次生林,为开展小兴安岭不同森林生境的研究提供了良好的环境。对该保护区昆虫多样性的研究开展的较少,仅见殷秀琴等[12]和安静超等[13]对土壤昆虫群落多样性的研究报道,对地表甲虫组成及多样性的研究尚未开展。本研究选取凉水自然保护区内具有代表性的5种林型为研究地,调查分析各林型内地表甲虫多样性,探讨地表甲虫群落变化对生境类型和主要气候环境因子的生态响应,旨在为小兴安岭森林生态可持续发展规划、生物多样性保护及利用提供理论基础和科学依据。
1 研究区概况
凉水自然保护区位于小兴安岭山脉的南段达里带岭支脉的东坡,地带性土壤为暗棕壤,地带性植被是以红松为主的温带阔叶红松林,具有明显的温带大陆性季风气候特征,冬季寒冷干燥、夏季温热多雨,年平均降水680 mm左右,年平均气温-0.3 ℃,年平均最高气温7.5 ℃,年平均最低气温-6.6 ℃,≥10 ℃的积温为1 700 ℃[14]。本研究选取5种主要林型,分别为人工云杉林(A)、天然次生白桦林(B)、人工红松林(C)、次生阔叶红松林(D)、原始阔叶红松林(E)。其中,B、D和E为天然林,且属于小兴安岭植被自然演替从初级到顶级的典型演替序列。A和C属于人工次生林。每种林型设置2块样地,间距大于200 m,样地面积100 m×100 m。
2 研究方法
2.1 调查方法
取样时间为2012年6—9月份,每隔14 d取样一次,遇雨天顺延。采用巴氏罐诱法[15]采集标本。用一次性塑料水杯(高9.0 cm,口径7.5 cm)作为巴氏罐诱法容器,每块样地内设诱杯50 个,诱杯间距约1 m。引诱剂为醋、糖、医用酒精和水的混合物,质量比为2∶1∶1∶20,每个诱杯内放引诱剂40~60 mL[6]。诱杯放置时间3~5 d。将采集到的地表甲虫标本用75%酒精保存,带回实验室分类统计,标本鉴定主要依据《中国经济昆虫志》、《中国动物志》等相关论著, 并在有关专家的帮助下完成。
2.2 数据处理
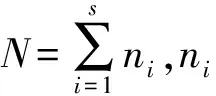
3 结果与分析
3.1 地表甲虫群落组成与数量分布
调查共捕获昆虫标本1150号,分属7科28种。其中埋葬甲科和步甲科为地表甲虫群落的优势类群,个体数量分别占总个体数量的46.52%和43.04%,花萤科、天牛科、叩甲科和隐翅虫科为该地区常见种,个体数量占总个体数的1%~10%,叶甲科个体数量不足1%,为该地区地表甲虫群落的稀有种(表1)。云冷杉林、次生阔叶红松林和原始阔叶红松林地表甲虫群落优势类群均为埋葬甲科,分别占各自群落总个体数的70.37%、50.00%和59.65%,天然次生白桦林和人工红松林为步甲科,分别占各自群落总个体数的65.22%和77.14%。不同林型中各科的种类数分布与个体数量分布表现出一致的规律。
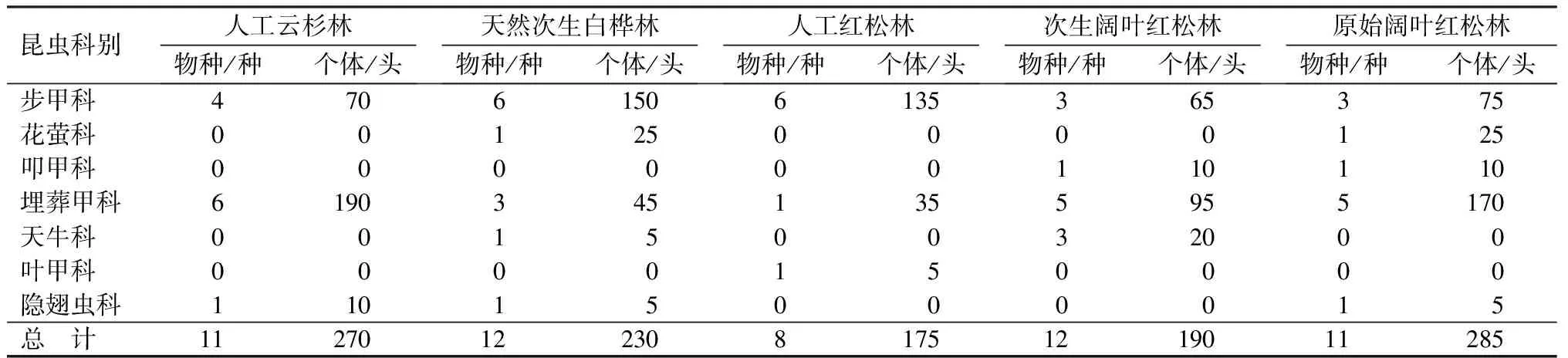
表1 凉水自然保护区各林型昆虫组成
3.2 不同林型地表甲虫群落多样性差异
由表2可见,不同林型多样性指数除人工红松林外,差异均不大,从大到小顺序为B(2.150 9)、A(2.034 2)、D(2.020 8)、E(2.005 9)、C (1.755 0)。均匀度指数从大到小顺序为B(0.865 6)、A(0.848 3)、C(0.844 0)、E(0.836 5)、D(0.813 2)。丰富度指数表现为白桦林和次生阔叶红松林最大,其次为云杉林和原始阔叶红松林,人工红松林最小。群落个体数量由多到少依次为E、A、B、D、C。其中多样性指数和均匀度度指数均为白桦林最大,人工红松林昆虫种类和个体数量均最少。群落优势度指数表现为人工红松林最大,白桦林最小,从大到小依次为C、D、E、A、B。通过相关性分析表明,多样性指数与丰富度指数显著正相关(r=0.946,P<0.05),与均匀度指数弱相关(r=0.238),与优势度指数显著负相关(r=-0.827,P<0.05)。
3.3 不同林型地表甲虫群落相似性
以5种林型样地内地表甲虫科数和数量进行聚类分析(图1),结果将5种林型生境分为3类,天然次生白桦林和次生阔叶红松林最为相似,原始阔叶红松林次之,人工红松林和人工云杉林与其它林型差异均较大。
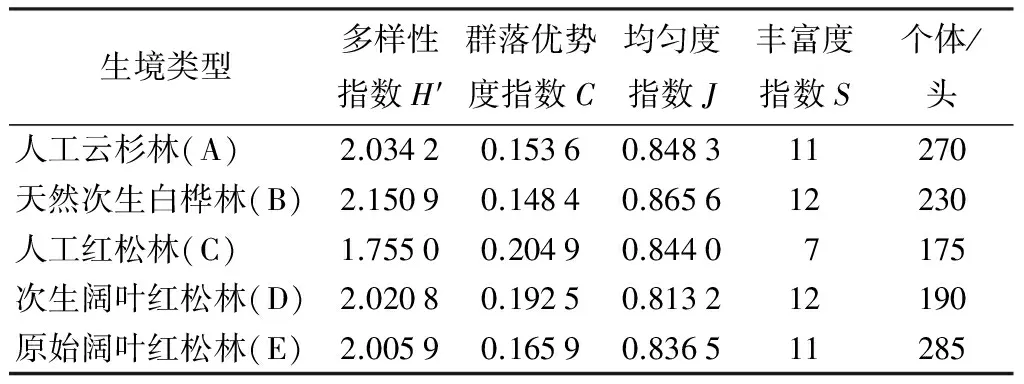
表2 凉水自然保护区不同林型地表甲虫多样性
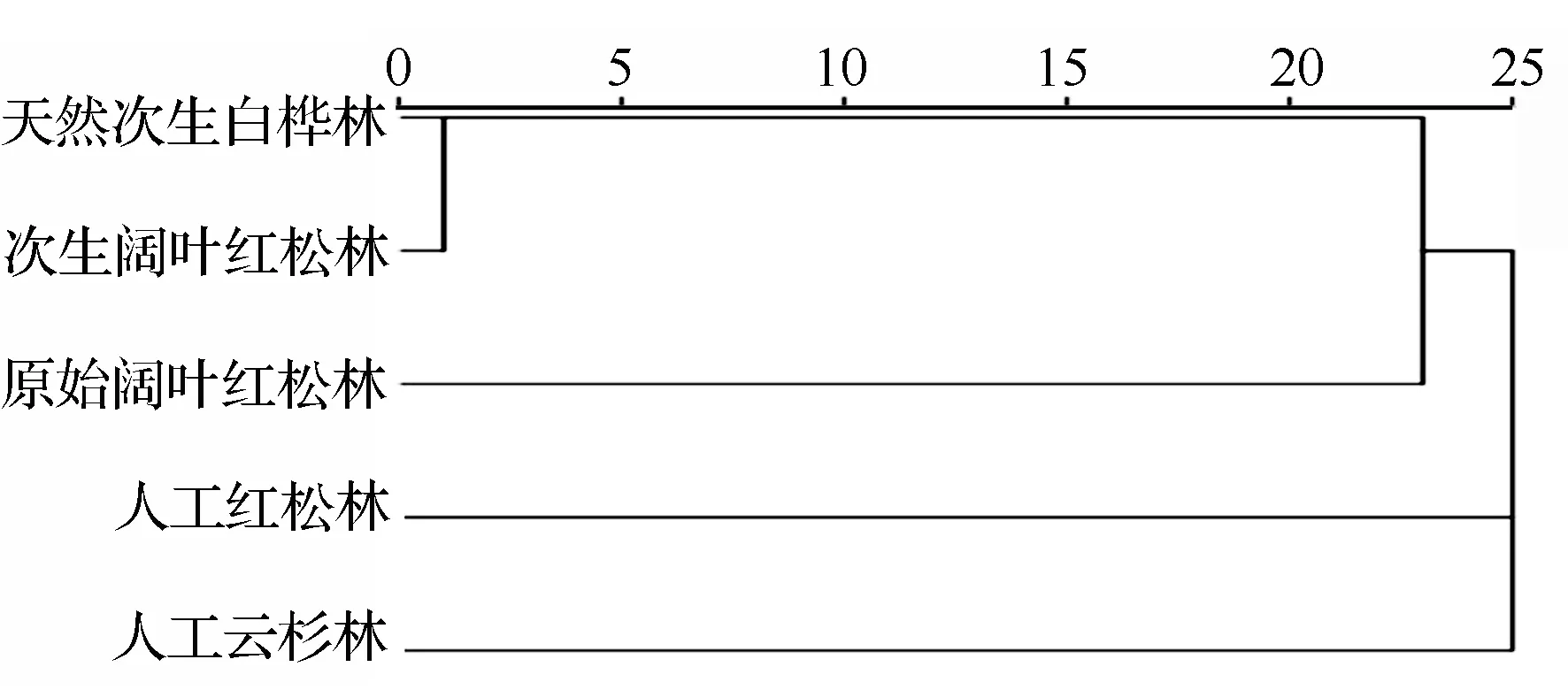
图1 凉水自然保护区不同林型地表甲虫群落的聚类图(欧氏距离)
3.4 不同林型地表甲虫群路多样性季节动态
不同林型生境昆虫群落Shannon多样性指数的时间动态如图2所示:A样地7月较高,8月中旬最低,至8月末出现第2个高峰值;B样地在7月中旬和8月末出现2个高峰值,7月末较低;C样地在7月末和8月末较高,9月较低;D样地在7月末和8月末出现2个高峰值,8月中旬最低;E样地在6月末至8月中旬逐渐升高,最后逐渐降低。其中,A、B、C和D样地均出现2个高峰值,E样地只在8月中旬出现1个高峰值,整个生长季群落波动较小。各样地均匀度指数时间动态表现为:A样地和D样地变化趋势一致,在8月中旬均匀度为0,其他时期波动较小;B样地6月末至7月末逐步上升,后逐步下降;C样地7月末至8月中旬下降,随后逐步上升;E样地6月末至7月中旬逐步上升,至8月中旬保持平稳趋势,随后逐步下降。各样地丰富度指数时间动态表现为:A样地和D样地变化趋势相似,在8月中旬最低,分别在7月中旬和8月末出现2个高峰值;B样地在7月末最低,7月中旬和8月末出现2个高峰值;C样地全年只在7月末出现1个高峰值;E样地仅在8月末出现1个高峰值。各样地地表甲虫群落Shannon多样性指数与丰富度指数变化趋势表现出一致性(相关系数r=0.910~0.975,P<0.05),除A和C样地多样性指数与均匀度指数表现为显著相关(相关系数r=0.885~0.964,P<0.05)外,其它样地群落均表现为弱相关。多样性指数与优势度指数均表现为显著负相关或弱相关(相关系数r=0.067~-0.997,P<0.01)(表3)。
表3 凉水自然保护区不同生境下群落多样性指数(H′)与其他多样性指数的相关性
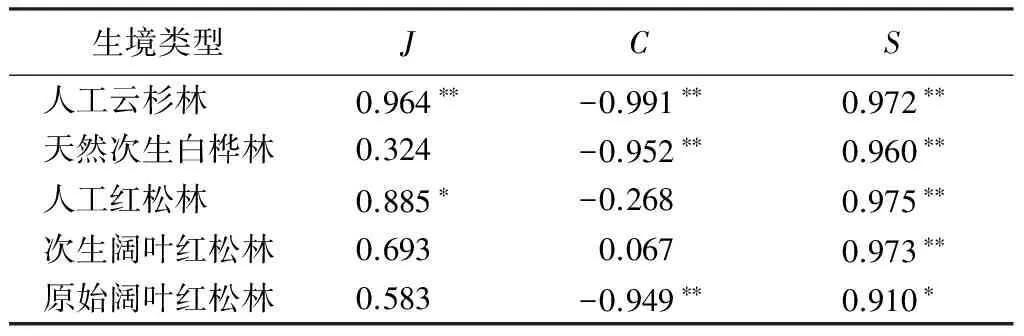
生境类型JCS人工云杉林0.964∗∗-0.991∗∗0.972∗∗天然次生白桦林0.324-0.952∗∗0.960∗∗人工红松林0.885∗-0.2680.975∗∗次生阔叶红松林0.6930.0670.973∗∗原始阔叶红松林0.583-0.949∗∗0.910∗
注:** 为0.01水平上显著相关,*为0.05水平上显著相关。
3.5 不同林型地表甲虫群落多样性与气候环境因子的相关性
不同林型地表甲虫群落多样性各特征指数的时间动态与主要气候因子相关分析结果见表4。云杉林样地地表甲虫群落多样性指数与15、20 cm土壤温度相关系数较大,表现为负相关,与降水量、相对湿度表现为弱相关;均匀度指数与20~40 cm土壤温度相关系数较大,与水汽压、10~15 cm土壤温度弱相关,与相对湿度的相关系数为0;物种丰富度表现出与多样性指数相似的规律。
天然次生白桦林样地地表甲虫群落多样性指数与各气候环境因子相关系数均较小,呈弱相关;均匀度指数与相对湿度和40 cm土壤温度表现为显著正相关,与降雨量和5 cm土壤温度相关系数较大;物种丰富度与环境因子均表现为弱相关。
人工红松林样地地表甲虫群落多样性指数与降雨量、相对湿度、5~15 cm土壤温度和40 cm土壤温度相关系数较大,其中与降雨量、相对湿度、40 cm土壤温度表现为正相关,与5~15 cm土壤温度表现为负相关,与水汽压和20 cm土壤温度为弱相关;均匀度指数与5 cm土壤温度表现为显著负相关,与相对湿度和40 cm土壤温度表现为较大的正相关,与气温、10~20 cm土壤温度表现为较大的负相关;物种丰富度指数与降雨量、相对湿度和40 cm土壤温度表现为较大的正相关,其中40 cm土壤温度呈显著正相关。
次生阔叶红松林样地地表甲虫群落多样性指数仅与降雨量表现为较大的正相关,与水汽压和相对湿度正相关,与其他环境因子表现为弱相关;均匀度指数与20~40 cm土壤温度表现为较大的负相关,与相对湿度表现为弱相关;物种丰富度与降雨量、水汽压表现为较大的正相关,与气温、5~20 cm土壤温度均表现为弱相关。
原始阔叶红松林样地地表甲虫群落多样性指数仅与40 cm土壤温度表现为显著正相关,与其他环境因子均为弱相关;均匀度指数除与水汽压、40 cm土壤温度呈显著正相关,与降雨量、相对湿度、20 cm土壤温度相关系数较大,与其他环境因子呈弱相关;物种丰富度与5、40 cm土壤温度相关系数较大,与气温、水汽压和5~10 cm土壤温度表现为负相关,其他环境因子表现为弱相关。
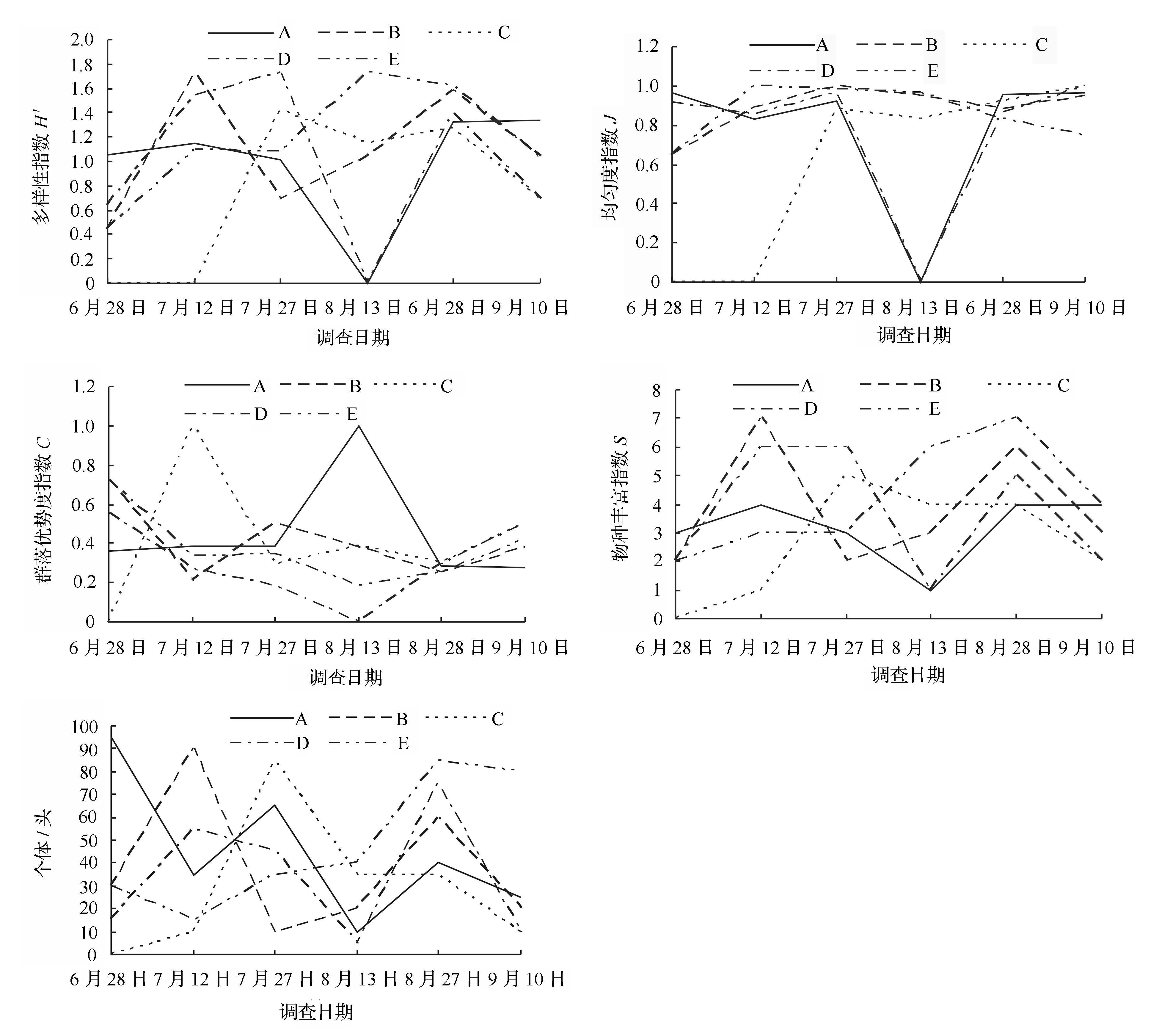
A.人工云杉林;B.天然次生白桦林;C.人工红松林;D.次生阔叶红松林;E.原始阔叶红松林。
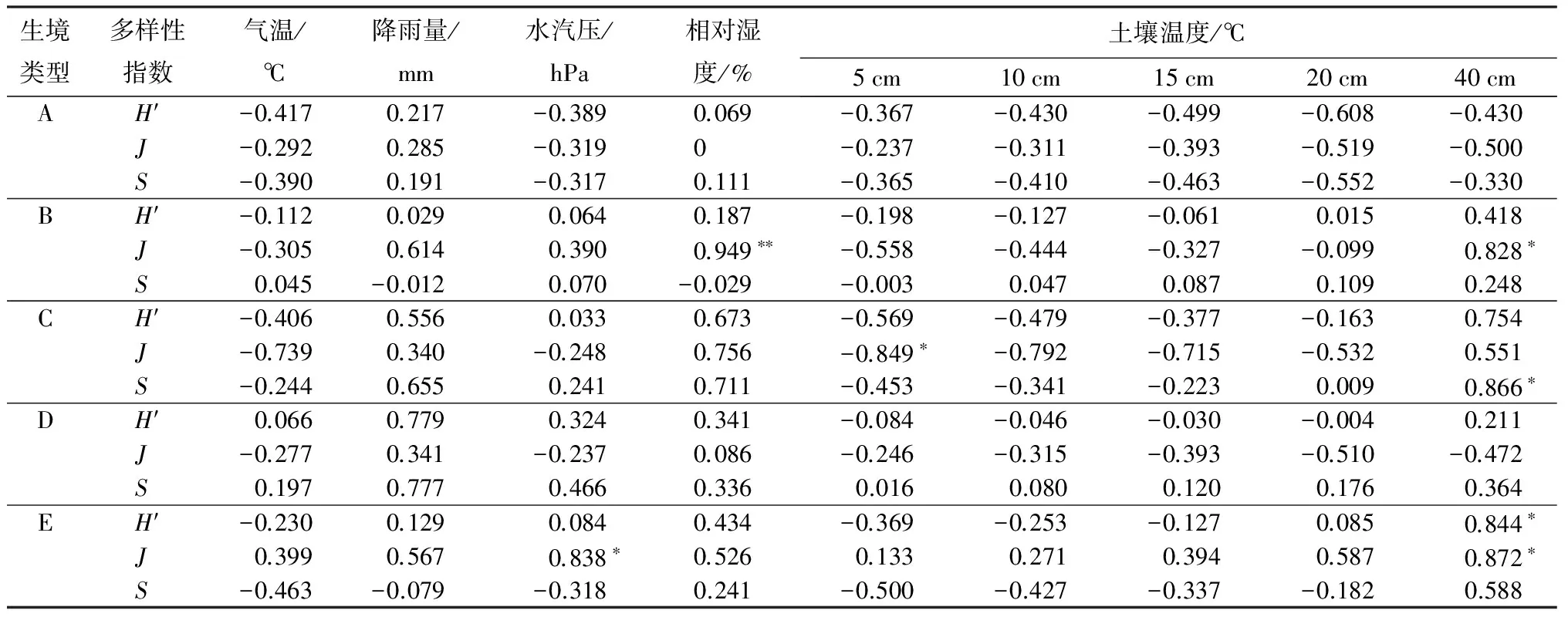
表4 凉水自然保护区不同林型下地表甲虫群落多样性指数与环境因子相关系数
注:A.人工云杉林;B.天然次生白桦林;C.人工红松林;D.次生阔叶红松林;E.原始阔叶红松林。** 为P<0.01,*为P<0.05。
4 结论与讨论
凉水自然保护区森林地表甲虫的群落组成中,埋葬甲科和步甲科个体数量分别占总个体数量的46.52%和43.04%,为研究地的优势类群,这与贾玉珍等[8]对长白山阔叶红松林的研究结果相一致。不同林型样地中,白桦林、次生阔叶红松林和原始红松林是小兴安岭天然森林植被演替的正向序列,前者的优势类群为步甲科,而后两者均为埋葬甲科。人工林中,云冷杉林中的优势类群为埋葬甲科,红松林为步甲科。笔者认为云冷杉林地表以苔藓类和蕨类为主,天然红松林地表有较厚的枯落物层,较适合埋葬甲等腐尸昆虫活动,白桦林和人工红松林地表较为干燥且枯落物层较薄更适宜步甲科捕食性昆虫生存。以往的研究[18]也表明,地表甲虫的丰富度和数量与林内的微生境(尤其是林冠层和枯落物等环境因子)密切相关。以各林型样地地表甲虫群落物种和数量为属性进行聚类分析,较为明显的将5种林型分为3类,白桦林至原始红松林等天然林类型聚为一类,2种人工林各为一类。进一步表明在森林植被自然演替过程中地表甲虫群落的组成结构发生了一定变化,同时较人工林存在较大区别。
各生境全年地表甲虫群落多样性分析,人工红松林多样性指数和丰富度指数均最小,显著小于其它林型样地,其它林型差异均不大,同时均匀度在各林型样地间差异也不大,该地区地表甲虫群落多样性指数与丰富度指数显著正相关,与均匀度指数弱相关,表明不同林型样地群落多样性受物种数量的影响较为显著。在天然演替序列中,白桦林多样性略大于次生阔叶红松林和原始红松林,说明在地带性顶级植被群落分布区中一定程度的干扰能增加昆虫群落的物种多样性。
不同林型地表甲虫群落时间动态反映出,原始红松林多样性指数、均匀度指数和丰富度指数变化趋势在整个生长季均较为平稳,人工红松林波动较大。均匀度和丰富度是与多样性密切相关的特征参数,在不同研究中结果有所不同。贺达汉等[19]认为荒漠草原昆虫群落的多样性指数与均匀度是一致的。刘文萍等[20]的研究结果表明不同生境的蝶类群落多样性指数与均匀度不一致。而在时间序列上凉水保护区各生境地表甲虫多样性和丰富度变化趋势保持一致。可见,由于该地区地表甲虫物种数量较少,导致丰富度对群落结构的影响较大。
天然林演替序列中各林型地表甲虫群落受气候环境因子的影响有一定差异,演替初期阶段受相对湿度、40 cm土壤温度的影响较大,演替中间阶段受降雨量的影响较大,到顶级阶段受40 cm土壤温度、水汽压的影响显著。人工林地表甲虫群落中,人工红松林受到各项气候环境因子的影响均较大,人工云杉林受到深层土壤温度的影响较大。笔者认为,云杉林地表以蕨类为主的地表植被盖度远远大于人工红松林,其优势类群为埋葬甲科,对此产生一定的影响。在除原始红松林外的不同林型中,昆虫群落与气候因子的相关性随着优势类群的变化而产生较大差异,步甲科占优势的群落与深层土壤温度表现为正相关,而埋葬甲科占优势的群落表现为负相关,原始红松林虽然埋葬甲科在整个生长季中表现为优势类群,但该类群是在温度逐渐降低的时期逐渐占优势,因此昆虫群落与深层土壤温度也表现为正相关。研究认为,土壤温度和降雨量对小兴安岭地带性植被演替中地表甲虫群落具有重要调控作用,在较短的时间尺度上原始红松林天然演替系列比人工林对气候环境因子变化具有更强的缓冲性,但在长期时间尺度上,地表甲虫群落对气候变化的响应还需进一步跟踪调查研究。
[1] Christensen N L, Bartuska A M, Brown J H, et al. The report of the ecological society of America committee on the scientific basis for ecosystem management[J]. Ecological Applications,1996,6(3):665-691.
[2] 刘云慧,宇振荣,王长柳,等.坝上地区农田和恢复生境地表甲虫多样性[J].生态学报,2011,31(2):465-473.
[3] 贺春霞,杜沛宜,于晓东,等.大巴山北坡地表甲虫的物种多样性[J].应用生态学报,2009,20(6):1459-1464.
[4] 于晓东,罗天宏,周红章.东灵山地区地表甲虫群落组成及季节变化[J].昆虫学报,2002,45(6):785-793.
[5] 娄巧哲,徐养诚,马吉宏,等.古尔班通古特沙漠南缘地表甲虫物种多样性及其与环境的关系[J].生物多样性,2011,19(4):441-452.
[6] 于晓东,罗天宏,周红章.横断山区东部四种林型地表甲虫的物种多样性[J].动物学研究,2004,25(1):7-14.
[7] 于晓东,周红章,罗天宏.云南西北部地区地表甲虫的物种多样性[J].动物学研究,2001,22(6):454-460.
[8] 贾玉珍,赵秀海,孟庆繁.长白山阔叶红松林不同演替阶段地表甲虫组成和多样性[J].应用与环境生物学报,2011,17(6):797-802.
[9] 于晓东,罗天宏,周红章.四川蜂桶寨国家自然保护区地表甲虫物种多样性[J].昆虫学报,2003,46(5):609-616.
[10] 冀卫荣,胡俊杰,李友莲.山西庞泉沟自然保护区步甲物种组成及多样性格局[J].昆虫学报,2008,51(9):953-959.
[11] 于晓东,罗天宏,杨建,等.卧龙自然保护区落叶松林不同恢复阶段地表甲虫的多样性[J].动物学研究,2006,27(1):1-11.
[12] 殷秀琴,吴东辉,韩晓梅.小兴安岭森林土壤动物群落多样性的研究[J].地理科学,2003,23(3):316-322.
[13] 安静超,殷秀琴.小兴安岭凉水自然保护区土壤昆虫群落特征及多样性研究[J].东北师大学报:自然科学版,2010,42(3):139-144.
[14] 马建章.凉水自然保护区研究[M].哈尔滨:东北林业大学出版社,1993:18-26.
[15] Baars M A. Catches in pitfall traps in relation to mean densities of carabid beetles[J]. Oecologia,1979,41:25-46.
[16] 赵志模,郭依泉.群落生态学原理与方法[M].重庆:科学技术文献出版社重庆分社,1990.
[17] 贾玉珍,赵秀海,孟庆繁.长白山针阔混交林不同演替阶段的昆虫多样性[J].昆虫学报,2009,52(11):1236-1243.
[18] 于晓东,罗天宏,周红章.林业活动和森林片断化对甲虫多样性的影响及保护对策[J].昆虫学报,2006,49(1):126-136.
[19] 贺达汉,田畴,任国栋,等.荒漠草原昆虫的群落结构及其演替规律初探[J].中国草地,1988,6:24-28.
[20] 刘文萍,邓合黎.木里蝶类多样性的研究[J].生态学报,1997,17(3):266-271.
Diversity of Ground-Dwelling Beetle with Different Forest Type in Liangshui Nature Reserve/
Gu Wei, Ma Ling, Liu Zheqiang, Jiao Yue, Wang Lidong, Zhang Chen(Northeast Forestry University, Harbin 150040, P. R. China)//Journal of Northeast Forestry University.-2014,42(1).-131~135
Ground-dwelling beetle; Community structure; Diversity; Habitat types; Cluster analysis; Liangshui Nature Reserve
顾伟,男,1985年10月生,东北林业大学凉水自然保护区,助理工程师。
马玲,东北林业大学林学院,教授。E-mail: maling63@163.com。
2013年4月11日。
Q968.1
1) 林业公益性行业专项(201104069)。
责任编辑:程 红。
By pitfall trapping, the experiment was conducted to investigate the species composition and quantitative distribution of ground dwelling beetles in five kinds of main forest types on Liangshui Nature Reserve and analyze the diversity variation trend of ground dwelling beetles and the effect of climate environmental factors on ground dwelling beetles. 28 species are identified to seven families, and Carabidae and Silphidae are dominant families. There is no significant difference in the beetle diversity among the other four habitats except Korean pine plantation. By clustering, there are significant differences of composition and structure of ground dwelling beetles between the original Korean pine forest natural succession series and plantation. By the time dynamic of ground dwelling beetles, the diversity index, evenness index and richness index on the original Korean pine forest are relatively stable during the whole growing season, but the Korean pine plantation has significant fluctuations, diversity index of ground-dwelling beetles and richness index trends consistent in five habitats. Soil temperature has great influence on ground-dwelling beetles of all habitats, and rainfall has important regulation effects on ground-dwelling beetle community structure in the natural vegetation succession habitats.
