横断山脉纵谷区不同林型土壤胡敏素组分特征
2020-03-19何忠俊秋新选梁社往
周 红, 何忠俊, 肖 蒙, 秋新选, 梁社往
(1.云南农业大学 资源与环境学院, 昆明 650201; 2.云南农业大学农学与生物技术学院, 昆明 650201; 3.云南省林业调查规划院, 昆明650051)
土壤腐殖质是土壤有机质的主要存在形态,由于腐殖质复杂芳香结构的存在,使其具有明显的微生物抗性,因此土壤腐殖质是有机碳库中比较稳定的组分[1],被认为是影响森林土壤碳库的重要参考指标。以往对土壤腐殖质各组分的研究多集中于土壤腐殖酸组分(富里酸、胡敏酸),而对胡敏素组分相关报道较少,但近年来人们在对土壤腐殖质各组分研究时发现胡敏素占土壤腐殖质中绝大部分,且在碳截获、土壤结构、养分保持、生物地球化学循环等方面都有着重要作用[2]。胡敏素被定义为任何pH值条件下都不可溶的腐殖质组分,但窦森[3]利用腐殖质组成法与Pallo修改法结合能够将HM进一步分组为铁结合胡敏素(HMi)、黏粒结合胡敏素(HMc)、不溶性胡敏素(HMr),这一分组方法对土壤胡敏素组分更深入的研究奠定了良好的基础。目前,关于土壤胡敏素的研究多集中于不同施肥措施[4]、耕作措施[5]、土地利用方式[6]等方面,除此之外,胡敏素在重金属及有机污染物修复[7-8]方面的作用也不可小觑,而从不同林型植被对土壤胡敏素组分性质影响的研究少有报道。因此研究不同林型植被下土壤胡敏素组分特征对维持森林土壤碳平衡具有重要作用。
横断山脉纵谷地区主要包括滇西北的怒江州、迪庆州、丽江市、大理州的部分地区。由于特殊的地理、气候条件,该区被认为是全球生态景观类型和生物多样性最为丰富的地区之一[9];同时它也是典型生态脆弱区之一[10],因此也一直备受研究者关注。但近些年来,由于自然因素与人为因素的综合作用,横断山脉纵谷区森林资源受到了不同程度的破坏,质量下降,数量减少,森林生态系统出现退化,有的甚至退化为荒山荒地[11]。这已严重威胁到当地居民的生存和发展,因此加强该区域森林资源的保护对生态环境的建设及当地社会经济发展都具有重要意义。
因此,本研究以横断山脉纵谷区白桦林(BetulaplatyphyllaSuk.)、丽江云杉林(Picealikiangensis)、大果红杉林(Larixpotaniniivar.macrocarpa)、川滇高山栎林(Quercusaquifolioides)、丽江铁杉林(TsugaforrestiiDownie)下暗棕壤为研究对象,探讨不同林型下土壤腐殖质组分分布特征,以期从土壤胡敏素组分含量、光学性质和土壤腐殖质稳定性角度揭示不同林型植被固碳能力,为横断山脉纵谷地区植被恢复工作筛选适宜的碳汇造林模式提供参考。
1 材料与方法
1.1 研究区概况
本研究采样地位于滇西北横断山脉纵谷地区,境内地势北高南低,海拔高差悬殊,垂直气候和立体生态环境特征明显。境内水热差异显著,气候属温带—寒温带气候,年平均气温4.7~16.5℃,年极端最高气温25.1℃,最低气温-27.4℃;年日照平均时数为1 740~2 190 h;年降雨量600~1 000 mm,土地利用方式以林地为主,主要土壤类型为暗棕壤,土壤质地为黏土。
1.2 采样点地上植被状况
供试土壤采自横断山脉纵谷区,该地区植被类型主要有寒温性针叶林、寒温性阔叶林,优势树种主要有白桦林、丽江云杉林、川滇高山栎林、丽江铁杉林、大果红杉林等。林下主要植被有柳叶忍冬(LoniceralanceolataWall.)、箭竹(FargesiaspathaceaFranch.)、亮叶杜鹃(RhododendronvernicosumFranch.)、冬青(IlexchinensisSims)、鼠尾草(Salviaofficinalis)、羽叶鬼针草(Bidensmaximowicziana)、凤尾蕨(Spiderbrake)、蛇莓(Duchesneaindica(Andr.) Focke)。
1.3 样品采集与分析
本研究土样于2016年4月进行采集,在横断山脉纵谷区范围内预先进行森林类型、森林土壤的类型调查,选择该区具有代表性的森林类型(白桦林、丽江云杉林、川滇高山栎林、丽江铁杉林、大果红杉林)设置样方,样方大小设置为20 m×20 m,每个样方内采用对角线5点混合采样法,去掉表面枯枝落物,按土壤发生层(Ah,E,B,C)分别采集,最后各层按比例混合均匀,放于袋内带回实验室,经风干、过0.25 mm筛制样,供试验分析,每个样本做3次重复。采样点地理环境状况详见表1。
土壤有机碳采用重铬酸钾容量(外加热法)测定;土壤腐殖质采用腐殖质分组修改法测定[12];土壤胡敏素各组分的分离:按腐殖质组成修改法提取胡敏酸(HA)、富里酸(FA)之后剩下的残渣HM,用0.5 mol/L H2SO4溶解铁、铝氧化物和氢氧化物,然后用稀碱提取出铁结合胡敏素(HMi),再通过HF/HCl破坏土壤黏土物与土壤腐殖质的紧密结合,用稀碱提取出黏粒结合胡敏素(HMc),最后的残余物为不溶性胡敏素(HMr);用722 W分光光度计在波长400,600,465,665 nm分别测定富里酸、胡敏酸的吸光值,并计算E4/E6和色调系数(ΔlgK)。
采用Excel分析软件进行数据统计、图表制作,SPSS 17.0对各指标进行单因素方差分析,文中统计值均为3次重复下的平均值。
2 结果与分析
2.1 不同林型土壤腐殖质组成分析
由表2可得出不同林型下土壤腐殖质组成不同,同一林型不同土层间腐殖质组分皆达到显著性差异(p<0.05)。其中,除白桦林、丽江云杉林外,土壤腐殖酸、富里酸垂直变化趋势为:即Ah(腐殖质层)>E(淋溶层)>B(淀积层)>C(母质层)。5种林型土壤腐殖酸含量变化趋势为:丽江云杉林(84.02 g/kg)>川滇高山栎林(68.54 g/kg)>大果红杉林(42.15 g/kg)>白桦林(40.76 g/kg)>丽江铁杉林(16.77 g/kg),其中丽江云杉林腐殖酸含量显著高于其他4种林型,丽江铁杉林(16.77 g/kg)的5倍。综上所述,从整个剖面土壤腐殖酸的分布来看,相对于我国其他区域土壤[13],横断山脉纵谷区不同林型暗棕壤腐殖酸含量较高,这主要是由于横断山脉纵谷区森林茂密,枯枝凋落物大量累积于土壤表层,再加上冬干夏雨、干湿明显的气候环境的影响,使得土壤腐殖质含量较高。
胡敏酸、富里酸作为腐殖酸的组成部分,胡敏酸与富里酸的比值(HA/FA)可用来表示土壤腐殖化程度,HA/FA比值越大,土壤腐殖化程度越高,质量越好。从图1可看出,同一林型间不同土层HA/FA差异显著(p<0.05);不同森林类型下土壤腐殖质层HA/FA皆高于其他土层;且各林型土层间表现为差异显著(p<0.05)。分析得出,不同林型土壤腐殖化程度随着土壤发生层深度增加呈降低趋势,HA/FA值在B层、C层基本趋于稳定,5种不同林型下B层土壤HA/FA分别为0.17,0.03,0.18,0.39,0.46,C层土壤HA/FA分别为0.13,0.03,0.19,0.19,0.39。综合分析得出,不同林型植被下各土层HA/FA值为0.03~0.79,即最高为大果红杉林的腐殖质层,最低为丽江云杉林的淀积层、母质层,这说明大果红杉林下土壤腐殖化程度较高,形成良好土壤结构能力强,反之丽江云杉林较低。

表2 不同林型土壤腐殖质组分含量 g/kg
注:表中数据表示平均值±标准差,同列不同大写字母表示同一林型不同土层间差异显著,不同小写字母表示不同林型同一土层间差异显著(p<0.05,Ducan法)。

注:图中数据表示平均值±标准差,同列不同小写字母表示同一林型不同土层间差异显著,不同大写字母表示不同林型同一土层间差异显著(p<0.05,Ducan法)。
图1 不同林型土壤腐殖化程度
2.2 不同林型土壤胡敏素组分数量分析
分析表3发现,不同林型土壤胡敏素各组分中不溶性胡敏素组分皆高于可溶性组分(铁溶性胡敏素、黏粒结合胡敏素)。同一林型土壤剖面胡敏素各组分含量差异性表现为显著(p<0.05),其中4种林型(白桦林、丽江云杉林、大果红杉林、丽江铁杉林)土壤剖面变化规律均表现为Ah层>E层>B层>C层。从不同林型同一土层来看,各林型间胡敏素各组分含量表现为差异性显著(p<0.05),除各林型C层的HMi外。其中HMi最高值出现在丽江云杉林的Ah层(6.22 g/kg),最低值为大果红杉林的C层(0.25 g/kg);HMc最高值出现在丽江云杉林的Ah层(4.54 g/kg),最低值为大果红杉林的C层(0.13 g/kg);HMr最高值出现在川滇高山栎林的Ah层(187.40 g/kg),最低值为大果红杉林的C层(4.84 g/kg)。综上所述,5种不同林型中丽江云杉林胡敏素组分含量普遍高于其他4种林型。

表3 不同林型土壤胡敏素组分情况 g/kg
注:表中数据表示平均值±标准差,同列不同小写字母表示同一林型不同土层间差异显著,不同大写字母表示不同林型同一土层间差异显著(p<0.05,Ducan法)。
2.3 不同林型土壤可溶性胡敏素光学性质分析
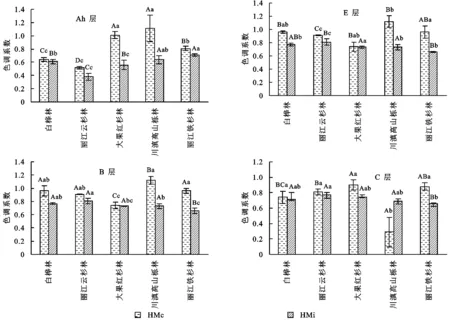
传统观点认为,土壤腐殖质的颜色与土壤腐殖化程度一一对应,因此,色调系数(ΔlgK)、光密度值(E4/E6)被用来表征土壤腐殖质分子结构的复杂程度,色调系数与光密度值呈正相关关系,色调系数越高,光密度值越高,化学稳定性越高,其分子结构越简单[14]。因此,本文采用ΔlgK,E4/E6作为参考指标来探讨可溶性胡敏素组分分子结构。从图2可以看出,不同林型土层HMi,HMc色调系数各不相同,除川滇高山栎林C层外,其他林型各土层HMc的色调系数皆高于HMi色调系数,这可说明土壤HMc的分子结构比HMi分子结构简单,但其复杂程度相差不大。E4/E6所表现的变化趋势与色调系数并不全然一致(图3),除丽江铁杉林各土层表现为E4/E6表现为HMc大于HMi外,其他林型土层间E4/E6未表现明显变化规律。除川滇高山栎林外,其他林型土层间都达到了显著差异(p<0.05)。从Ah层来看,除白桦林外,其他林型皆表现为HMi 土壤有机碳主要来源于植物残体等地表凋落物的输入[15],土壤腐殖质作为有机碳主体,在土壤固碳方面发挥着重要作用。本研究中不同林型土壤腐殖酸(HE)大小变化为:丽江云杉林(84.02 g/kg)>川滇高山栎林(68.54 g/kg)>大果红杉林(42.15 g/kg)>白桦林(40.76 g/kg)>丽江铁杉林(16.77 g/kg),其中丽江云杉林含量显著高于其他4种林型。有研究表明[16],土壤剖面腐殖酸的垂直分布主要受植被类型、根系分布的影响。这可能是因为丽江云杉林下植物残体量较高,根系固碳能力较强,进而提高了土壤有机碳的输入。随着土壤剖面深度的增加,土壤腐殖酸含量逐渐降低,表层枯枝落叶和根系分解为土壤提供了丰富的碳源,而深层土壤相对表层土壤缺乏碳源、能量,微生物活性降低,从而影响了土壤有机碳腐殖化过程,进而影响土壤腐殖质含量。胡敏酸、富里酸是土壤腐殖酸的组成部分,而腐殖质各组分含量高低取决于形成量和分解量的大小,即取决植被类型和根系分布、微生物活性等因素。不同林型土壤胡敏酸(HA)含量变化规律与腐殖酸一致,而富里酸则表现为丽江云杉林>川滇高山栎林>白桦林>大果红杉林>丽江铁杉林,其中白桦林富里酸含量高于大果红杉林,这可能是因为不同林型植被下土壤微生物数量、多样性差异及凋落物分解速率不同。有研究表明,土壤腐殖质的形成主要靠微生物的聚合作用,土壤微生物能利用富里酸组分中氨基酸等含氮化合物,不断地转化成胡敏酸[17]。 土壤胡敏酸较富里酸更为活跃,分子量较大,微生物抗性较强,对良好土壤结构形成、养分保持方面占重要地位。HA/FA是衡量土壤腐殖质品质优劣的重要指标,即胡敏酸含量越高,分子量越大,分子结构复杂,腐殖质品质越好。本研究中,不同林型土壤HA/FA大小变化为:大果红杉林>川滇高山栎林>丽江云杉林>白桦林>丽江铁杉林,其中大果红杉林最高,且皆高于其他4种林型。这说明大果红杉林下土壤腐殖质品质相对其他4种林型较好,此外,5种不同林型土壤HA/FA均小于1,其变化范围为0.03~0.79,综合来看,5种土壤腐殖质品质总体较差,主要为富里酸型土壤。随着土壤剖面深度的增加,HA/FA总体呈下降趋势,其中土壤Ah的腐殖化程度明显高于其他土层,这与党亚爱等[13]研究结果不一致,这可能与横断山脉纵谷区夏季温暖多雨、冬季寒冷干旱的气候环境有关,其次森林植被覆盖度高,人类活动干扰少有关,因此表层土壤有机质腐殖化过程进行加快。 注:图中数据表示平均值±标准差,不同小写字母表示不同林型同一土层各组分差异显著,不同大写字母表示不同林型同一土层各组分差异显著(p<0.05,Ducan法)。 图2 不同林型土壤可溶性胡敏素的色调系数 土壤胡敏素(HM)是不同于HA,FA的独立的腐殖物质组分,它占土壤有机碳的绝大部分,在土壤肥力、生态环境方面都占有重要地位。本研究中土壤胡敏素各组分数量各有差异,从不同层次土壤胡敏素组分主要分布情况分析,不溶性胡敏素组分含量占胡敏素绝大多数,其次是铁结合胡敏素、黏粒结合胡敏素。腐殖质结合态与土壤酸碱度有关[18],本研究供试土壤pH值范围在3.99~5.53,酸性土壤中铁铝氧化物成为土壤主要胶结物质,从而铁结合态胡敏素相对黏粒结合态胡敏酸较高。从不同土壤发生层来看,胡敏素各组分含量随着土层深度增加而降低,其主要变化规律为:腐殖质层>淀积层>淋溶层>母质层。从不同森林类型来看,白桦林、丽江云杉林、大果红杉林、川滇高山栎林的黏粒结合胡敏素(HMc)含量表现为差异显著(p<0.05),其中白桦林、丽江云杉林皆大于大果红杉林、川滇高山栎林,即泥质页岩坡积物母质发育的土壤黏粒结合胡敏素高于花岗岩坡积物母质。这是因为胡敏素在腐殖质组分中的占比大小,一般与土壤中的黏土矿物和无定形铁、铝氧化物的含量呈正相关。由此可知当土壤成土母质相同时,不同林型对土壤胡敏素组分的影响不大。 注:图中数据表示平均值±标准差,不同小写字母表示不同林型同一土层各组分差异显著,不同大写字母表示不同林型同一土层各组分差异显著(p<0.05,Ducan法)。 图3 不同林型土壤可溶性胡敏素的E4/E6 从色调系数指标来看,5种不同林型植被下大部分土壤铁结合胡敏素(HMi)的色调系数值小于黏粒结合胡敏素(HMc)色调系数,HMi色调系数大小与HMc相差不大;但从E4/E6来看,HMi与HMc在不同林型土壤间变化规律不明显。综合ΔlgK,E4/E6变化规律分析,土壤Ah层HMi与HMc的分子结构较其他土层复杂,这与张志春等[19]研究结果不一致,有研究表明随着土壤温度、含水量增高更有利于HMi,HMc的分子结构复杂化。但从其他土层HMi与HMc光学性质变化规律来看,HMi,HMc的分子结构不仅仅主要受温度、含水量的影响[20],可能和时间作用长短因素有关。此外,即不同林型植被所产生有机物与土壤腐殖质分子结构复杂程度有关,本研究中数据也仅是验证了这一观点,不同植被所产生的有机物与土壤腐殖质分子结构间影响机制还有待深一步研究。各林型间HMi,HMc分子结构变化规律不明显,其中HMc的分子结构在川滇高山栎林的C层最为复杂,而HMi的分子结构在丽江云杉林的Ah层最为复杂。总而言之,土壤腐殖化进程是一个复杂且受到多种因素共同作用且长久的变化过程。从两个参考指标的变化规律看,E4/E6值在不同林型土壤间的变化幅度比色调系数大,E4/E6值对各种影响因素的响应能力较色调系数更高。 (1) 横断山脉纵谷区5种不同林型暗棕壤丽江云杉林下腐殖酸含量显著高于其他4种林型。不同林型暗棕壤HA/FA表现为大果红杉林>川滇高山栎林>丽江云杉林>白桦林>丽江铁杉林,大果红杉林相对于其他林型土壤腐殖质聚合度较高、品质较好。 (2) 横断山脉纵谷区5种不同林型暗棕壤胡敏素组分中不溶性胡敏素占绝大部分。丽江云杉林下胡敏素组分含量最高。相对于其他4种林型,丽江云杉林更有利于土壤碳的积累。 (3) 横断山脉纵谷区5种不同林型暗棕壤Ah层HMi,HMc的分子结构较其他土层复杂,各林型间HMi,HMc分子结构变化规律不明显,除川滇高山栎林的C层外,各林型土层ΔlgK皆表现为HMc>HMi,但E4/E6变化规律并不总是表现为HMc>HMi;其中HMc的分子结构在川滇高山栎林的C层最为复杂,而HMi的分子结构在丽江云杉林的Ah层最为复杂。 综上所述,大果红杉林、丽江云杉林更有利横断山脉纵谷区碳素的利用、积累,该研究结果可为提高横断山脉纵谷区森林固碳能力提供参考。本文仅就横断山脉纵谷区5种主要林型腐殖质特征进行研究,具有一定的局限性。不同林型土壤固碳量有所差异,因此加强筛选相对维持高碳水平的树种对增汇减排具有重要意义。3 讨 论
3.1 不同林型土壤腐殖质组分分布特征
3.2 不同林型土壤胡敏素组分分布特征

3.3 不同林型土壤可溶性胡敏素光学性质特征
4 结 论
