高速跑时下肢双关节肌做功特性及拉伤风险的生物力学分析
2014-07-19钟运健傅维杰魏书涛
钟运健,刘 宇,傅维杰,魏书涛,伍 勰,李 庆
高速跑时下肢双关节肌做功特性及拉伤风险的生物力学分析
钟运健1,2,刘 宇2,傅维杰2,魏书涛2,伍 勰2,李 庆3
目的:建立下肢肌肉功能模型及下肢骨骼肌肉系统模型,量化快速跑时下肢双关节肌长度速度变化及应力变化,并进一步量化单位面积功率及做功总量,从生物力学视角分析短跑运动员下肢双关节肌做功特性及拉伤风险。方法:采集优秀短跑运动员高速跑时运动学和动力学数据,建立下肢神经肌肉骨骼模型和下肢肌肉功能模型,进行下肢关节运动学和动力学分析,并计算受试下肢肌肉功能参数变化,结合最优化算法量化单块肌肉应力变化,进而分析单块肌肉单位面积功率和做功总量。结果与结论:双关节肌应力峰值、长度峰值、收缩速度峰值和做负功单位面积功率峰值均显著高于其他下肢单关节肌;腓肠肌在支撑前期被拉长的情况下对抗较大外负荷(高应力值)离心收缩,且出现做负功的单位面积功率峰值,拉伤风险极高;腘绳肌在摆动末期和支撑前期处于较大程度拉伸且应力值高,在摆动中期的屈髋伸膝阶段快速做负功(出现单位面积功率峰值),均易被拉伤;股四头肌的拉伤时相为摆动前期的伸髋屈膝阶段,股四头肌在此阶段一直做负功且出现做负功的单位面积功率峰值;双关节肌在一个完整步态中所做负功总和(负功积累)显著高于其他单关节肌。此外,双关节肌在步态某些阶段,同时对两个关节分别做负功和正功。本研究通过单块肌肉力学负荷相关指标为分析下肢双关节肌肉在快速跑时拉伤风险提供了生物力学依据。
快速跑;双关节肌;拉伤风险;生物力学
前言
下肢肌肉中的腘绳肌、股直肌和腓肠肌在运动中很容易发生急性损伤,特别常见于高水平的短跑、足球和橄榄球运动员[15,16,33]。虽然运动中的各种技术动作(如踢球、身体扭转、跳跃等)均能使上述双关节肌的负荷发生改变,但研究发现,运动员的受伤均发生在接近或达到其高速跑阶段[16,33]。据此,分析下肢肌肉快速跑的力学特性对于理解双关节肌的损伤风险和机理非常关键。
肌肉损伤发生的内在原因多种多样,大部分肌肉的急性拉伤和单块肌肉力量(应力)息息相关。从力学角度来看,拉伤是过载的结果(外部力量或内部肌力)[9],即肌肉在拉伤瞬间承受负荷的能力低于当时肌肉所受负荷,故肌肉应力是肌肉拉伤的重要影响因子[20,21]。本研究团队在前期研究中发现,快速跑时双关节肌腘绳肌、股直肌和腓肠肌在整个步态中的应力峰值也显著高于其他肌肉[10]。但肌肉在向心收缩情况下承受负荷的能力较强,即使负荷很高也不容易被拉伤。 Transfeldt等(2001)研究发现,离心练习是唯一伴随着肌肉拉伤的形式[31]。但离心收缩也不是肌肉拉伤的唯一风险因子,如缓慢的肌肉牵张练习,这种慢牵张基本上不可能引起拉伤[22]。故Brooks 等人(2001,2006)随后提出,肌肉应力和拉伸速度的乘积(单位面积功率)大小才是肌肉拉伤的重要指标[14,15]。此外,Anthony 等(2012)将一个完整步态中单位面积所做负功的总和(负功积累)作为肌肉拉伤的另一重要因素,认为肌肉对负荷都有一定的适应能力和抵抗能力,但面对重复过多过重的负荷,身体的任何部位都会不断积累磨损或疲劳而引起肌肉组织、肌腱和韧带慢性损伤,进而使肌体对外负荷的适应能力和承受能力下降而引起肌肉拉伤[13]。综上可知,就真正的人体活动而言,单独的或其中一、两个指标并不能完全解释肌肉拉伤的原因,这也意味着单块肌肉力量(应力)、肌肉长度、肌肉收缩速度、收缩性质及做功状况变化等因素的交互作用最终导致肌肉拉伤[16]。
因此,本研究的目的是,建立下肢骨骼肌肉模型,量化快速跑时下肢双关节肌力学负荷和工作状态(应力变化、长度变化、收缩速度、单位面积功率及做功总量),并从生物力学视角分析短跑运动员下肢双关节肌工作特性及拉伤风险。
1 研究对象与方法
1.1 研究对象
本研究受试者为8名优秀短跑运动员(国际健将1名、健将3名、一级4名),100 m个人最好成绩范围10 s 27到10 s 80,年龄、身高、体重分别为22.4±3.2岁、180.7±6.3 cm、74.3±5.5 kg。所有受试者于实验前确认24 h内未从事剧烈运动,确定其下肢和足部半年内无明显损伤,解剖结构和机能正常,身体状况以及运动能力良好,理解本实验意图并签署同意书。
1.2 实验方法
采用VICON红外高速摄影系统(英国,VICON公司)摄像,摄像头为MX13,采样频率为300 Hz,软件版本为WORKSTATION5.1,MARKER球直径为14 mm,专用实验服装。
采用KISTLER三维测力台(瑞士,KISTLER公司),型号9287B,面积600×900 mm2,内置信号放大器,采样频率为1 200 Hz,通过数模转换器与VICON系统连接并同步,牢固安放于专门为本次实验在田径场内挖设的测力台坑内。
采用VISUAL3D三维分析软件(美国,C-MOTION公司),软件版本为Visual3D Version 3.91.1,对实验数据进行前期数据处理分析。
图 1 本实验MARKER标定位置图
按照实验拍摄范围的要求架设红外高速摄像机,拍摄范围约为9×2×2 m3,连接摄像机与VICON工作站。设置VICON系统并进行拍摄范围三维空间的标定。运动员充分热身,穿着专用实验服装,安放MARKER球[4,5,10,11]。运动员起跑位置大约距测力台40 m。运动员听从实验员的口令,全力奔跑,通过拍摄区。同时开启实验设备,采集实验数据,每个运动员需成功采集数据两次。
1.3 建立下肢环节动力学模型和下肢骨骼肌肉系统模型
对VICON和KISTLER采集的原始运动学数据进行筛选处理,通过软件Visual3D建立下肢多环节链接模型,将三维数据转化为二维数据,并对其进行优化处理,最后量化下肢运动学数据(各关节角度、角速度等)和动力学数据(肌力矩、惯性力矩、重力矩和外力矩等)。
根据前期研究所建立的环节互动动力学模型和建立下肢骨骼-肌肉系统模型[4,10,11],选取与下肢运动相关的11组主要肌肉(6组单关节肌和5组双关节肌,分别为臀大肌、股二头肌长头、股二头肌短头、半膜肌、半腱肌、股直肌、骼肌、股四头肌群、比目鱼肌、腓肠肌和胫骨前肌)。
1.4 建立下肢肌肉功能模型,量化快速跑时下肢肌肉功能参数
以单大卯(2003)建立下肢各运动环节相应坐标系和统计学模型所采用布里恩角,作为描述下肢多刚体系统相邻环节(坐标系)相对转动的广义坐标[2]。测量受试对象骨盆、股骨、胫骨、足部及用于回归髋-膝、膝-踝关节中心距的人体骨性形态学参数,借助广义坐标可将肌肉在不同环节坐标系中所表达的起止点(或代起止点)的坐标转换到肌肉起点所在环节坐标系中,最后计算出肌肉功能参数,也就建立了下肢肌肉功能模型。
本研究基于受测对象个体的下肢骨性形态学参数和快速跑时下肢运动学参数,通过下肢肌肉功能模型计算机应用分析系统[6]分别计算出受测对象快速跑时下肢各主要肌肉对于各关节伪力臂变化等肌肉功能参数。
1.5 量化快速跑时下肢肌肉长度变化和速度变化
首先,确定肌肉长度与肌拉力线:即肌肉附着点走向为直线的肌肉,其长度量化为起点和止点的直线连线长度,该连线称之为肌拉力线。
基于受测对象个体的下肢骨性形态学参数和快速跑时下肢运动学二维参数,通过下肢肌肉功能模型计算机应用分析系统[6],分别计算出受测对象快速跑时下肢各主要单块肌肉长度变化和速度变化。
1.6 建立优化方程,由已知的关节力矩优化求解肌应力
参照前期的研究所确立的快速跑时下肢单块肌力分布的优化求解方法[10],结合下肢动力学数据及肌肉功能参数等,建立优化方程,设定目标函数及求解方法,量化各单块肌肉应力变化。
1.7 量化下肢单块肌肉单位面积功率变化
计算下肢各单块肌肉单位面积功率Pi公式:
公式中,σi为各单块肌肉i活动时肌肉应力,vi为单块肌肉i的收缩速度。本研究定义,如果vi为正,则该肌肉为向心收缩;反之为离心收缩;单块肌肉单位面积功率为正值时,代表肌肉向心收缩做正功;单块肌肉单位面积功率为负值时,代表肌肉离心收缩做负功。
1.8 统计学方法
各参数值均用平均值±标准差方式表示,所有数据资料用Excel 2003和Spss 13.0软件进行统计分析,采用独立样本t检验分析各阶段不同肌肉间对于各参数的影响,其中,显著性水平α设为0.05。
2 研究结果
本研究将一个步幅分为支撑期和摆动期两个阶段,运动员左脚触地阶段为支撑期(临界点为标准化时刻17.7%±1.2%),而剩下的阶段为摆动期,其中,摆动期根据时相不同又划分为摆动前期和摆动后期(以大腿竖直摆在重心垂线位置为界,临界点为标准化时刻55.1%±2.3%)。
2.1 关节角度特征
在支撑期,髋关节持续伸展,膝关节先屈曲至标准步态10%左右开始伸展。摆动前期,髋关节和膝关节主要表现为屈曲。摆动后期,髋关节继续屈曲到标准步态70%左右开始伸展直至触地,膝关节伸展至标准步态90%左右开始屈曲。踝关节在整个步态中只有支撑期表现为较大程度的屈伸,支撑前期为屈曲,支撑后期为伸展(图2)。
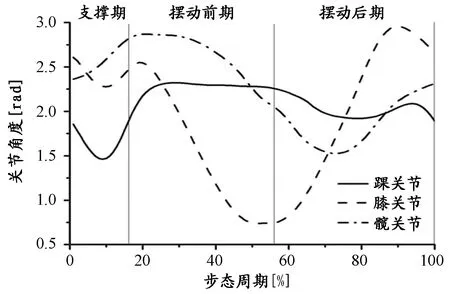
图 2 本研究某受试者完整步态中下肢各关节角度(rad)变化曲线图
2.2 肌肉功能特征
双关节肌腘绳肌(本研究主要指股二头肌长头、半腱肌、半膜肌)在整个支撑期表现为向心收缩,一直持续到摆动前期的标准步态45%左右开始离心收缩,至摆动后期的标准步态80%左右开始向心收缩,长度百分比峰值(股二头肌长头:112.5%±3.7%)出现在摆动末期(标准步态80%左右)。双关节肌股四头肌群(主要指股直肌)离心收缩阶段主要出现在摆动末期(标准步态90%左右开始)直至摆动前期的标准步态40%左右,长度百分比峰值(120.1%±4.5%)出现在摆动前期(标准步态40%左右)。腓肠肌在整个支撑期均处于被拉长状态(肌肉长度大于静息状态的100%),其离心收缩阶段主要出现在支撑前期(标准步态10%之前),整个步态的长度百分比峰值(111.5%±3.6%)出现在标准步态10%左右(图3,表1)。
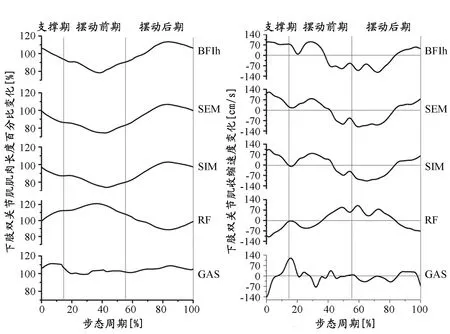
图 3 本研究某受试者部分下肢肌肉完整步态长度百分比(%)(左)和收缩速度(cm/s)(右)变化
腘绳肌、股直肌和腓肠肌在整个步态中均高速向心或离心收缩,腘绳肌向心和离心收缩速度峰值均接近120 cm/s,股直肌向心和离心收缩速度峰值分别为-109.6±7.3 cm/s和99.8±7.2 cm/s,腓肠肌的向心收缩速度峰值为119.6±8.3 cm/s,而离心收缩速度峰值高达-142.7±6.2 cm/s,分别出现在触地瞬间和离地瞬间(图3,表1)。
腘绳肌在支撑期和摆动后期处于较高的活化状态(图4),做功也主要体现在这两个阶段,腘绳肌在支撑期和摆动末期(标准步态85%以后)主要做正功,并在支撑期出现整个步态做正功的单位面积功率峰值(股二头肌长头峰值为:208.6±22.3 W/cm2),在摆动后期标准步态的55%~85%左右,腘绳肌主要离心收缩做负功,并在此阶段出现做负功的单位面积功率峰值(股二头肌长头峰值为-202.8±19.6 W/cm2)(图4,表1)。股直肌的活化主要出现在摆动前期,在标准步态20%~35%左右离心收缩做负功,单位面积功率峰值为-94.8±12.6 W/cm2,标准步态35%~55%左右向心收缩做正功,单位面积功率峰值为133.8±20.9 W/cm2(图4,表1)。腓肠肌的活化主要集中在支撑期,标准步态11%之前为离心收缩做负功,单位面积功率峰值为-149.7±38.8 W/cm2,在支撑后期主要为向心收缩做正功,单位面积功率峰值为85.2±19.8 W/cm2(图4,表1)。

图 4 本研究某受试者部分下肢肌肉完整步态肌应力(N/cm2)(左)
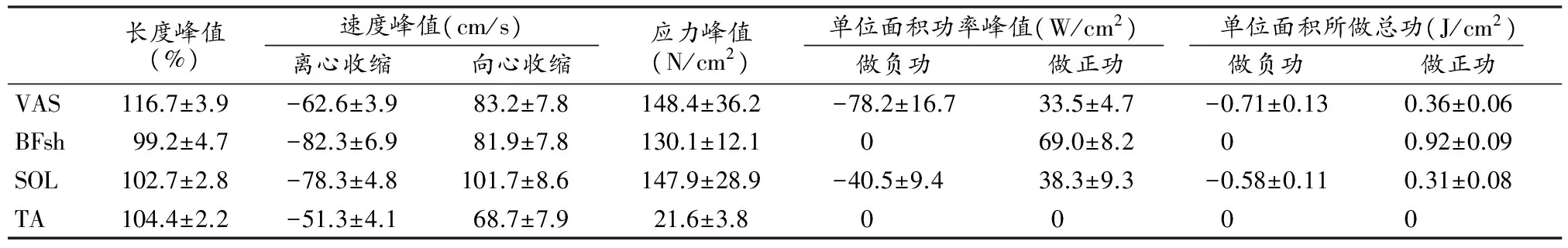
表1显示,下肢各双关节肌(腘绳肌、股直肌和腓肠肌)在一个完整步态中的长度峰值、向心及离心收缩速度峰值、应力峰值、做正功和做负功时单位面积功率峰值、单位面积所做正功总和和负功总和(负功积累)均显著高于其他单关节肌。

表 1 本研究受试者下肢主要肌肉单个步态长度、速度、应力、单位面积功率峰值及单位面积所做总功一览表
续表 1
3 讨论
就真正的人体活动而言,单独的或其中一、两个指标并不能解释肌肉拉伤。这也意味着肌肉力量(应力)、肌肉长度、肌肉收缩速度和收缩性质,甚至肌肉功率等其他因素一起综合导致真实生活中的肌肉拉伤[16]。本研究结果所展示出的双关节肌应力峰值、长度峰值、收缩速度峰值、做负功单位面积功率峰值和单个步态所做负功积累等数据,以及双关节肌在同一时刻分别对不同关节做正功和负功的复杂情况,为分析下肢双关节肌肉在快速跑时拉伤风险提供了生物力学依据。
腓肠肌:有研究通过比赛视频证实了某一棒球运动员在比赛中快速跑时腓肠肌拉伤,发生在支撑期的蹬伸阶段[24],该研究只根据这偶然获得的视频对拉伤瞬间身体重心所处的位置(腓肠肌的前方)进行了分析,未能探讨腓肠肌在此阶段被拉伤的内在肌肉力学机制。本研究显示,腓肠肌在支撑期一直处于被拉伸状态,但支撑前期离心收缩(标准步态10%之前),且整个步态的长度峰值也出现在支撑前期,而腓肠肌在蹬伸阶段(支撑后期)表现为向心收缩(图3)。此外,腓肠肌应力峰值(213.4±32.9 N/cm2)出现在支撑前期(标准步态10%左右)(图4),此时,地面反作用力背屈踝关节和伸展膝关节,为抵抗这些背屈外力矩使踝关节快速伸展蹬离地面(跖屈)[4],腓肠肌收缩很强烈。支撑阶段在高负荷的情况下,快速收缩使腓肠肌在支撑期的单位面积功率的正、负值也分别出现在蹬伸阶段和触地前期(图4)。综上所述,相比于蹬伸阶段腓肠肌向心收缩做正功而言,腓肠肌在触地前期快速离心收缩并承受强大的外负荷,出现应力峰值和做负功的单位面积功率峰值,这些肌肉力学指标都提示了腓肠肌的拉伤极易出现在触地前期,和视频所呈现的结果并不一致。这种研究结果的不同可能与其他学者提出的运动员肌肉拉伤反应延迟的假设[18,27,34]有关,如果运动员在触地前期已经拉伤,那么,也就有可能在蹬伸期承受大负荷时感到疼痛而退出运动。
腘绳肌:Simonsen等(1985)、Wood等(1987)和Yu等(2008)分析了摆动期腘绳肌的长度变化,也发现其长度峰值出现在摆动末期的快速屈髋(大腿下压)伸膝阶段,此外,上述研究也发现,腘绳肌的速度峰值出现在摆动末期的长度峰值之后,认为摆动末期为腘绳肌拉伤的危险时期,但腘绳肌在长度峰值之后的收缩形式必定为向心收缩,仅以长度峰值和向心收缩状态下的速度峰值作为其在摆动末期易拉伤的证据还稍显单薄[28,32,34]。Thelen等(2005)和Anthony等(2012)通过骨骼肌肉建模软件分别量化了快速跑时摆动阶段和完整步态中腘绳肌的力量变化,发现腘绳肌在摆动末期也出现了肌力峰值,支持摆动末期为腘绳肌拉伤的拉伤时相[13,30]。本研究的结果显示,双关节肌腘绳肌(主要指股二头肌长头、半腱肌、半膜肌)的长度峰值(股二头肌长头达静息长度的112.5%±3.7%)同样出现在摆动末期(标准步态80%左右)。其向心收缩速度曲线的3个波峰分别出现在触地瞬间、摆动中期(伸髋屈膝阶段)和摆动末期,且3个峰值没有显著的差异。和其他大部分研究结果一样[13,30],本研究的结果显示,腘绳肌在整个摆动后期活化程度很高,应力峰值(股二头肌长头峰值为:231.7±18.1 N/ cm2)也出现在摆动末期(标准步态85%左右)。
此外,和摆动末期一样,腘绳肌在支撑前期也处于较高的活化状态(图4),结合应力和速度的单位面积功率(图4)看,腘绳肌在摆动末期(标准步态80%~100%)和支撑前期(标准步态0~10%)快速做正功,而整个步态做正功的功率峰值出现在支撑前期。虽然腘绳肌在这2个时期表现为向心收缩状态(做正功,图4),但均被很大程度地拉伸(图2),这种在被极大拉长(远高于静息长度,股二头肌长头峰值高达静息长度的112.5%±3.7%)时为抵抗强大惯性力矩和外力矩而快速做功状态,使其处于被拉伤的风险之中。另外,虽然单纯从整块肌肉的长度来看,腘绳肌在摆动末期和支撑前期向心收缩做正功,但其相对于膝关节而言却被拉伸做负功,双关节在摆动末期和支撑前期这种分别对髋关节和膝关节同时做正功和负功的复杂交错状况也可能是其容易被拉伤的重要原因,也提示应进一步在肌腱和肌纤维等微观层面更为深入地探讨双关节肌在复杂运动过程中拉伤的机制。
离心收缩是导致肌肉拉伤的重要收缩状态[20,21,31,34],图3显示,腘绳肌离心收缩阶段主要在标准步态40%~80%的屈髋伸膝阶段,离心收缩速度的峰值(股二头肌长头峰值为-113.1±7.1 cm/s)也出现在标准步态的70%左右,和Anthony等(2012)[13]的研究结果类似。腘绳肌离心收缩速度峰值出现在标准步态的50%~80%之间,同时,腘绳肌在标准步态50%~70%出现一个冲击波式肌力波峰(图4),这就导致腘绳肌在摆动期中间阶段(标准步态60%~65%)出现了腘绳肌做负功的单位面积功率峰值(股二头肌长头、半腱肌和半膜肌做负功的功率峰值分别高达:-202.8±19.6 W/cm2、-184.7±17.4 W/cm2和-90.3±13.3 W/cm2(图4,表1),腘绳肌在摆动期中间阶段(屈髋伸膝)高速离心收缩下快速激活,被拉伤的可能性似乎比摆动末期(伸髋伸膝)和支撑前期(伸髋伸膝)更大,Anthony等(2012)[13]研究结果中的腘绳肌功率曲线呈现了和本研究类似的结果,但作者未对此阶段加以讨论。
股四头肌:股四头肌(主要指股直肌)拉伤常常发生于足球运动员和橄榄球运动员身上[17,24]。训练过程中发生拉伤的概率相对较高,大多数发生在与球接触期[24]。在正常快速跑步态中,双关节肌股四头肌(主要指股直肌)在摆动末期和支撑前期的伸髋伸膝阶段和腘绳肌的收缩性质相反,为离心收缩(图3)。在此阶段,股直肌的活化程度几乎为零。而事实上,在高速运动状态下尤其在支撑阶段,股四头肌的肌力为零是几乎不可能的,这种结果和本研究的研究局限性有关:本研究中的冗余肌力是将肌力矩通过数理方法进行估算,根据目前学术界的通用做法[13,23,25,30],将动作中拮抗肌的肌力简化为零,未能考虑骨骼肌的预激活和主动肌群和拮抗肌群为了固定关节等情况下的共收缩。在摆动末期和支撑前期,腘绳肌作为主动肌活化以产生肌力矩惯性力矩和地面反作用力矩[11]。此时,股四头肌作为拮抗肌参与,在正常动作情况下被拉伤的可能性较低,故将动作中拮抗肌的肌力简化为零的研究限制对本研究结果影响有限。在摆动前期的伸髋屈膝阶段,股直肌也表现为离心收缩(图3),而整个摆动前期,股直肌作为主动肌处于高活化状态(图4),这也导致其在摆动前期的伸髋屈膝阶段一直在做负功,且做负功的单位面积功率峰值出现在此时期,这些生物力学依据均提示,双关节肌股四头肌容易在摆动前期的伸髋屈膝阶段被拉伤。
一般而言,肌肉对负荷都有一定的适应能力和抵抗能力,而且重复的负荷会使适应性得到加强,但面对重复过多过重的负荷,身体的任何部位都会不断积累磨损或疲劳而引起肌肉组织、肌腱和韧带慢性损伤,进而使肌体对外负荷的适应能力和承受能力下降而引起肌肉拉伤。这种快速跑时下肢肌肉所承受的重复高负荷可细分为一个完整步态肌肉所做正功总值和负功总值。离心收缩是导致肌肉拉伤的重要收缩状态[20,21,31,34],故骨骼肌在一个完整步态中,单位面积所做负功的总和(负功积累)也是引发肌肉抵抗力下降而致其拉伤的重要影响因子[13,30]。本研究受测对象单个步态中,各肌肉单位面积所做正功和负功的值显著高于其他单关节肌肉,尤其是股二头肌长头、半腱肌、半膜肌、股直肌单位面积做负功的值分别高达-6.15±1.27 J/cm2、-3.67±0.58 J/cm2、-3.63±0.65 J/cm2、-2.02±0.33 J/cm2,腓肠肌虽然要低很多,为-0.89±0.16 J/cm2(表1),但仍然显著高于其他单关节肌,短跑运动员快速跑时,单个步态较高的负功积累也是其下肢双关节肌在快速跑时容易被拉伤的重要原因。另外,从本研究中下肢肌群各肌肉的肌肉力学指标峰值来看,股二头肌长头、半腱肌、半膜肌、股直肌和腓肠肌的长度峰值、收缩速度峰值、肌应力峰值及单位面积功率峰值均显著高于下肢其他单关节肌(表1)。快速跑时,由于快速转动产生较大的惯性力矩和外力矩[4,5,11],双关节肌抵消上述多环节的力矩快速做功所导致的高应力值致使其极易被拉伤。
4 结论
腓肠肌在支撑前期被拉长的情况下,对抗较大外负荷(高应力值)离心收缩,且出现做负功的单位面积功率峰值,拉伤风险极高;腘绳肌在摆动末期和支撑前期处于较大程度拉伸且应力值高,在摆动中期的屈髋伸膝阶段快速做负功(出现单位面积功率峰值),均易被拉伤;股四头肌的拉伤时相为摆动前期的伸髋屈膝阶段,股四头肌在此阶段一直做负功且出现做负功的单位面积功率峰值。双关节肌应力峰值、长度峰值、收缩速度峰值、做负功单位面积功率峰值和所做负功的总和(负功积累)均显著高于其他下肢单关节肌,此外,双关节肌在步态某些阶段的同一时刻有分别对两个关节做负功和正功的复杂情况。本研究通过单块肌肉力学负荷相关指标为分析下肢双关节肌肉在快速跑时拉伤风险提供了生物力学依据。
[1]刘宇.人体多关节运动肌肉控制功能的生物力学分析[M].台北:“中国文化大学”出版部,1999:6-8.
[2]单大卯.人体下肢肌肉功能模型及其应用的研究[D].上海:上海体育学院,2003.
[3]王人成,杨年峰,朱长虹,等.人体下肢摆动相冗余肌力分析[J].清华大学学报(自然科学版), 1999,39(11):104-106.
[4]魏书涛,刘宇,钟运健,等.短跑支撑期股后肌损伤的动力学分析[J].中国运动医学杂志,2009,28(6):639-643.
[5]魏书涛,刘宇,傅维杰,等.短跑控制的生物力学分析[J].体育科学,2010,30(9):37-43.
[6]伍勰,魏文仪,单大卯.下肢肌肉功能模型应用软件的开发与应用[J].医用生物力学,2008,23(3):229-233.
[7]徐力,郭巧,陈海英.跑步运动下肢冗余肌力的优化求解和分析[J].北京理工大学学报,2004,24(10):869-873.
[8]徐力,郭巧,陈海英.基于模拟退火算法的冗余肌力优化求解[J].北京生物医学工程,2005,24(2):108-111.
[9]张胜年,陆爱云.骨骼肌急性拉伤后材料力学研究进展[J].上海体育学院学报,2000,24(1):33-37
[10]钟运键,刘宇,裘艺,等.快速跑完整步态中下肢肌肉的应力变化及肌肉功能探讨[J].体育科学,2010,30(10):30-35.[11]钟运健,刘宇,魏书涛,等.优秀短跑运动员途中跑时下肢关节力矩及肌群功率分析[J].中国运动医学杂志,2011,30(1):26-31.[12]An K-N,KAUFMAN K R,CHAO E Y S.Estimation of muscle and joint forces[A].In:Allard P,Stokes I A F,Blanchi J P.Three-Dimensional Analysis of Human Movement[M].Human Kinetics,Champaign,IL,1995.201-214.
[13]ANTHONY G,SCHACHE TIMW,DORN,PETER D,etal.Mecha nics of the human hamstring muscles during sprinting[J].Official J Am College Sports Med,2012,(8):647-658.
[14]BROOKS S,FAULKNER J.Severity of contraction-induced injury is affected by velocity only during stretches of large strain[J].J Appl Physiol.2001,91:661-666.
[15]BROOKS J H M,FULLER C W,KEMP S P T,etal.Incidence,risk,and prevention of hamstring muscle injuries in professional rugby union[J].Am J Sports Med,2006,34(8):1297-1306.
[16]EKSTRAND J,HAGGLUND M,WALDEN M.Epidemiology of muscle injuries in professional football (soccer)[J].Am J Sports Med.2011;39:1226-32.
[17]HAWKINS R D,FULLER C W.A prospective epidemiological study of injuries in four English professional football clubs[J].Br J Sports Med 1999;33(3):196-203.
[18]HEIDERSCHEIT B C,HOERTH D M,CHUMANOV E S,etal.Identifying the time of occurrence of a hamstring strain injury during treadmill running:a case study[J].Clin Biomech,2005,(20):1072-1078.
[19]HUNTER J P,MARSHALL R N,MCNNAIR P J.Segment-interaction analysis of the stance limb in sprint running[J].J Biomech,2004,37(9):1439-1446.
[20]LIEBER R L,FRIDEN J.Muscle damage is not a function of muscle force but active muscle strain[J].J Appl Physiol,1993,74(2):520-6.
[21]LIEBER R L,FRIDEN J.Mechanisms of muscle injury gleaned from animal models[J].Am J Phys Med Rehabil,2002,81(11):S70-9.
[22]MAGNUSSON S,AAGAARD P,SIMONSEN E,etal.Passive tensile stress and energy of the human hamstring muscles in vivo[J].Scand J Med Sci Sports,2000,10:351-359.
[23]NIEMINEN H,NIEMI J,TAKALA E P,etal.Load-sharing patterns in the shoulder during isometric flexion tasks[J].J Biomech,1996,28:555-566.
[24]ORCHARD J.Intrinsic and extrinsic risk factors for muscle strains in Australian footballers[J].Am J Sports Med,2002:300-303.
[25]PRILUTSKY B I,ZATSIORSKY V M.Optimization-based models of muscle coordination[J].Exe Sport Sci Rev,2002,(30):32-38.
[26]RICHARD A,BRAND DOUGLAS R,PEDERSEN,etal.The sensitivity of muscle force predictions to changes in physiologic cross-sectional Area[J].J Biomech,1986,19(8):589-596.
[27]SCHACHE A G,WRIGLEY T V,BAKER R,etal.Biomechanical response to hamstring muscle strain injury[J].Gait Posture,2009,(29):332-338.
[28]SIMONSEN E B,THOMSEN L,KLAUSEN K.Activity of mono- and biarticular leg muscles during sprint running[J]. Eur J Appl Physiol Occup Physiol,1985,54(5):524-32.
[29]SOFIA HEINTZ.Muscular forces from static optimization[A].Technical Reports from Royal Institute of Technology[R].KTH Mechanics SE-100 44 Stockholm,Sweden,2006:37-40.
[30]THELEN D G,CHUMANOV E S,HOERTH D M,etal.Hamstring muscle kinematics during treatmill sprinting[J].Med Sci Sports Exe,2005,37(1):108-14.
[31]TRANSFELDT E,ROBERTSON D,BRADFORD D.Ligaments of the lumbosacral spine and their role in possible extraforaminal spinal nerve entrapment and tethering[J].J Spinal Disord,2001,(6):507-512.
[32]WOOD G.Biomechanical limitations to sprint running[J].Med Sports Sci,1987,(25):58-71.
[33]WOODS C,HAWKINS R D,MALTBY S.The football association medical research programme:An audit of injuries in professional football-analysis of hamstring injuries[J].Br J Sports Med,2004,38(1):36-41.
[34]YU B,QUEEN R M,ABBEY A N,etal.Hamstring muscle kinematics and activation during overground sprinting[J].J Biomech,2008,41(15):3121-3126.
BiomechanicalAnalysisofWorkCharacteristicsandStrainRisksofBi-articularMusclesinLowerExtremityduringSprinting
ZHONG Yun-jian1,2,LIU Yu2,FU Wei-jie2,WEI Shu-tao2,WU Xie2,LI Qin3
Objective:The aim of this study was to establish models regarding the muscle functions and musculoskeletal system in lower extremity,quantify the changes in muscle length,contraction velocity,muscle stress,muscle power,and total work of the bi-articular muscles during sprinting and analyze the biomechanical characteristics of bi-articular muscle work and strain risks.Methods:the kinematic and kinetic data from elite sprinters were collected.Two models of the muscle functions and musculoskeletal system in lower extremity were built to calculate the changes in the parameters of muscle function in lower extremity.Furthermore,the muscle stress,the power,and the total amount of work done per unit area of a single muscle were also examined combined with the quantification via optimization algorithm.Results and Conclusion:the peak values of muscle stress,muscle length,contraction velocity,and negative muscle power per unit area in bi-articular muscles were significantly greater than other mono-articular muscles.During the early swing of the stride cycle,the gastrocnemius were eccentric contracted to counteract the loads from external forces,and thus reached a maximal negative power.These results indicated that the gastrocnemius were susceptible to injury during the early stance.For the hamstrings,muscle length and muscle stress were higher during the late swing and early stance phases.Besides,the hamstrings performed quick negative work and produced a maximal power during mid-swing.Hence,these periods were identified as the potential phases for hamstring muscle strain injury.The quadriceps performed much negative work and reached a maximal power during the early swing of the stride cycle,whereas the quadriceps were at a high risk of injury in this period of the gait.The negative work done in bi-articular muscles was significantly higher than other mono-articular muscles during a full gait circle.Additionally,during certain phases of gait,the positive and negative work were simultaneously done with bi-articular muscles on two separated joints.This study provided the biomechanical basis for the strain risk of bi-articular muscle during sprinting through a series of related indicators of the mechanical load on single muscle.
sprinting;bi-articularmuscle;strainrisk;biomechanics
1000-677X(2014)11-0060-07
2014-05-04;
:2014-10-12
国家自然科学基金资助项目(30871210)。
钟运健(1976-),男,江西兴国人,副教授,博士,主要研究方向为神经动作控制的生物力学机制,Tel:(0791)83838791,E-mail:zhong7019442@163.com;刘宇(1959-),男,河北张家口人,教授,博士,博士研究生导师,主要研究方向为神经动作控制的生物力学机制和体育工程,Tel:(021)51253239,E-mail:yuliu@sus.edu.cn。
1.南昌大学 教育学院 体育系,江西 南昌 330031; 2.上海体育学院 运动科学学院,上海 200438;3.清华大学 体育部,北京 100084 1.Nanchang University,Nanchang 330031,China;2.Shanghai University of Sport,Shanghai 200438,China;3.Tsinghua University,Beijing 100084,China.
G804.6
:A
