不同食用菌品种产漆酶规律的研究*
2014-07-18孙淑静郭艳艳吴晓华单书凯李念静翁彩红张燎原胡开辉
孙淑静,郭艳艳,吴晓华,单书凯,李念静,翁彩红,张燎原,胡开辉
(福建农林大学生命科学学院,福建 福州 350002)
不同食用菌品种产漆酶规律的研究*
孙淑静,郭艳艳,吴晓华,单书凯,李念静,翁彩红,张燎原,胡开辉**
(福建农林大学生命科学学院,福建 福州 350002)
本研究对6种食用菌在液体和栽培料培养时产漆酶能力和产酶规律进行了比较分析。实验结果表明,在液体培养条件下,凤尾菇、秀珍菇和鸡腿菇均在第6天左右达到酶活性峰值,金针菇酶活性持续增加,在第10天达到峰值。在栽培料培养条件下,凤尾菇、秀珍菇和鸡腿菇均在大试管中长至一半时达到酶活性峰值,金针菇酶活性持续增加,在原基形成时达到峰值。在2种培养条件下,生长速度较快的凤尾菇、秀珍菇、金针菇fl98、鸡腿菇漆酶活性均较高,而生长速度慢的白玉HM03和滑菇漆酶活性较低,漆酶产生规律相同,产漆酶能力较强的菌种菌丝生长较快,产酶高峰多出现较早,而产漆酶能力较弱的菌种菌丝生长较慢。以上结果说明食用菌中漆酶在菌丝生长速度上起着重要的作用,可为以后食用菌提高其生长速度、定向育种提供重要的参考。
食用菌;生长速度;漆酶
漆酶(Laccase)是1种含铜的多酚氧化酶,属蓝色多铜氧化酶家族[1],能够催化酚类、羧酸、生物色素、芳胺类、甾体类激素、金属有机化合物等物质生成醌类化合物、羰基化合物和水[2]。近几年的研究表明,漆酶在纸浆的生物漂白[3,4]、新型传感器的研制和生物检测[5,6]、环境污染物的降解[7,8]、生物燃料电池的阴极反应[9]、食品饮料中酚类物质的去除[10]等领域具有潜在的应用价值。研究还发现很多食用菌菌体中富含漆酶, 它有利于呼吸过程中电子传递的正常运行,呼吸过程正常运转,为菌体的生命活动提供充足的可利用的能量,从而加速菌体的生长发育[11]。此外,在菌丝体生长发育过程中,漆酶在分解木质素的同时还会产生酚类或醌类化合物,而酚类或醌类物质是有毒物质,是1种很好的杀虫剂,能够抑制杂菌的生长,从而防止杂菌进一步污染,并影响菇类菌体的生长发育[12]。由此可见,漆酶参与真菌体内的许多生物化学反应,具有重要的生理功能[13],影响着真菌的发育及形态的发生[14]。但食用菌在不同条件下菌丝产漆酶规律的相关性研究甚少。针对目前有些珍贵食用菌生产周期长、生产成本高的实际情况,如何缩短食用菌生产周期及提高抗杂菌能力、降低成本,成为目前食用菌育种亟待解决的关键问题之一。本实验通过测定不同条件下不同食用菌菌丝生长速度、漆酶酶活及产酶规律,分析其相关性,从而为食用菌根据漆酶定向选育高产性能的新品种提供理论依据。
1 材料与方法
1.1 材料
1.1.1 供试菌株
凤尾菇(Pleurotussajor-caju)、金针菇fl98(Flammulinavelutipes)、秀珍菇(Pleurotusgeesteranus)、鸡腿菇(Coprinuscomatus)、滑菇(Pholiotanamek)、白玉HM03(Hypsizigusmarmoreus),均为实验室保藏菌种。
1.1.2 供试培养基
PDA综合培养基:马铃薯200 g、葡萄糖20 g、蛋白胨10 g、KH2PO43 g、MgSO41.5 g、VB10.01 g、琼脂20 g,加水至 1 000 mL,pH自然;液体加富培养基:马铃薯200 g、葡萄糖20 g、酵母粉3 g、蛋白胨3 g、KH2PO41 g、MgSO40.5 g、VB10.1 g,加水至 1 000 mL,pH自然;生产培养基:棉籽壳71%、麦麸23%、石灰1%、玉米粉5%,含水量60%~65%。
1.2 方法
1.2.1 菌株生长速度测定试验
将凤尾菇、金针菇fl98、秀珍菇、鸡腿菇、滑菇、白玉HM03六种菌株分别接种于PDA培养基,每个菌株做3个重复,24℃恒温培养箱中培养10 d,每隔1 d测其菌丝生长直径。
根据生产培养基配方,将栽培料装入21 mm×180 mm试管中,每个菌种装料10管,121℃高压灭菌2 h,冷却后以5%的比例接入原种,一定时间后测其菌丝生长速度。
1.2.2 粗酶液的提取
(1)液体加富培养基中酶液的提取
将待测菌株接入含有50 mL液体加富培养基的三角瓶中,每瓶6片(直径2 cm),每个菌株做3个重复,110 r·min-1,24℃恒温振荡培养10 d,每隔1 d提取1次发酵液,经纱布过滤,离心后取其上清液,即为粗酶液,4℃冰箱保存备用[15-17]。
(2) 生产培养基中酶液的提取
分4个阶段取样:菌丝生长过程(平均取5次)、菌丝满袋、原基形成、幼菇形成。每1次取样分别取若干栽培料打碎、混匀,四分法取3 g加30 mL蒸馏水,24℃浸提4 h,过滤后离心,取其上清液即为粗酶液,4℃冰箱保存备用[18]。
1.2.3 漆酶酶活性测定
以ABTS为底物,3 mL反应体系中含有醋酸-醋酸钠缓冲液(pH 5.0)2.7 mL、粗酶液0.1 mL和1 mmol·L-1的ABTS 0.2 mL,25℃水浴5 min,测其在420 nm处反应开始前3 min内吸光值的增加。将粗酶液在沸水中煮沸15 min作为对照。定义每分钟使吸光值增加0.1所需的酶量为1个酶活力单位(U)[19,20]。
2 结果与分析
2.1 不同菌株生长速度测定
不同菌株菌丝生长速度情况见表1和图1。
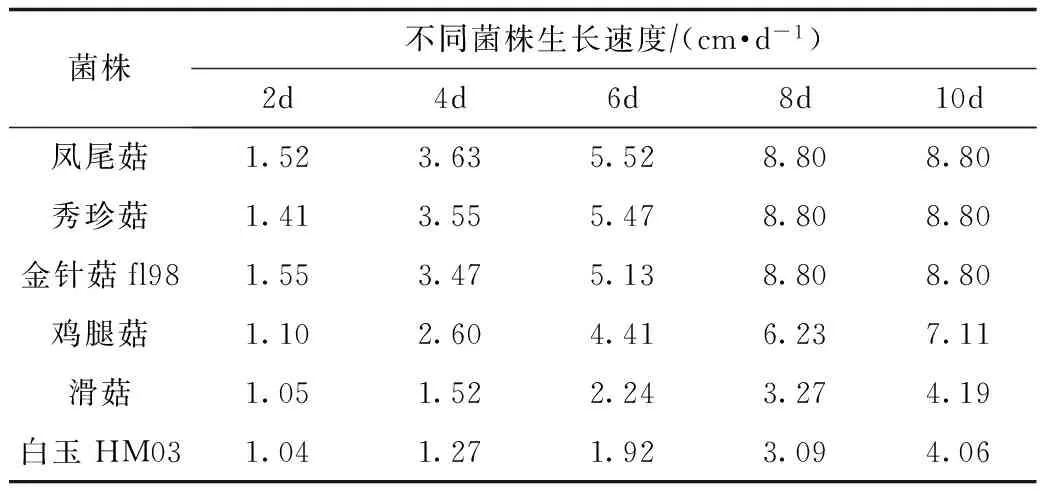
表1 不同菌株在PDA平板上的生长速度比较
由表1可知,不同菌株在PDA平板上生长速度差异较大。凤尾菇、秀珍菇、金针菇fl98生长速度较快,8 d长满平皿(平皿直径8.8 cm);而鸡腿菇生长速度较慢,13 d长满平皿;滑菇和白玉HM03生长速度最慢,长满平板需20 d左右。

由图1可见,在栽培料中凤尾菇、秀珍菇生长速度最快,金针菇fl98次之,鸡腿菇较慢,滑菇和白玉HM03最慢,与PDA平板上的结果基本一致。
2.2 不同菌株在液体培养基中漆酶变化规律
不同菌株在液体培养基中漆酶酶活变化见图2。

由图2可知,凤尾菇在生长过程中酶活性一直很高,在第4天达到酶活性的峰值;秀珍菇和鸡腿菇酶活性也较高,在第6天达到峰值。凤尾菇、秀珍菇和鸡腿菇的产酶规律都是生长前期酶活性较低,随着培养时间增加,酶活性逐渐达到峰值,接着降低,在生长后期又略有增加。滑菇、白玉HM03前期酶活较低,后期略有增加。金针菇fl98漆酶活性持续增加,在生长后期达到酶活性的峰值。相关文献表明,漆酶参与菌体的呼吸作用,为菌体的生命活动提供更多的可利用的能量,从而加速菇类的生长发育过程[11]。本试验选取6种不同生长速度的菌种对其进行液体培养,在生长过程中测其漆酶活性,以探讨漆酶活性与菌丝生长速度的关系。
2.3 不同菌株在栽培料中漆酶变化规律
不同菌株在栽培料中漆酶酶活变化见图3。
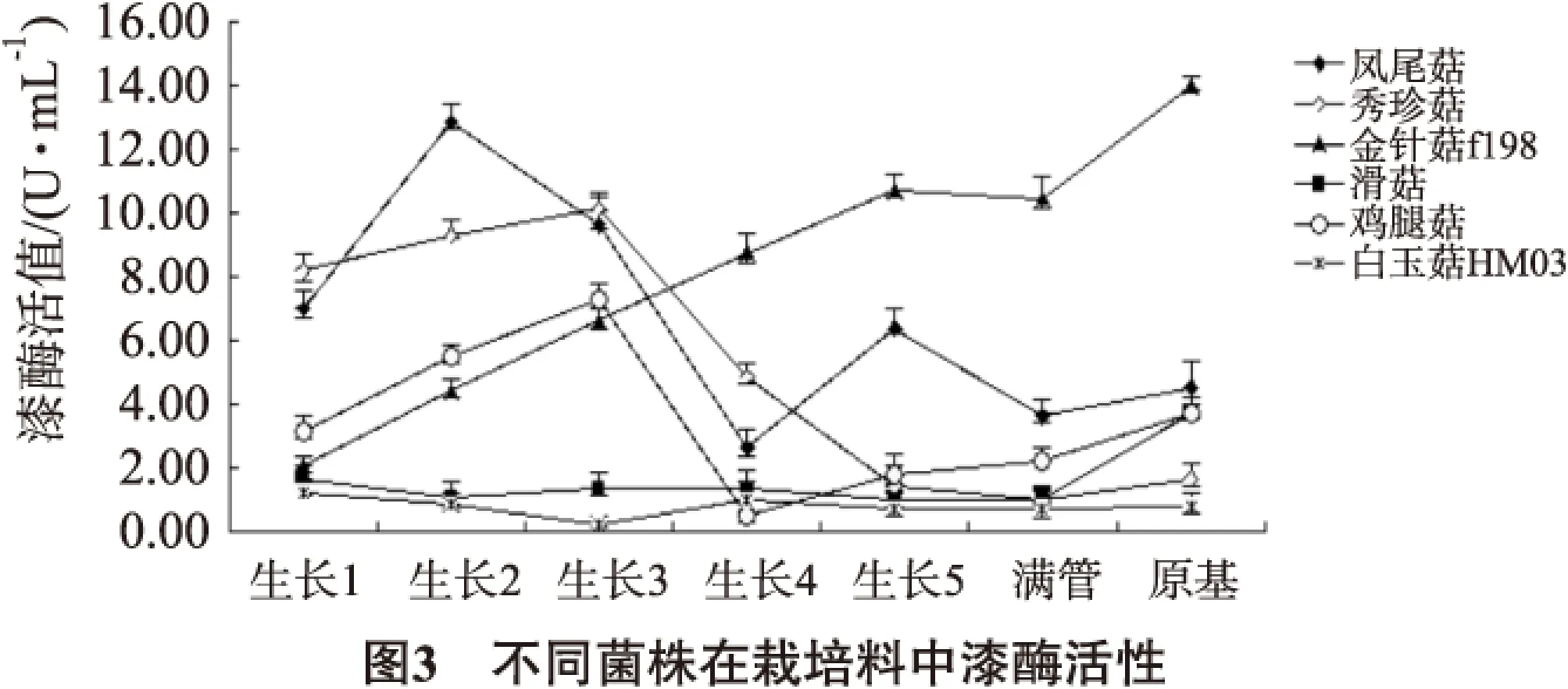
金针菇生长过程中酶活性持续增加,在原基形成时达到峰值,且酶活性高于凤尾菇;凤尾菇漆酶活性很高,且在其生长前期酶活性达到峰值,之后持续降低,在生长后期略有增高;秀珍菇酶活性也较高,在菌丝生长至半管时达到酶活性峰值,之后一直下降,后期亦略有增加;鸡腿菇酶活性略低于凤尾菇和秀珍菇,在菌丝生长至半管时也达到酶活性峰值,在形成原基时酶活性也较高;滑菇酶活性相对较低,生长前期酶活性也较低,在原基形成时酶活性最高;白玉HM03生长过程中酶活性一直较低。
3 小结与讨论
在液体培养基中,金针菇的漆酶酶活性持续增加,在第10天达到峰值;栽培料中,金针菇在菌丝生长阶段,漆酶活性也较高,这一规律与相关报道结论一致[21,22]。栽培料中,凤尾菇、秀珍菇、鸡腿菇的漆酶活性均随栽培时间的延长由高逐渐降低,这与金顶侧耳(Pleurotuscitrinopileatus)[23]、巴西蘑菇(Agaricusblazei)[24]等食用菌的情况相似;液体培养时,凤尾菇、秀珍菇的漆酶活性较高,鸡腿菇的漆酶活性也相对较高,这与其生长速度的测定结果相一致。结合不同菌种的生长速度和酶活性的测定结果,可以得到,生长速度较快的金针菇 fl98、凤尾菇和秀珍菇,其漆酶酶活性亦较高;生长速度中等的鸡腿菇,其酶活性相对较高;而生长速度比较慢的滑菇、白玉 HM03,几乎没有酶活性。以上结果说明食用菌中漆酶在菌丝生长速度方面起着重要的作用,可为以后食用菌提高其生长速度、定向育种提供重要的参考。
[1]钞亚鹏,钱世钧. 真菌漆酶及其应用[J].生物工程进展,2001,21(5):23-27.
[2]吴晴. 白腐真菌及其漆酶的应用研究[J].安徽科技, 2010(3):47-48.
[3]Nurdan KP, Merih S, Azmi T.Laccase production by trametes versicolor and application to denim washing[J].Process Biochemistry, 2005, 40(5): 1673-1678.
[4]Sigoillot C, Record E, Belle V, et al.Natural and recombinant fungal laccases for paper pulp bleaching[J].Appl Microbiol Biotechnol, 2004, 64(3): 346-352.
[5]Gomes SASS, Nogueira JMF, Rebelo MJF. An amperometric biosensor for polyphenolic compounds in red wine[J].Biosensors and Bioelectronics, 2004, 20(6): 1211-1216.
[6]Fabio V, Antonio C, Santa R, et al. A high sensitivity amperometric biosensor using a monomolecular layer of laccase as biorecognition element[J]. Biosensors and Bioelectronics, 2004, 20(2): 315-321.
[7]Atef J, Francisco G, Michel JP, et al. Role ofPycnoporuscoccineuslaccase in the degradation of aromatic compounds in olive oil mill wastewater[J]. Enzyme and Microbial Technology, 2005, 36(4): 478-486.
[8]Rigas F, Dritsa V, Marchant R, et al. Biodegradation of lindane byPleurotusostreatusvia central composite design[J]. Environment International, 2005, 31(2): 191-196.
[9]Katz E, Filanovsky B, Willner I. A biofuel cell based on two immiscible solvents and glucose oxidase and microper oxidase monolayer functionlized electrodes[J]. New J Chem, 1999(23): 481-487.
[10]Servili M, De Stefano G, Piacquadio P, et al. A novel method for removing phenols from grape must[J]. Am J Enol Viticulture, 2000(51): 357-361.
[11]Turner EM, Wright M, Ward T, et al. Production of ethylene and other volatiles and changes in cellulase and laccase activities during the life cycle of the cultivated mushroom,Agaricusbisporus[J]. Gen Microbiol, 1975, 91(1): 167-176.
[12]Bollag JM, Shuttleworth KL, Anderson DH. Laccase-mediated detoxification of phenolic compounds[J]. Appl Environ Microbiol, 1988, 54(12): 3086-3091.
[13]Bourbonnais R, Paice MG. Oxidation of nonphenolic substrates: An expanded role for laccase in lignin biodegradation[J]. Febs Letters, 1990, 267(1): 99-102.
[14]Leatham G, Slahanman M A. Studies on the laccase ofLentinusedodes: specificity, localization and association with the development of fruiting bodies[J]. Gen Microbiol, 1981, 125(1): 147-157.
[15]兰瑞芳,林少琴,林玉满,等. 杏鲍菇漆酶的生物学特性[J]. 食用菌学报,2002,9(2):14-16.
[16]俞苓,刘民胜,陈有容. 杏鲍菇液体培养中胞外酶活性变化[J]. 食用菌,2003(1):7-9.
[17]王宜磊, 毕红卫. 多孔菌漆酶性质研究[J]. 淄博学院学报:自然科学与工程版,2002,4(4): 12-14.
[18]刘朝贵, 邵坤, 聂和平,等. 不同培养料对鸡腿菇胞外酶活性影响的研究[J]. 西南师范大学学报:自然科学版,2008,33(1):40-42.
[19]朱海潇,黄桂英,胡开辉,等. 凤尾菇漆酶性质及应用的研究[J]. 福建农业报,2008,23(1):48-52.
[20]堵国成, 赵政, 陈坚. 真菌漆酶的活性测定及其在织物染料生物脱色中的应用[J]. 江南大学学报, 2003,2(1): 83-90.
[21]刘苹, 唐志红, 李梦玉, 等. 金针菇漆酶活性测定的最佳反应条件及液体培养胞外酶的研究[J]. 食品科技, 2012, 37(6):35-36.
[22]冯伟林, 蔡为明, 金群力,等. 金针菇生长发育期间相关胞外酶的活性变化研究[J]. 浙江农业学报, 2012, 24(3): 430-433.
[23]李娟, 倪新江, 樊守金, 等. 金顶侧耳不同生长期几种胞外酶活性变化[J]. 吉林农业大学学报, 2006,28(6):24-26.
[24]杨立红,倪新江,黄清荣. 栽培基质对巴西蘑菇胞外酶活性的影响[J]. 吉林农业大学学报,2004,26(3):284-286.
Study on the Production Rules of Laccase of Different Edible Fungi
SUN Shu-jing, GUO Yan-yan, WU Xiao-hua, SHAN Shu-kai,LI Nian-jing, WENG Cai-hong, ZHANG Liao-yuan, HU Kai-hui
(College of Life Sciences, Fujian Agriculture and Forestry University, FuzhouFujian350002)
The capacities and production rules of laccase by six different types of edible fungi were analyzed and compared in two media. The results showed the laccase activities ofPleurotussajor-caju,PleurotusgeesteranusandCoprinuscomatusreached peak at 6th day in liquid medium. The laccase activity ofFlammulinavelutipesincreased continuously and reached the peak at 10th day. The laccase activities ofPleurotussajor-caju,PleurotusgeesteranusandCoprinuscomatusreached the peak when the mycelia grew up the half part of big test tube in cultivation material. The laccase activity ofFlammulinavelutipesincreased continuously and reached the peak at the period of anlage formed. Under two culture conditions,Pleurotussajorcaju,Pleurotusgeesteranus,FlammulinavelutipesandCoprinuscomatushad high laccase activities with short growth cycle, whileHypsizigusmarmoreusandPholiotanamekhad little laccase activity with long growth cycle. The production rules of laccase were same under two culture conditions, the strains with high production capacity of laccase grew fast and produced laccase at early stage of growth cycle, and the strains with weak production capacity of laccase grew slowly. The fruiting test results indicated that the laccase played an important role in mycelial growth of the edible fungi, and it can provide important reference for growth rate and directional breeding.
Edible fungi; Growth rate; Laccase
*项目来源:福建省发改委农业五新工程项目“珍稀食用菌真姬菇新品种闽真1号工厂化栽培技术熟化与中试研究”,项目编号:闽发改委投资[2012]931号;福建省教育厅项目“双重靶向性筛选食用菌新品种体系的建立”,项目编号:JA11079;福建省发改委生物产业技术专项“福建省生物产业技术专项:真姬菇品种改良与农业可再生资源适应性研究及示范”,项目编号:2011878;福建省高校产学合作科技重大项目“真姬菇(闽真2号)菌包成熟度多尺度生化参数指标”,项目编号:2013N5101。
孙淑静(1978-),女,副教授,主要从事食用真菌学研究。E-mail:shjsun2004@126.com
**通信作者: 胡开辉,男,教授。E-mail:hukh62@sohu.com
2014-03-25
S646.9
A
1003-8310(2014)03-0038-03
