不同类型雄性不育苎麻细胞学机理研究
2014-06-08薛丽君邢虎成马艳玲
薛丽君,邢虎成,2*,马艳玲
(1.湖南农业大学苎麻研究所,湖南 长沙 410128;2.湖南省种质资源创新与资源利用重点实验室,湖南 长沙410128)
自20世纪初以来,在各种作物上已先后发现了雄性不育现象。利用雄性不育已经生产了水稻、棉花、油菜、甘蓝等作物的杂交种[1–4]。中国苎麻雄性不育研究始于20世纪60年代[5]。目前,发现有2种雄性不育类型:①由细胞核控制的雄性不育[6];②核–质互作雄性不育[7]。
刘飞虎等[8]报道了蛋白质和氨基酸等的差异可能是导致苎麻雄性不育的原因。宋军等[9–10]报道物质 (可溶性糖、淀粉、可溶性蛋白质和游离脯氨酸)代谢和同工酶代谢差异是不育系、恢复系之间的主要差异。段继强等[11]报道不育系atpA 基因的序列变异和(或)异常表达可能与苎麻细胞质雄性不育关系密切。
笔者所在课题组对黑皮蔸MS系列和大叶绿MS系列等21份雄性不育种质进行了田间观察、花粉活力测定、自交种子发芽试验和天然杂交种子发芽试验,结果将湖南农业大学苎麻研究所收集的21份苎麻种质归为4类,即雄花萎缩型、雄花不开裂型、花粉不育型、待定型[12]。在此基础上,笔者对雄花萎缩型、雄花不开裂型、花粉不育型雄性不育株的花芽分化和花药发育过程进行观察,并与可育材料进行比较,分析不育系的花药败育时期和发育特点,旨在为探索苎麻雄性不育的机理及今后将苎麻不育种质应用于生产实践提供参考数据。
1 材料与方法
1.1 供试苎麻
雄性不育材料雄花萎缩型种质圆青5号、花被不开裂型种质弯子苎麻和花粉不育型种质黄小叶。雄性可育材料中苎1号。以上材料均来自湖南农业大学耘园试验基地,栽培时间为3年以上,每种质1小区,不设重复,常规管理。
1.2 方 法
1.2.1 取样与固定
从孕蕾期开始取样,直到开花为止,放于青霉素小瓶中,用FAA(50%乙醇89 mL+6 mL冰醋酸+5 mL福尔马林+5 mL甘油)固定,于4 ℃冰箱中保存。每次取样记录花芽大小、取样时间并给样本编号(表1)。由于不同品种苎麻开花时间不同,因此本试验是以花芽长度作为取样的标准。
1.2.2 石蜡切片观察
固定的材料采用常规石蜡切片法制成永久切片,切片厚度9 μm,番红–固绿染色,中性树胶封片,在奥林巴斯BX51型光学显微镜下观察并拍照。

表1 取样时间及花芽长度Table 1 Sampling time and flower bud size
2 结果与分析
2.1 雄性可育中苎1号雄花的形成与发育
雄花花芽由茎下部叶腋的副芽形成。雄花花芽分化分为花序分化期、花被形成期、雌雄蕊形成期和雌雄蕊分化期。花序分化时,副芽顶端细胞分裂加快,使尖端横径加宽,形成花序原始体,它可分化出花序轴和许多分支轴,在花轴分支的周围会分化出5~9个小突体,即为雄花的原始体(图1–A)。继续分化,花的原始体继续膨大,其基部产生花被原基突起,逐渐向花的顶部伸长,形成花蕊的一种保护机构(图1–B)。当花被伸长并向顶部包围时,呈圆锥状的花原基中央突起就是雌、雄原始体,以后圆锥状的中央突起形成雌蕊原始体,而圆锥状的基部出现4个小突起,即为雄蕊原始体(图1–C)。苎麻雄花中的雌蕊珠心原始体,并不随着整个花的发育继续分化,而是在进一步的发育过程中逐渐萎缩消失,因此成熟后子房壁内只是一个空腔(图1–D)。
雄蕊原基最初呈小丘状,随后伸长成棒状,外形上并没有花药和花丝的变化。继续发育,花药呈现4个或2个瓣状结构,每瓣发育成1个花粉囊(图1–E)。从纵切面看,花药内部已发生分化,外面为表皮层,花药里面为孢原细胞,细胞质较浓,染色较深(图1–F)。靠中轴的几列细胞,是药隔的原始细胞,在花粉粒之间组成药隔,内具维管束,被染成红色(图11–G)。花药分化,最外层是表皮细胞,表皮内的孢原细胞进行平周分裂,产生中层、绒毡层和造孢细胞3层。造孢细胞通过几次有丝分裂形成数量较多的花粉母细胞,再经减数分裂成为四分体。当花粉母细胞进行减数分裂时,中层细胞会逐渐消失,绒毡层的细胞逐渐长大,四分体形成不久后,花粉母细胞的壁开始破裂,花粉粒从中释放出来,绒毡层细胞溶解(图1–H)。花药成熟后,花粉囊壁仅为一层表皮,其余各层消失,囊中分散的花粉粒即散发出去。
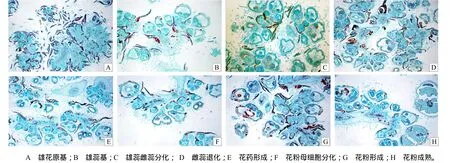
图1 中苎1号花芽发育情况(×40)Fig.1 Development of flower bud of Zhongzhu No.1 (×40)
2.2 雄花萎缩型园青5号雄花的形成与发育
雄花萎缩型园青5号花芽长至0.3 cm时,已发育到雌雄蕊分化期(图2–A),而可育中苎1号的花芽长至0.4 cm时还处于花序分化期,可见,雄花萎缩型苎麻品种的花芽发育较快。雌蕊原始体形成退化,即雌蕊在发育过程中会退化消失,形成空腔(图2–B),这与可育系材料中苎1号的相同。
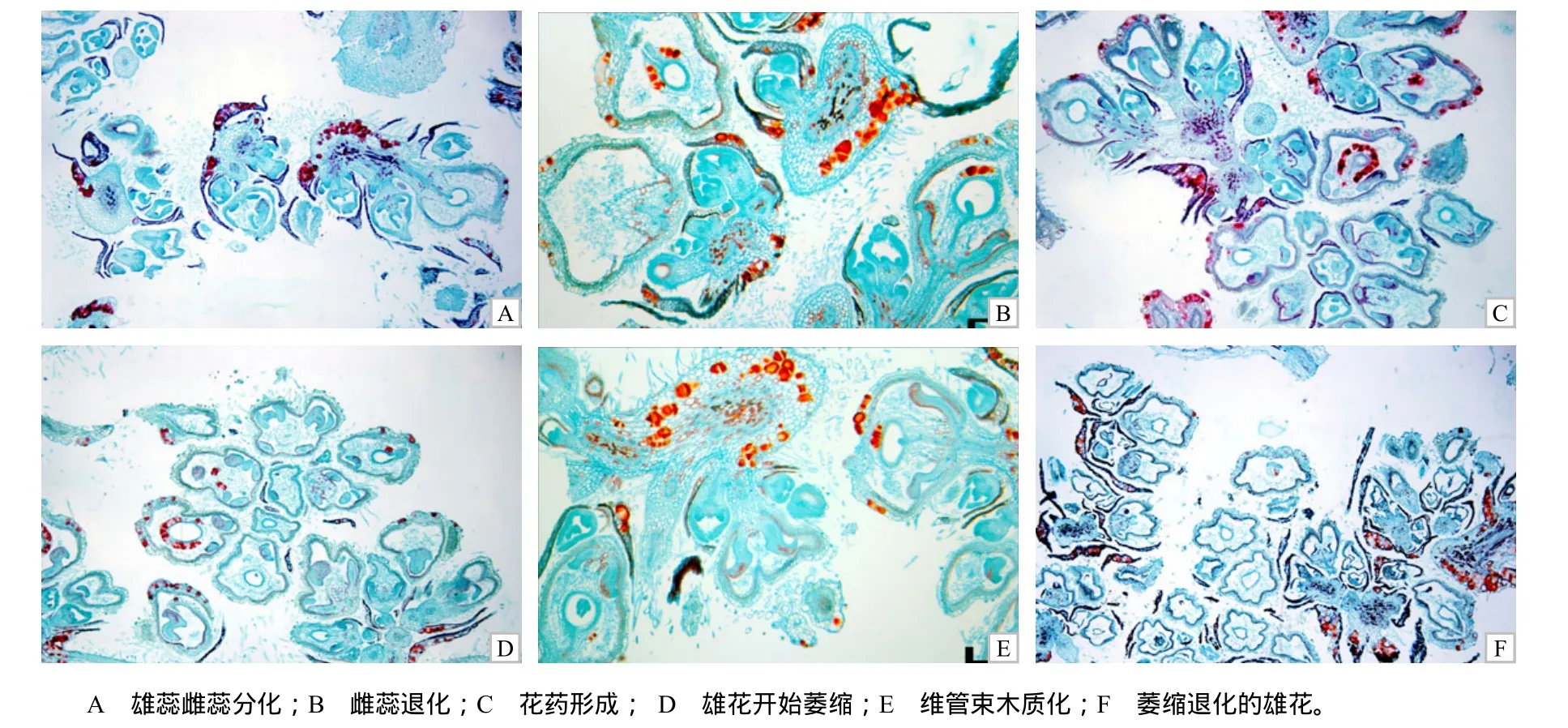
图2 雄花萎缩型园青5号苎麻花芽发育情况(×40)Fig.2 Flower bud development for the male flowers atrophic type Yuanqing No. 5 ramie (×40)
雄花萎缩型园青5号在花芽长至0.8 cm时,雌蕊、雄蕊都在正常发育,雄花已有花药和花丝的变化(图2–B);花芽在分化出花丝和花药后就停止生长或生长极其缓慢,花芽长至1.4~1.5 cm,花粉囊形成开始萎缩变形(图2–C),花药染色逐渐变浅(图2–D),由于固绿主要用于细胞质的染色,说明孢原细胞的细胞质在变稀;花药和花丝的表皮细胞位于花药边缘(图2–E)。最后花药萎缩干瘪,花粉囊逐渐消失,形成中空的结构,甚至消失而失去育性(图2–F)。
2.3 花被不开裂型弯子苎麻雄花的形成与发育
花被不开裂型弯子苎麻花芽发育较中苎1号快,其花芽长至0.3 cm时雌、雄蕊已经分化,花药内部已发生分化,表皮下的孢原细胞细胞质较浓、染色较深,有些花药中可看到已有花粉囊形成,靠中轴的药隔原始细胞清晰可见,其中维管束细胞被染成红色(图3–A)。
当花芽长至0.7 cm时,花粉囊形成,花被内侧变黑,花被外侧有木质化现象并被染成红色,这种花被的木质化现象会导致在花粉成熟时花被无法开裂而造成雄性不育;雌蕊退化消失成空腔,这与中苎1号的相同,但花被不开裂型弯子苎麻退化的雌蕊会逐渐木质化而被染成红色(图3–B);有些花药发育畸形或者萎缩干瘪,未形成瓣状结构(图3–C),药隔维管束不是位于中轴且面积较大(图3–D);有些花粉囊中产生的花粉粒会逐渐变黑(图3–E)。当花芽长至4.5 cm时,一些雄花花被部分木质化而导致不能开裂散粉(图 3–F)。

图3 花被不开裂型弯子苎麻的花芽发育情况(×40)Fig.3 Flower bud development for male indehiscent type Wanzi ramie (×40)
2.4 花粉不育型黄小叶雄花的形成与发育
花粉不育型黄小叶花芽长至0.3 cm时已完成雄花花芽的分化,花药中可看到花粉囊、维管束和药隔。可见,花粉不育型黄小叶的花芽发育也比中苎1号的快。此时花药呈瓣状结构,有2个花粉囊(图4–A)。孢原细胞细胞质较浓,染色深,药隔中维管束细胞被染成红色(图4–A)。花芽长至0.7 cm时花粉囊形成,花粉囊中已有花粉粒生成(图4–B),随后持续在这一阶段;当花芽长至2.3 cm时,药隔消失,花粉囊中的花粉粒颜色逐渐变深(图4–C);花芽长至3.8 cm时,花丝发育异常,花粉囊中花粉异常且数量很少(图4–D)。黄小叶在形成花粉后,花粉囊壁直到花芽长至6.0 cm时都未破裂(图4–E、F)。
从图5可见,在黄小叶部分花药发育过程中,从造孢母细胞时期开始出现花丝发育的异常膨大(图5–A),尤其是花丝与花药连接处发生弯曲(图5–B、C),这可能导致黄小叶花粉发育不良而不育。小孢子母细胞时期花药的表皮、中层细胞和绒毡层细胞都存在分解现象(图5–D)。在小孢子二分体时期,花药内部的绒毡层细胞出现降解(图5–E)。在四分体时期也可以看到花药内壁绒毡层细胞开始降解(图5–F),因此,推测黄小叶苎麻花粉不育是由于绒毡层和花丝发育异常导致。

图5 黄小叶花丝和绒毡层的切片观察结果Fig.5 Filaments and tapetum layer of Huangxiaoye
3 讨 论
不同雄性不育材料的败育时期不尽相同,从雄蕊分化到花粉粒成熟的各个时期均有可能[13–16]。花芽分化经历苞片原基分化期、萼片原基分化期、花瓣原基分化期、雄蕊原基分化期、雌蕊原基分化期、雄蕊瓣化期和雌蕊瓣化期[17–18],而花粉发育经历孢原细胞、造孢细胞、小孢子母细胞、四分体、单核小孢子、花粉等阶段,在这一发育过程中任何一个阶段出现异常都将产生败育花粉,最终导致雄性不育。笔者研究的3个雄性不育苎麻败育的时期都发生在花药花丝形成以后,即在花粉发育阶段。这与小麦[13]、亚麻[14]、萝卜[15]、梨[16]等雄性不育材料败育的时期相似。但熊少兵等[19]的研究发现苎麻雄性不育系GS14–1雄蕊发育受阻于雌雄蕊原基分化期或花药形成初期,可能与取样时间和花的大小有关。
绒毡层细胞为小孢子发育提供营养物质[20],并在减数分裂后适时分泌胼胝质,使小孢子分离。绒毡层提前解体而无法给小孢子的发育提供营养物质,会导致花粉失去育性,而在减数分裂后,绒毡层没有适时的分泌胼胝质使小孢子分离,也会导致花粉败育[21]。本试验中花粉不育型黄小叶小孢子败育可能是花药发育过程中绒毡层异常导致的。另外在开花时,花被上部展开,花丝弹直,将药室弹开,花粉粒外扬,在试验中看到雄性不育苎麻黄小叶中花丝发育异常,这是否会导致苎麻雄性不育需要进一步研究。
植物雄性不育系花药中的维管束出现异常现象,在番茄、小麦、高粱、水稻等作物中均有报道,并认为药隔维管束出现的异常现象与花粉的败育有一定关系。此外,维管束是植物体的输导组织,如发育不良则直接影响物质的交换和运转,从而影响小孢子的发育[22]。在本试验中,观察到花被不开裂型弯子苎麻的维管组织出现木质化现象,可以认为维管组织的异常现象是造成花粉败育的原因之一。雄花萎缩型园青5号的败育也可能与维管束异常有关。维管束异常可能导致花药在发育过程中缺乏某种物质而发生畸变。
本研究从形态学方面阐述了苎麻雄性不育材料雄花花芽的发育特点,要进一步探索苎麻雄性不育的细胞学机理则要从绒毡层、细胞核、减数分裂、四分体等方面做更细致的观察。
[1]朱四元,陈金湘.棉花雄性不育系的选育及不育机理的研究进展[J].江西棉花,2004,26(1):11–16.
[2]徐一兰,唐海明,官春云.油菜细胞质雄性不育的分子生物学及杂种优势利用研究进展[J].作物研究,2006,20(5):446–452.
[3]章清杞,张艳华.水稻雄性不育及不育基因定位研究进展[J].亚热带农业研究,2009,5(2):78–83.
[4]方智远,孙培田,刘玉梅,等.甘蓝显性雄性不育系的选育及其利用[J].园艺学报,1997,24(3):249–254.
[5]吴刚,孙焕良,伍波,等.苎麻不育特性的研究现状与展望[J].作物研究,2007,87(S1):690–692.
[6]罗来尧.苎麻杂种优势利用的初步研究[J].中国麻作,1984(3):23–27.
[7]刘飞虎,梁雪妮,张寿文,等.苎麻雄性不育系育性鉴定和遗传分析[J].中国麻作,2000,22(1):7–10.
[8]刘飞虎,梁雪妮.苎麻雄性不育系生理生化特点初步研究[J].中国麻作,2000,22(3):17–21.
[9]宋军,张中华,丁明忠,等.苎麻雄性不育材料的两种同工酶初步分析[J].中国麻业科学,2008,30(3):141–143.
[10]宋军,张中华,潘光堂.苎麻雄性不育材料的生理生化特性初探[J].热带亚热带植物学报,2007,15(5):428–432.
[11]段继强,李建永,杜光辉,等.苎麻线粒体基因CoxⅡ和atpA与细胞质雄性不育相关性分析[J].中国农业科学,2009,42(2):434–445.
[12]邢虎成,揭雨成,周清明,等.苎麻雄性不育种质的鉴定分类研究[J].中国农学通报,2011,27(15):106–111.
[13]张建奎,董静,宗学凤,等.温光敏核雄性不育小麦雄性败育的细胞学观察[J].植物遗传资源学报,2009,10(3):411–418.
[14]伊六喜,张辉,贾霄云,等.雄性不育和可育亚麻的生殖期形态学与细胞学比较[J].内蒙古农业科技,2011(6):21–25.
[15]苏小俊,高军,袁希汉,等.萝卜雄性不育系的花器性状和形态学研究[J].江苏农业科学,2008(2):133–135.
[16]胡静静,赵静,沈向.黄金梨雄性不育的细胞学研究[J].中国农学通报,2010,26(2):185–188.
[17]莫宁捷,吕长平,成明亮,等.菏泽牡丹引种到长沙地区后的花芽分化[J].湖南农业大学学报:自然科学版,2008,34(2):193–195.
[18]吕长平,成明亮,莫宁捷.长沙地区芍药花芽形态分化研究[J].湖南农业大学学报:自然科学版,2009,35(2):142–144,154.
[19]熊少斌,杜威,龙波,等.苎麻雄性不育系与保持系花药发育的比较观察[J].云南大学学报:自然科学版,2011,33(6):735–739.
[20]聂明建,王国槐,朱卫平.甘蓝型油菜3种类型雄性不育系花药败育的细胞学研究[J].中国农业科学,2007,40(7):1543–1549.
[21]田英,李云翔,秦垦,等.宁夏枸杞雄性不育材料小孢子发生的细胞形态学观察[J].西北植物学报,2009,29(2):263–268.
[22]徐树华.水稻雄性不育系及其保持系颖花输导组织的比较观察[J].中国农业科学,1984,8(2):14–18.
