高温胁迫对广东万年青生理特性的影响
2014-06-08沈素贞聂东伶吴思政梁文斌柏文富
沈素贞,聂东伶,吴思政,梁文斌,柏文富
(1.中南林业科技大学生命科学院,湖南 长沙 410004;2.湖南省森林植物园,湖南 长沙 410116)
广东万年青(Aglaonema modestum Schott)又名亮丝草、斩蛇剑、冬不调草、铁扁担、九节连等,属天南星科广东万年青属多年生常绿直立草本植物,原产中国广东和菲律宾。广东万年青四季常绿,多生长于阴湿的环境中,最适生长温度为10~30 ℃,不宜在干燥和日光暴晒的环境中生长[1]。高温对其生长发育的影响较大。广东万年青是中国南方地区常见的园林观赏植物。该地区夏季高温天气(温度最高可达43 ℃)严重影响广东万年青的正常生长及其景观效应的发挥。国内外有关观赏植物耐高温特性的探讨[2–5]较多,对其研究主要集中在水培技术方面[6–7],而对广东万年青高温适应性的研究鲜有报道。笔者从植株形态及生理、生化等方面探寻广东万年青植株受高温胁迫伤害的机理和适应机制,旨在为其顺利渡过南方高温夏季和充分发挥景观效应提供参考。
1 材料与方法
1.1 材 料
试材为湖南省森林植物园苗圃长势基本一致、叶色浓绿、生长较旺的广东万年青盆栽苗,苗高约25 cm。
1.2 试验设计
2012年8月,将盆栽苗置于LRH–250A型智能人工气候箱中,25 ℃预处理3 d。预处理期间采用周期性光照(3 000 lx,13.5 h/d)条件培养,每天补水1次,使各处理土壤含水量基本保持在70%~75%。预处理结束后,将苗分放在5台人工气候箱中,设置5个处理温度:25 ℃(对照)、30 ℃(轻度高温胁迫,T1)、35 ℃(中度高温胁迫,T2)、40 ℃(重度高温胁迫,T3)、45 ℃(强高温胁迫,T4)。除温度外,其他条件与预处理保持一致。分别于各处理后2、4、6、8 d及常温(25 ℃)恢复3 d取样,并测定生理指标,观察其形态变化。每个处理3盆苗。重复3次。
1.3 测定指标与方法
在试验过程中,观察不同温度胁迫下的整体形态特征及叶片颜色变化、萎蔫状况;细胞质膜透性采用DDS–11A 型电导仪测定;丙二醛(MDA)含量采用硫代巴比妥酸(TBA)显色法测定;过氧化物酶(POD)活性采用愈创木酚显色法测定;可溶性糖含量采用蒽酮比色法测定。
1.4 数据统计与分析
分别采用Excel 2003和SPSS 19.0对数据进行统计分析。
2 结果与分析
2.1 高温胁迫对广东万年青叶片的伤害
观察发现,处理温度越高,叶片的形态变化越明显。对照组叶片浓绿;T1处理组叶片浅绿;T2处理组叶片稍有黄色;T3处理组叶片黄色明显,面积增大;T4处理组的叶片部分皱缩枯黄。温度越高,广东万年青叶片的伤害程度越大。25~30 ℃是其生长的适宜温度,超过40 ℃,叶片的外形会发生较大改变。
2.2 高温胁迫对广东万年青细胞膜透性的影响
如图1所示,在高温胁迫过程中,对照组的细胞质膜相对透性变化趋势平缓,其余各处理组均高于对照组。T1与T2处理组细胞质膜相对透性先略微下降后上升,常温恢复3 d后又下降,说明广东万年青可以在30 ℃和35 ℃正常生长;40 ℃和45 ℃高温下细胞质膜相对透性一直上升,其中T4处理组在胁迫第8天质膜透性剧烈上升,高达胁迫初期的3倍左右;常温恢复3 d后,质膜透性继续升高,说明植物细胞在此高温胁迫下受损严重且无法恢复,广东万年青无法在此高温下正常生长。

图1 各处理广东万年青细胞的质膜相对透性Fig.1 Relative permeability of plasma membrane of Aglaonema leaves under different temperatures
2.3 高温胁迫对广东万年青叶片丙二醛含量的影响
从图2可知,对照组丙二醛含量在整个处理过程变化不大,其余各高温处理组丙二醛的含量均高于对照组。T1和T2处理组丙二醛含量先升高后趋于平缓,T2比T1的增加幅度大,常温恢复3 d后,T1处理组有下降趋势,而T2处理组的下降趋势不明显。在40 ℃和45 ℃胁迫下,丙二醛含量一直呈增加趋势,且T4比T3增加的幅度大,直到高温解除恢复3 d后丙二醛含量也没有下降,说明过高的温度已经使植物体内的保护系统受到破坏,没有恢复的能力,丙二醛含量出现持续增加现象。综上分析可知,温度越高,广东万年青叶片丙二醛增加趋势越明显,超过40 ℃,广东万年青不能正常生长。
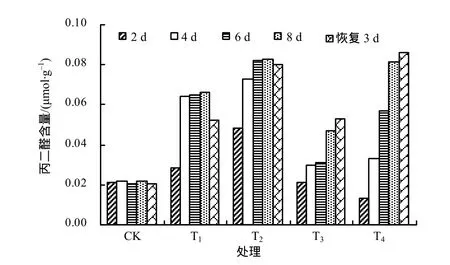
图2 各处理广东万年青叶片的丙二醛含量Fig.2 MDA content in Aglaonema leaves under different temperatures
2.4 高温胁迫对广东万年青过氧化物酶活性的影响
由图3可知,对照组的POD活性处于较高水平,且在整个处理过程中变化不大。T1处理的POD活性较对照组低,且随着胁迫时间的增加有下降趋势,恢复3 d后又上升,可见,30 ℃环境下广东万年青的生长没有受到影响。在35 ℃高温胁迫下,广东万年青POD活性在胁迫后4 d迅速上升,第6天急剧下降,高温胁迫解除后恢复3 d,活性有所恢复。T3和T4处理组POD活性的变化趋势与T2的类似,POD活性都呈现先迅速上升后急剧下降趋势,在这2个温度胁迫解除后恢复3 d,POD活性仍继续下降,说明广东万年青在此高温胁迫下已经受到严重伤害,无法恢复。
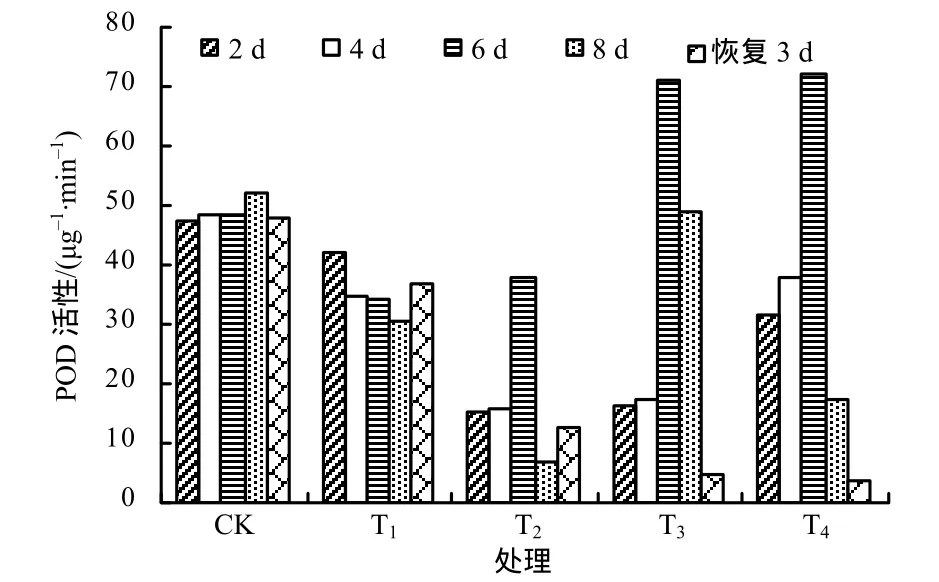
图3 各处理广东万年青叶片细胞的POD活性Fig.3 POD activities in Aglaonema leaves under different temperatures
2.5 高温胁迫对广东万年青可溶性糖含量的影响
由图4可知,对照组可溶性糖含量变化较平缓,其他各处理组在前4 d均增加,说明较高温度对可溶性糖的积累有促进作用。高温处理4~8 d,对照组和T1处理组的可溶性糖含量变化不大,说明广东万年青适宜在该处理温度范围生长;高于35 ℃的高温处理使可溶性糖含量迅速降低,说明此高温范围已经超过了植物体的承受能力。常温恢复3 d后,温度越高,处理的可溶性糖含量越低。可溶性糖含量与温度呈负相关。
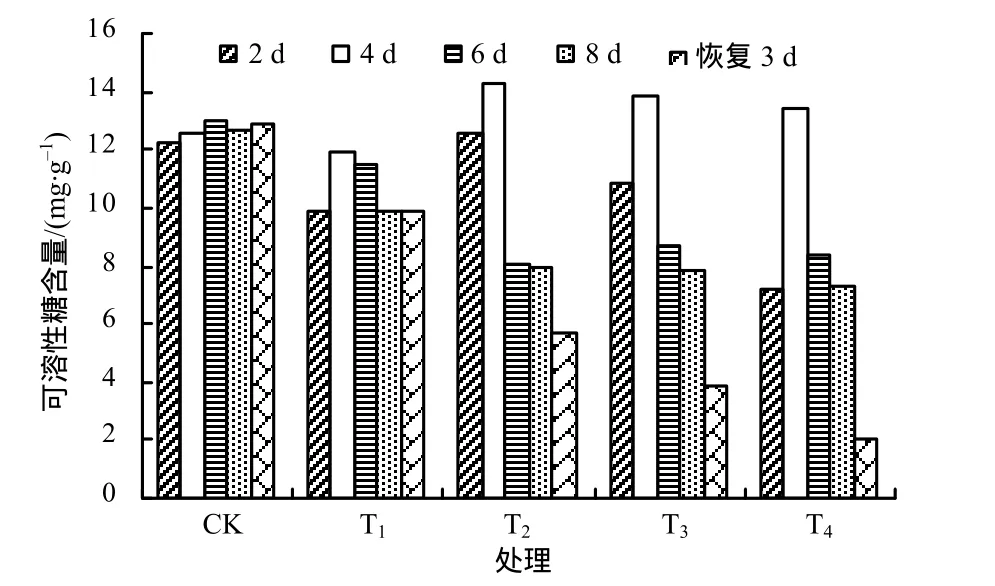
图4 各处理广东万年青叶片的可溶性糖含量Fig.4 Soluble sugar content in Aglaonema leaves under different temperatures
综上分析可知,高温在短期内对可溶性糖的积累起促进作用;随着高温的持续,可溶性糖含量降低,且温度越高,可溶性糖含量越低。
3 结论与讨论
植物受高温伤害的主要部位是细胞膜系统。大量研究表明,高温胁迫可导致植物叶片细胞膜受到破坏,质膜相对透性增高[8–9]。本研究中,广东万年青细胞质膜相对透性对不同高温胁迫的响应情况不同,在30 ℃与35 ℃高温胁迫下,细胞质膜相对透性随处理时间的增加呈先略微下降后平稳上升趋势。本研究结果与邓飞等[10]的研究结果一致。这可能是因为植物在受到高温刺激后,细胞质膜迅速启动保护应激,产生一些物质积累在细胞膜下,致使细胞质膜相对透性降低。随着胁迫时间的增加,细胞膜部分遭到破坏,电解质渗透率缓慢增加,细胞质膜相对透性增加。在40 ℃和45 ℃高温胁迫下,细胞质膜相对透性呈上升趋势,这可能是因为过高的温度超过了细胞的承受力,有些细胞无法抵御高温而遭到破坏,膜内电解质外渗,细胞膜透性大大提高。
关于丙二醛(MDA)含量与植物耐热性的关系,对不同植物的研究结果[12–13]不同。本研究中,温度的高低与广东万年青叶片丙二醛含量的增幅呈正相关,30 ℃和35 ℃胁迫4、6、8 d的丙二醛含量接近,40 ℃胁迫4、6 d的丙二醛含量接近,这与孙玉芳等[14]的研究结果一致。这可能是由植物可以承受一定时间范围的高温胁迫[14]所致。在高于35 ℃的高温环境中,广东万年青叶片内的丙二醛含量在常温恢复3 d后依然呈上升趋势,这可能是由植物在长时间高温环境中细胞膜系统遭到破坏而造成MDA含量持续增加所致。
POD可以清除体内的过氧化氢,防止过氧化氢和超氧离子相互作用,从而维持体内的活性氧代谢平衡[15]。广东万年青POD活性在30 ℃条件下较对照有所降低,恢复3 d后又上升,说明30 ℃抑制了POD的活性,但是没有破坏酶本身。在受到35、40、45 ℃高温胁迫时,POD活性随胁迫时间的增加呈先上升再下降的趋势。这可能是因为在胁迫初期,植物为了抵御胁迫伤害,通过叶片内活性氧激增诱导POD含量增加来清除超氧阴离子,从而保护植物免受伤害。随着胁迫时间的增长,植物受害程度增加,POD活性受到的影响开始减小。这与刘大林等[16]的研究结果基本一致。胁迫解除后,35 ℃胁迫处理的POD活性有恢复趋势,40 ℃和45 ℃胁迫处理的POD活性继续下降,说明广东万年青无法承受高于40 ℃的高温。
周中亮[17]认为,高温胁迫下,植物体内的可溶性糖作为能源物会不断消耗减少[17]。本研究中发现,随着胁迫时间的增加,广东万年青叶片可溶性糖含量大致出现先增加后降低的趋势,可能是因为在胁迫初期淀粉等大分子物质降解加快,可溶性糖含量上升,但胁迫一段时间后,大分子物质大量被降解,糖类不再积累,其含量趋于稳定甚至下降,胁迫后期可溶性糖含量的下降还可能是因为高温抑制了光合作用,减少了碳水化合物的生成,而高温加剧了呼吸作用和能量的消耗。广东万年青在35、40、45 ℃高温胁迫处理后常温恢复3 d,其叶片可溶性糖含量继续下降,说明这些高温胁迫已对广东万年青造成严重伤害。
植物的耐热性受多方面因素影响,其耐热机制也不相同,单凭个别指标对植物耐热性进行鉴定难免有失偏颇。本研究中将形态观察与生理指标相结合研究广东万年青的耐热性机理,初步认为广东万年青的最适生长温度为25~30 ℃。这与沈夏淦[1]的研究结果一致。综合分析后可得到如下研究结论:
1) 温度越高,对广东万年青叶片形态的伤害越大,25~35 ℃是其生长的适宜温度,超过40 ℃叶片的外形会受到严重破坏。
2) 叶片质膜相对透性经低于35 ℃胁迫处理后的变化不大,在40 ℃和45 ℃高温胁迫下上升,常温恢复3 d继续升高。
3) 丙二醛含量的增幅与温度成正相关,在高于35 ℃高温胁迫处理后常温恢复3 d,丙二醛含量未下降。
4) 低于30 ℃处理后过氧化物含量的变化不大;胁迫温度高于35 ℃后,温度越高,过氧化物酶活性的变化越大。
5) 高温在短期内对可溶性糖的积累起促进作用,但随着高温胁迫的持续,可溶性糖含量降低,且温度与可溶性糖含量呈负相关。
[1]沈夏淦.广东万年青[J].植物杂志,1998:23.
[2]王凯红,刘向平,张乐华,等.5种杜鹃幼苗对高温胁迫的生理生化响应及耐热性综合评价[J].植物资源与环境学报,2011,20(3):29–35.
[3]张施君,周厚高,钟云娟.高温胁迫对观赏百合苗期的生理影响研究[J].中国生态农业学报,2006,14(3):103–104.
[4]骆俊,韩金蓉,王艳,等.高温胁迫下牡丹的抗逆生理响应[J].长江大学学报,2011,8(2):223–226.
[5]姚士才,秦贺兰.5个小菊新品种耐热性综合评价与鉴定[J].中国农学通报,2006,22(12):217–219.
[6]黄小均,石大兴,王米力,等.水培广东万年青光合日变化研究[J].安徽农业科学,2005,33(5):799–800.
[7]孙廷,辛向阳,张建芳.广东万年青水培试验研究[J].安徽农业科学,2011,39(6):3220–3221.
[8]刘艳萍,姚莹莹,罗晓雅,等.高温胁迫对几种乔木树种生理生化特性的影响[J].河南农业科学,2011,40(11):126–128.
[9]吕长平,刘林艳.高温胁迫对芍药部分生理生化指标的影响[J].湖南农业大学学报:自然科学版,2008,34(6):664–667.
[10]任飞,邓少虹,卢春玲,等.蔷薇植物对高温胁迫的生理响应[J].南方农业学报,2012,43(11):1664–1667.
[11]康恩祥,陈年来,安翠香.低温弱光对西葫芦幼苗抗氧化酶活性和质膜透性的影响[J].中国蔬菜,2007(4):16–19.
[12]周斯建,义鸣放.高温胁迫下铁炮百合花幼苗形态及生理反应的初步研究[J].园艺学报,2005,32(1):145–147.
[13]程玲,邱永福,田志宏,等.不同温度对马蹄金生理生化特性的影响[J].四川草原,2004(10):23–27.
[14]孙玉芳,王三根,尹丽,等.高温胁迫对黄连生理特性的影响研究[J].中国农学通报,2006,22(4):236–238.
[15]杨东,罗群.温度胁迫对菊科杂草生理指标的影响及其适应[J].吉首大学学报,2006,27(4):75–79.
[16]刘大林,王秀萍,胡楷崎,等.高温胁迫对两种冷季型草坪草生理生化特性的影响[J].生态学杂志,2012,31(4):811-815.
[17]周中亮,包满珠,王文恩.高温胁迫对6个高羊茅株系生理指标的影响[J].草业科学,2011,28(7):1284–1290.
