不同产地木荷优树无性系生长和开花性状的分析
2014-05-11辛娜娜徐肇友王帮顺徐有明周志春
辛娜娜,张 蕊,徐肇友,王帮顺,徐有明,周志春
(1.华中农业大学园艺林学学院,湖北武汉430070;2.中国林业科学研究院亚热带林业研究所,浙江富阳311400; 3.浙江省龙泉市林业科学研究院,浙江龙泉323700)
不同产地木荷优树无性系生长和开花性状的分析
辛娜娜1,2,张 蕊2,徐肇友3,王帮顺3,徐有明1,①,周志春2
(1.华中农业大学园艺林学学院,湖北武汉430070;2.中国林业科学研究院亚热带林业研究所,浙江富阳311400; 3.浙江省龙泉市林业科学研究院,浙江龙泉323700)
为比较不同产地木荷(Schima superba Gardn.et Champ.)优树无性系的生长性状(包括树高、胸径、冠幅和一级侧枝数)和开花性状(包括开花物候期和花朵数量)的遗传差异,采用方差分析、相关性分析和聚类分析方法,对来源于福建建瓯和连城、浙江庆元及龙泉和遂昌、江西上犹的22个木荷优树无性系的生长和开花性状进行了综合分析。结果表明:供试木荷无性系的树高、胸径、冠幅和一级侧枝数的的变异系数为19%~32%,始花时段、盛花时段、末花时段和花朵数量的变异系数为37%~73%,表明不同无性系生长和开花性状的变异较大。除木荷无性系一级侧枝数产地间的变异未达到显著水平外,其他生长和开花性状产地间和产地内的变异均达到显著或极显著水平。各生长和开花性状的无性系重复力为0.41~0.94,显示遗传控制程度中等偏强。无性系的树高、胸径和冠幅与花期的相关性均不显著,但与花朵数量呈显著或极显著正相关;来自较低纬度的无性系始花期最早,来自较高纬度或海拔的无性系始花期最晚。聚类分析结果显示:22个木荷无性系可聚为4个类型,类型1和类型4分别包含5个和7个无性系,这2类无性系的树高、胸径、冠幅和花朵数量总体上高于群体均值,生长表现优良、花朵数量较多,且类型4包含的无性系产地纬度较低、开花日期较早;类型2包含8个无性系,生长表现一般、花朵数量较少;类型3包含2个无性系,生长表现优良、花朵数量低于群体均值。研究结果显示:依据产地的纬度和海拔可初步判定木荷无性系的花期,可为木荷杂交育种亲本的选配和种子园建园过程中无性系的配置提供科学依据和理论基础。
木荷;无性系;生长;开花;遗传变异;聚类分析
林木育种亲本的选择和杂交与优树的培育密切相关,而根据生长和开花性状可以筛选出优良的亲本和杂交后代,因此,开展育种亲本生长、开花及结实特性方面的观测与评价对育种群体评估、杂交亲本的科学选配以及种子园优选建园无性系选调等具有重要的理论和实践指导意义。国内外对树木花期开展了一系列的研究,通过对树木的生长性状、开花习性、花期同步性、遗传距离和开花物候等方面[1-6]的研究,了解并掌握产地经纬度、世代和生长性状表现对树木花期的影响以及遗传距离和开花物候对杂交亲本交配成功率的影响,旨在为树木的高世代育种提供科学的理论依据。
木荷(Schima superba Gardn.et Champ.)为山茶科(Theaceae)木荷属(Schima Reinw.)常绿大乔木,为中国亚热带常绿阔叶林的主要建群种,广泛分布于中国南方各省区[7],具有速生、丰产、材质优异和适应性强等特点,是中国南方重要的珍贵用材树种以及主栽的生物防火和生态防护树种。但有关木荷育种的研究起步较迟,涉及到育种亲本选择和配置及花期物候观察等方面的研究较少,而对木荷育种亲本的生长性状和开花性状的全面测评、记录和分析将直接影响其优良种子的产量和质量,也关系到木荷高世代育种的进展速度。
2008年至2013年,中国林业科学研究院亚热带林业研究所用材树种研究组从木荷优良种源区的天然林分中选择优树800余株,嫁接并保存木荷优树无性系600余株,以此建立了木荷的第1代育种群体。基于该育种群体,作者对来源于浙南、闽北、闽西和赣南等产地的部分木荷优树无性系的生长性状和开花习性进行观察和记录,研究各无性系生长性状与开花性状的遗传差异及其与产地纬度和海拔的关系,并通过性状表现对供试无性系进行聚类分析,以期为木荷杂交育种亲本的选配、种子园建园过程中亲本的配置及经营提供科学依据。
1 材料和方法
1.1 材料
供试木荷优树无性系为2008年以嫁接方法建立的木荷第1代育种群体,保存于浙江省龙泉市林业科学研究院上圩基地。地理坐标为东经119°05′、北纬28°02′,海拔30~50 m;属中亚热带季风气候,年均温18.3℃~11.5℃,年降水量1 400~2 275 mm,年日照时数1 712~1 825 h,无霜期180~280 d。
根据地势将育种群体划分为不同小区,其中1至4小区的无性系已进入开花结实期,分别为来自浙南、闽北、闽西和赣南的木荷优树,每个优树无性系嫁接保存6~10株,共计343株。按随机完全区组设计进行构建,株距2.0 m、行距2.5 m。
2013年5月至6月对木荷育种群体1至4小区中分别来自福建建瓯(FJJO)和连城(FJLC)、浙江庆元(ZJQY)及龙泉(ZJLQ)和遂昌(ZJSC)、江西上犹(JXSY)的22个开花的优树无性系进行生长性状和开花习性观测,各产地无性系数量分别为9、3、3、4、1和2个。各产地的自然概况见表1。
1.2 方法
每个无性系选择3株已进入花期且生长表现正常的样株进行树高、胸径、冠幅、一级侧枝数和花朵数量的调查及花期的观测。
生长性状的划定标准及测量方法如下:树高为顶梢到树干基部的垂直高度;冠幅为树冠东西向和南北向最大长度的平均值;胸径为距地面1.3 m处树干的直径,用电子游标卡尺(精度0.01 cm)测量;一级侧枝数为主干上着生的分枝数。在树冠中部东、南、西、北4个方向上各选择1支生长正常的一级侧枝,分别统计其上的花朵数并计算一级侧枝的平均花朵数量,一级侧枝数与平均花朵数量的乘积即为整个植株的花朵数量。
花期各时段的确定标准为:每株有15%花开放的日期为始花期,每株有45%花开放的日期为盛花期,每株有90%花开放的日期为末花期;始花期至盛花期间的持续天数为始花时段,盛花期至末花期间的持续天数为盛花时段,末花期至花全开日期间的持续天数为末花时段。
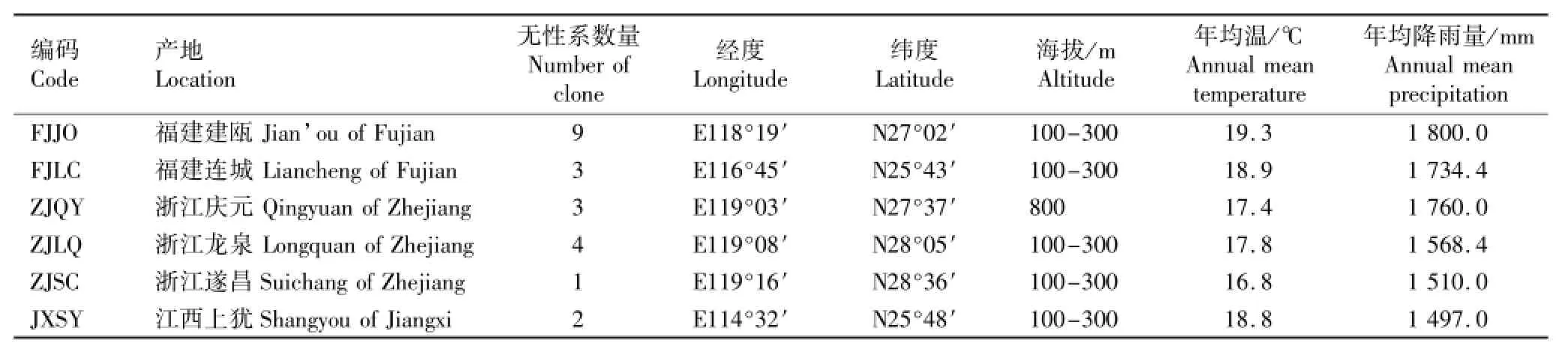
表1 供试木荷无性系数量及产地自然概况Table1 Clone number and location natural status of Schima superba Gardn.et Champ.tested
1.3 数据处理
参考Matziris[8]和梁机等[9]的方法,以各木荷优树无性系中最早记录的始花期5月15日为基点,观察并记录各植株花期与基点的间隔天数及花期不同时段的天数。树高、胸径和冠幅等指标的测定数据可直接用于统计分析,而一级侧枝数、花期及各花期的持续天数则采用平方根进行转换后用于统计分析。采用SAS 8.0软件中的GLM程序进行木荷优树无性系生长指标、花期和花朵数量等指标的方差分析。方差分析统计模型为Yijk=u+Pi+C(P)i(j)+eijk,式中,Yijk为第i个产地内第j个无性系第k个单株的观测值,u为群体平均值,Pi为第i个产地的效应,C(P)i(j)为第i个产地内第j个无性系的效应,eijk为机误。参照文献[10-12]的方法估算木荷生长性状和开花性状的无性系重复力及相关系数,并据此对各木荷优树无性系进行聚类分析[13]。
2 结果和分析
2.1 木荷优树无性系生长和开花性状的方差分析
木荷优树无性系生长和开花性状的方差分析结果见表2。
从表2可以看出:除一级侧枝数产地间的变异未达显著水平外,木荷优树无性系其他生长和开花性状在产地间和产地内的变异均达到显著或极显著水平。供试木荷优树无性系的树高、胸径、冠幅和一级侧枝数的平均值分别为2.5 m、2.70 cm、2.3 m和19.9,各生长性状的变异系数为19%~32%,表明不同优树无性系生长和分枝习性的遗传差异较大。由表2还可见:始花时段、盛花时段、末花时段和花朵数量的平均值分别为4.2 d、9.2 d、4.9 d和1 055.9,各开花性状的变异系数为37%~73%,说明供试木荷优树无性系的开花性状变异较大,因而,在进行木荷杂交亲本配置时应高度重视各无性系的花期差异,使无性系之间尽可能获得较多的交配机会。供试木荷优树无性系各生长和开花性状的无性系重复力为0.41~0.94,遗传控制程度中等偏强,因而,在杂交育种过程中需加强优树无性系生长性状的评价,并结合其子代测定优选育种亲本。
2.2 木荷优树无性系生长和开花性状的相关性分析
木荷优树无性系生长和开花性状的相关性分析结果见表3。
相关性分析结果显示:木荷优树无性系树高与胸径和冠幅之间呈极显著正相关;树高和胸径与一级侧枝数呈显著或极显著的正相关,说明速生无性系的分枝数较多,但一级侧枝数与冠幅的相关性不显著。供试木荷优树无性系的树高、胸径和冠幅与花期各指标的相关性均不显著,但与花朵数量呈显著或极显著正相关,这一现象为选择速生且花朵数量较多的无性系提供了证据。供试木荷优树无性系的始花期、始花时段、盛花期和盛花时段间呈极显著正相关,而始花时段与末花时段呈显著负相关,表明依据优树无性系的始花期可以判断其盛花期和末花时段。这一结果对了解木荷育种群体中优树无性系花期的持续时间及开展育种亲本的选配有一定的参考价值。
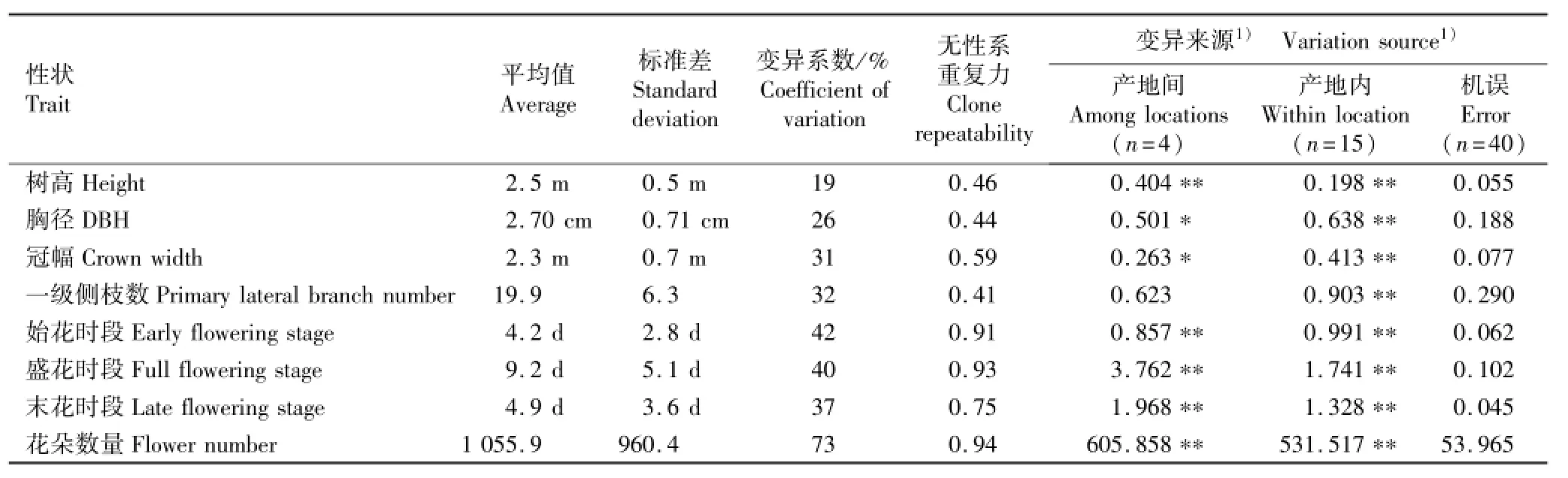
表2 木荷优树无性系生长和开花性状的方差分析结果Table2 Result of variance analysis on growth and flowering traits of Schima superba Gardn.et Champ.superior clones
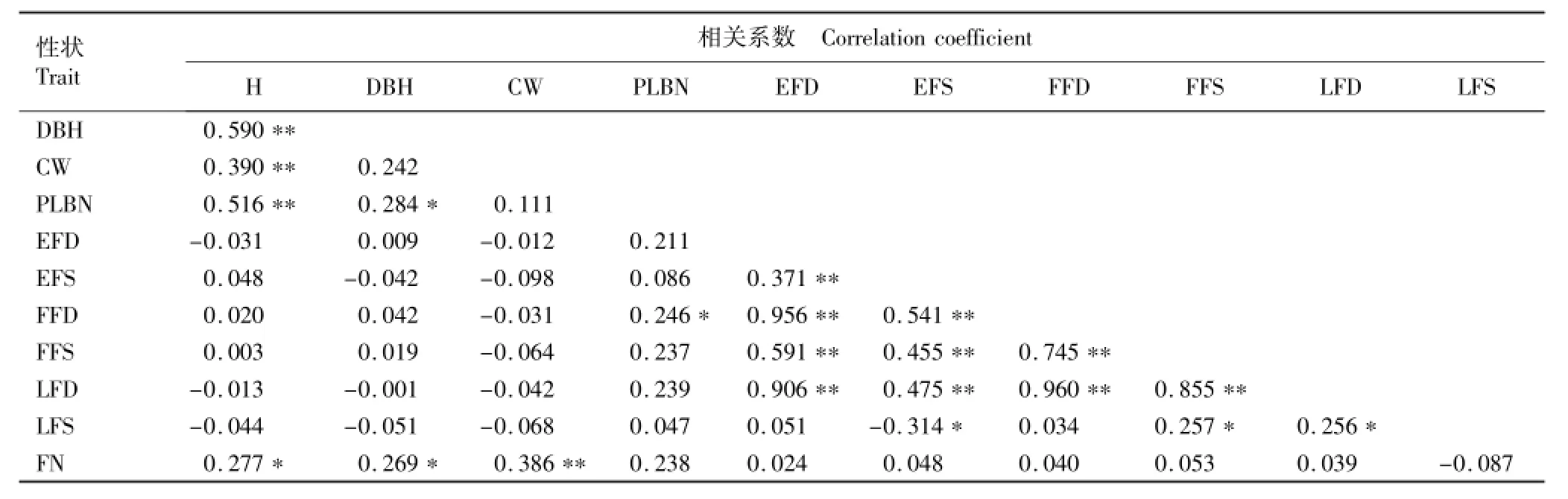
表3 木荷优树无性系生长和开花性状间的相关系数1)Table3 Correlation coefficient among growth and flowering traits of Schima superba Gardn.et Champ.superior clones1)
2.3 木荷优树无性系花期观测
供试木荷优树无性系花期及各花期持续时段的观测结果见表4。
由表4可以看出:最早进入始花期的优树无性系为产自福建连城的FJLC2,进入始花期、盛花期和末花期的日期分别为5月15日、5月19日和5月25日,分别持续4、6和2 d,共计12 d。最晚进入始花期的优树无性系是来自浙江庆元的ZJQY6、ZJQY9和ZJQY11以及来自浙江遂昌的ZJSC1,它们的始花期、盛花期和末花期完全一致,进入始花期、盛花期和末花期的日期分别为6月9日、6月15日和6月28日,分别持续6、13和3 d,共计22 d。进入始花期最早的无性系与进入始花期最晚的无性系间始花期相差25 d。结合产地的纬度和海拔综合分析可知,来自较低纬度产地的优树无性系(如产自福建连城的优树无性系FJLC2)进入始花期较早,而来自纬度相对较高产地的优树无性系(如产自浙江遂昌的优树无性系ZJSC1)进入始花期较迟;另外,来自纬度较低但海拔较高产地的优树无性系(如产自浙江庆元的优树无性系ZJQY6、ZJQY9和ZJQY11)的始花期也有明显的延迟现象。
由表4还可见:来自浙江龙泉的无性系ZJLQ7和来自江西上犹的无性系JXSY43的花期最短,整个花期仅持续8 d;来自福建建瓯的无性系FJJO9和FJJO45的花期最长,整个花期持续时间达32 d。从观测结果看,22个木荷优树无性系的始花期、盛花期和末花期的日期及各时段持续天数差别很大,例如,来自浙江龙泉的无性系ZJLQ7与ZJLQ10的花期完全不重叠。多数木荷优树无性系的花期会随着产地纬度和海拔的增高而推迟,也有个别优树无性系产地纬度较低但开花日期未提前(如产自福建建瓯的无性系FJJO9)。由此看出,木荷育种亲本原产地的纬度和海拔对其花期的影响较大,因而,在木荷杂交育种亲本选配和种子园建园材料选择时须充分考虑育种亲本原产地的纬度和海拔的差异。

表4 木荷优树无性系花期观测结果Table4 Observation result of flowering period of Schima superba Gardn.et Champ.superior clones
2.4 木荷优树无性系聚类分析和性状评价
根据22个木荷优树无性系的生长和开花性状进行Q型聚类分析,结果见图1。不同类型木荷优树无性系的生长和开花性状见表5。
由图1和表5可以看出:在欧氏距离0.178处, 22个木荷优树无性系被划分为4个类型。类型1包括无性系ZJQY9、ZJQY6、ZJQY11、ZJSC1和FJJO9,其中大多数无性系产自浙江;该类型无性系的树高、胸径和冠幅等生长性状表现优良,花朵数量较多。类型2包括无性系ZJLQ30、JXSY43、ZJLQ10、ZJLQ8、 FJJO4、FJJO2、JXSY42和FJLC39,其中包括的无性系除来自浙江和福建两省外,产自江西的2个无性系也划分在此类型中;该类型无性系的树高、胸径和冠幅均小于群体均值,但与群体均值的差异均较小,生长性状表现一般,花朵数量相对偏少,仅为群体均值的62.5%。类型3包括无性系FJJO8和FJJO45,均产自福建建瓯;该类型无性系的树高、胸径和冠幅等生长性状表现较为优良,花朵数量略低于群体均值。类型4包含无性系ZJLQ7、FJJO31、FJLC31、FJLC2、FJJO32、FJJO21和FJJO34,其中大多数无性系产自福建;该类型无性系的树高、胸径、冠幅和花朵数量都略高于群体均值,生长性状表现优良,花朵数量也较多,且产地纬度较低、开花日期较早。
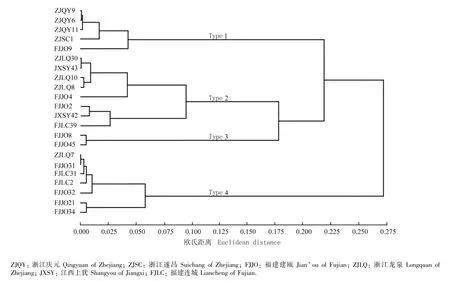
图1 基于生长和开花性状分析的22个木荷优树无性系的聚类图Fig.1 Cluster dendrogram of 22 superior clones of Schima superba Gardn.et Champ.based on analysis on growth and flowering traits

表5 不同类型木荷优树无性系的生长和开花性状比较Table5 Comparison on growth and flowering traits of different types of Schima superba Gardn.et Champ.superior clones
3 讨论和结论
方差分析结果显示:供试22个木荷优树无性系的树高、胸径和冠幅等生长性状及开花性状在产地间和产地内的变异总体上达到显著或极显著水平。育种群体由来源于不同产地的优树组成,无性系间的生长性状和开花习性差异较大,意味着除需加强优树无性系当代相关性状的观察外,同时还必须开展子代相应性状的观察,才能对育种亲本进行全面综合的评价。由于本研究涉及的育种群体中木荷亲本无性系的嫁接时间较短,因此无法对其材积等性状进行测评,但它们的树高和胸径的无性系重复力均较高,因而,可以将不同类型木荷无性系的生长表现作为育种亲本选配的依据。相关性分析结果显示:供试木荷优树无性系的树高和胸径与花期的相关性均不显著,这一研究结果与张一等[14]对马尾松(Pinus massoniana Lamb.)生长性状与开花性状相关性的研究结果相似。
对木荷优树无性系花期的观察结果表明:22个优树无性系的始花期、盛花期和末花期的日期及各时段天数存在很大差异。多数无性系的始花期随着产地纬度的升高而推迟,这与梁机等[9]获得的“1.5代高纬度杉木〔Cunninghamia lanceolata(Lamb.)Hook.〕无性系花期会推迟”的结果类似;无性系的产地海拔越高,开花日期越晚,因此,依据产地的纬度和海拔可初步判定木荷无性系的花期。此外,花期的长短还与年份、日照时数、温度和降雨量等因子有关,但由于作者未对物候因子进行详细记录,研究结果稍显不足。今后,应开展对木荷无性系开花习性和物候因子的连年观测与分析,以期揭示整个木荷育种群体中无性系的开花习性与不同年份物候因子间关系的规律,为杂交育种亲本选配和种子产量预测提供科学的数据。
在生长和开花性状比较分析的基础上通过聚类分析可将供试的22个木荷优树无性系分为4个类型。在各类型内无性系的树高、胸径及花期等性状相近,而在不同类型间无性系的生长性状和开花性状却存在明显差异。类型1包括的无性系产地纬度和海拔均较高(如浙江庆元海拔达800 m),推测该类型无性系的抗寒能力可能强于其他类型的无性系[15];但由于产地的纬度和海拔相对较高,类型1中各无性系的开花日期均相对较迟。类型4包括的无性系产地纬度均较低,如福建连城和建瓯,这两个产地均为木荷的中心产区,水热资源优越,木荷的快速生长能力相对较强。类型2和类型3包括的无性系产地纬度在4个类型中居中,但各无性系的生长状况则并非介于类型1和类型4之间,这一研究结果与张一等[14]对马尾松的相关研究结果不同;而这些无性系的开花日期却随着纬度增高而推迟,说明产地纬度对木荷无性系开花日期的影响强于对其生长性状的影响。木荷优树无性系的生长状况和开花习性均为与原产地环境相适应的结果,其中,开花习性除受产地气候因素的影响外,还受到产地纬度和海拔等地理因素的影响。依据这一规律,将不同产地的木荷育种亲本无性系进行杂交,有可能筛选出具有亲本优良特性的后代。如来源于海拔相对较高的浙江庆元的木荷无性系具有抗寒优势,而来源于低纬度的福建连城和建瓯的木荷无性系则具有速生优势,它们的杂交后代则可能具有速生和抗寒等优良特性。
[1] WEBBER J E,YEH F C H.Test of the first-in,first-in pollination hypothesis in coastal Douglas-fir[J].Canadian Journal of Forest Research,1987,17:63-68.
[2] ERICKSON V J,ADAMS W T.Mating success in a coastal Douglasfir seed orchard as affected by distance and floral phenology[J]. Canadian Journal of Forest Research,1989,19:1248-1255.
[3] 王润辉,胡德活,郑会全,等.杉木2.5代种子园开花物候遗传变异分析[J].西南林业大学学报,2013,33(4):25-29.
[4] 段爱国,黄健韬,章允清,等.闽北山地不同世代杉木种子园球花产量调查[J].福建林学院学报,2010,30(1):39-44.
[5] 谭小梅.马尾松二代育种亲本选择及种子园交配系统研究[D].富阳:中国林业科学研究院亚热带林业研究所,2011.
[6] 谭小梅,金国庆,张 一,等.截干矮化马尾松二代无性系种子园开花结实的遗传变异[J].东北林业大学学报,2011,39(4): 39-42.
[7] 倪 健.中国木荷及木荷林的地理分布与气候的关系[J].植物资源与环境,1996,5(3):28-34.
[8] MATZIRIS D I.Genetic variation in the phenology of flowering in black pine[J].Silvae Genetica,1994,43:321-328.
[9] 梁 机,周传明.1.5代杉木种子园开花规律的观察分析[J].广西农业大学学报,1998,17(3):285-292.
[10] 孔繁玲.植物数量遗传学[M].北京:中国农业大学出版社, 2006:224-249.
[11] 续九如.林木数量遗传学[M].北京:高等教育出版社,2006: 26-33.
[12] ISIK F,GOLDFARB B,LeBUDE A,et al.Predicted genetic gains and testing efficiency from two loblolly pine clonal trials[J]. Canadian Journal of Forest Research,2005,35:1754-1766.
[13] 黄少伟,谢维辉.实用SAS编程与林业试验数据分析[M].广州:华南理工大学出版社,2001:182-196.
[14] 张 一,谭小梅,刘伟宏,等.马尾松二代育种群体生长和开花结实性状[J].南京林业大学学报:自然科学版,2011,35 (2):1-7.
[15] 沈 漫,王明庥,黄敏仁.植物抗寒机理研究进展[J].植物学通报,1997,14(2):1-8.
(责任编辑:张明霞)
Analysis on growth and flowering traits of Schima superba superior clones from different locations
XIN Na’na1,2,ZHANG Rui2,XU Zhaoyou3,WANG Bangshun3,XU Youming1,①,ZHOU Zhichun2(1.College of Horticulture and Forestry,Huazhong Agricultural University,Wuhan 430070,China; 2.Research Institute of Subtropical Forestry,Chinese Academy of Forestry,Fuyang 311400,China; 3.Forestry Academy in Longquan City of Zhejiang Province,Longquan 323700,China),J.Plant Resour.&Environ.2014,23(4):33-39
In order to compare genetic difference of growth and flowering traits of Schima superba Gardn. et Champ.superior clones from different locations,taking methods of variance analysis,correlation analysis and cluster analysis,growth trait(including height,DBH,crown width and primary lateral branch number)and flowering trait(including flowering phenophase and flower number)of 22 superior clones of S.superba from Jian’ou and Liancheng of Fujian,Qingyuan,Longquan and Suichang of Zhejiang,Shangyou of Jiangxi were comprehensively analyzed.The results show that coefficient of variation of height,DBH,crown width and primary lateral branch number of S.superba clones tested are 19%-32%,that of early flowering stage,full flowering stage,late flowering stage and flower number are 37%-73%,which indicates that variations of growth and flowering traits of different clones are great. Except variation of primary lateral branch number of S.superba clones among different locations is notsignificant,variations of other growth and flowering traits among different locations and within location reach significant or extremely significant levels.Clone repeatability of all growth and flowering traits is 0.41-0.94,which shows medium to strong genetic control degree.Height,DBH and crown width of clones have no significant correlation with flowering period,but they have significantly or extremely significantly positive correlations with flower number.Early flowering date of clones from locations with lower latitude is the earliest,and that of clones from locations with higher latitude or altitude is the latest. The cluster analysis result shows that 22 clones of S.superba can be classified into 4 types.In which, type 1 and type 4 include 5 and 7 clones,respectively,and height,DBH,crown width and flower number of clones in the two types are totally higher than their average value of all clones,and clones of the two types grow well and have more flowers,and clones in type 4 are from lower latitude location and their flowering date is earlier;type 2 includes 8 clones,grows normal and has less flowers;type 3 includes 2 clones,grows well and its flower number is lower than average value of all clones.It is suggested that according to latitude and altitude of location,flowering period of S.superba clones can be determined preliminarily,and scientific and theoretical bases on cross breeding parental selection and clone configuration of seed garden construction of S.superba can be provided.
Schima superba Gardn.et Champ.;clone;growth;flowering;genetic variation;cluster analysis
S718.42;S792.99
A
1674-7895(2014)04-0033-07
10.3969/j.issn.1674-7895.2014.04.05
2014-03-05
国家“十二五”科技支撑计划项目(2012BAD01B04);国家林业公益性行业科研专项重大项目(201104001);浙江省农业新品种选育重大科技专项(2012C12908-6);福建省林木种苗科技攻关四期项目
辛娜娜(1987—),女,安徽阜阳人,硕士研究生,主要从事林木遗传育种方面的研究。
①通信作者E-mail:xuyouming@mail.hzau.edu.cn
