沙培条件下磷、 钾、 钙亏缺对枳(Poncirus trifoliata)幼苗根系形态及营养吸收的影响
2014-04-08夏仁学杨环宇张德健
曹 秀, 夏仁学*, 杨环宇, 张德健, 赵 禹
(1 华中农业大学教育部园艺植物生物学重点实验室, 武汉 430070; 2 四川省资阳市雁江区农业局, 四川资阳 641399)
根系是树体生长发育的基础,对于多年生木本果树的生长发育尤为重要[1]。近年来,随着研究方法的改进和现代根系定量分析技术的发展,人们对根系在土壤中的空间分布已进行了大量研究[2]。特别是20世纪90年代以后,人们试图应用拓扑学理论将三维根构型分解成二维根构型, 进而通过测定有关的拓扑学参数对根构型给予定量描述[3-4]。二维根构型参数测量方法中的根系图像分析系统因具备不受根形状等因素的限制、 测量速度快、 人为因素干扰少、 精度高等优点而成为植物根系构型定量研究的有力工具[5]。目前,多数研究果树根系的学者主要从植物营养、 根系生态、 根际微生物等角度探讨其与土壤环境的相互关系[6,7],但是由于果树根系庞大、 分布错综复杂,对根系构型和空间分布等的研究相对较少。一般来说,根系的生长状况除受遗传特性的影响之外,土壤环境尤其是土壤养分状况、 水分和温度等的影响也是不可忽视的因素。果树根系虽然按照遗传特性发生, 但其生长习性受环境影响较大[1],其中土壤养分的有效性高低和分布状况对根系的形态构型有一定的调节控制作用。例如磷对主根的生长和侧根形成与生长有较大影响[8, 9],钾[10, 11]、 铁[12]、 硼[13]等养分对植物根系构型也有显著的影响。
柑橘为多年生常绿木本植物,多以枳(Poncirustrifoliata)为砧木。基于之前作者等研究发现的磷、 钾、 钙等矿质养分对枳根系的生长发育影响较大[14],故本研究在沙培条件下,对枳实生幼苗分别进行缺磷、 钾、 钙处理,利用计算机扫描及图像分析软件观测其根系总长、 总体积和总表面积等二维形态参数,分析不同粗度根系的生长发育特征,并用原子吸收法测定不同处理植株内的养分浓度,以探讨3种营养元素亏缺对枳根系形态的影响,以及根系形态变化对植株养分吸收的影响及其与植株生长间的关系,以期为深入了解柑橘砧木枳对不良土壤环境的适应性及提高其对土壤养分的利用率提供理论依据。
1 材料与方法
1.1 试验材料与设计

试验为单因素设计,共设4个处理,每处理3次重复。即以全素营养液(1/2Hoagland营养液)为对照,缺素处理分别为缺P、 K和Ca(表1)。处理后8个月采样,每重复选取长势较一致的9株植株进行相关指标的测定。

表1 试验处理及其配方
1.2 取样和测定方法
1.2.1 生长指标测定 每重复取样6株,用常规方法测定株高、 茎粗、 主根长度、 主根直径、 地上部和根系鲜重等生长指标。然后所有材料被封装后置于65℃烘箱中干燥至衡重(约24 h),称量其干样质量。
1.2.2 根系形态指标测定 每重复另取3株生长较一致的植株测量根系形态指标。采样后剪取植株根系用蒸馏水清洗干净后置于爱普生数码扫描仪(Expression 10000XL 1.0, Epson Inc. Japan)中扫描,并用WinRhizo Pro (S) v. 2004b软件(Regent Instruments Inc., Canada)分析获取根系形态相关指标根长、 平均根直径、 根表面积和根粗度分级等数据。
1.2.3 植株营养元素浓度测定 将1.2.1中干样研磨成粉末状,过100目筛(孔径0.15 mm),每个样品称取0.3 g于500℃马弗炉(Muffle)中干灰化6 h,用0.1 mol/L HCl溶解。用原子吸收光谱仪(SPECTR AA220)测定样品K、 Ca、 Mg、 Fe、 Cu、 Zn的含量。
1.3 数据处理
用Excel软件对数据进行初步整理和统计分析后,用SAS统计软件的ANOVA程序进行单因素方差分析(one-way ANOVA),并对不同处理间差异进行LSD法多重比较,显著性水平设定为α=0.05。
2 结果与分析
2.1 不同缺素处理枳实生苗生长发育状况
枳实生苗生长状况统计结果(表2)表明,在沙培条件下,缺磷显著降低了地上部和根系的生长量,对地上部的抑制作用强于根部,其中株高、 地上部干重和鲜重、 茎粗均显著低于其他各处理,分别比对照降低了49.7%、 60.2%、 68.0%和22.2%; 根部鲜重降低了37.1%,但主根直径显著增加,比对照增加了13.0%,且显著高于其他处理。缺钾处理对地上部生长影响较小,除地上部鲜重低于对照外,株高、 茎粗等与对照差异不显著; 但其对根系生长影响较大,与对照相比,缺钾处理植株根系干重、 鲜重以及主根直径均显著降低。缺钙使枳实生苗株高、 地上部鲜重和主根直径显著低于对照。
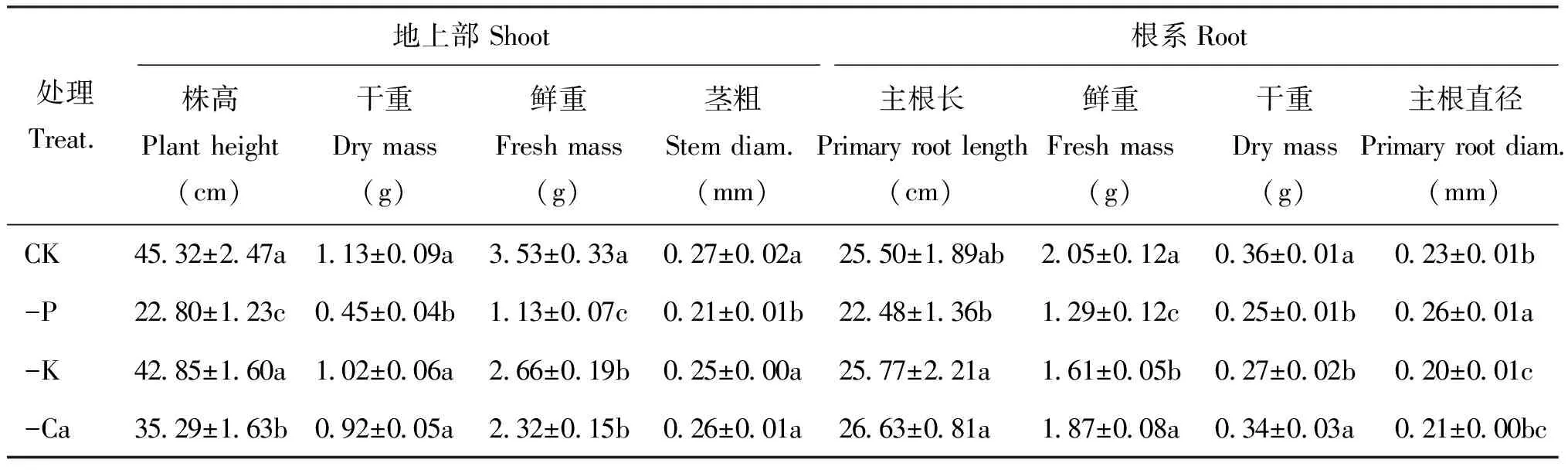
表2 缺素处理对枳实生幼苗地上部和根系生长的影响
2.2 不同缺素处理对枳幼苗根系形态的影响
不同缺素处理对枳实生幼苗根系的总长、 总表面积和总体积等形态结构的影响不尽相同(图1)。从图1中可以看到,缺磷、 钾以及钙处理, 根系总长、 总表面积和总体积均显著减少, 其中缺磷处理减少幅度最大,与对照相比分别减少了41.7%、 29.1%和 16.8%; 但缺磷处理却使根系平均直径增加了16.6%,显著大于其他处理,而缺钾和缺钙处理植株根系平均直径与对照无显著差异。
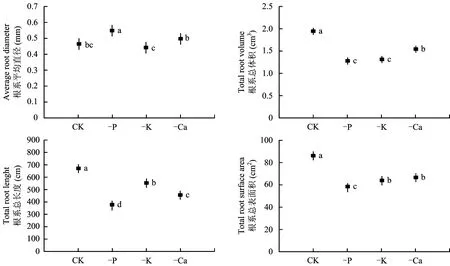
图1 缺素处理对枳幼苗根系总长度、 总表面积和总体积及根系平均直径的影响Fig.1 Effect of nutrients deficiency on the average root diameter, total root volume, length and surface area of Poncirus trifoliata seedlings
沙培条件下,缺磷、 钾、 钙均不同程度地抑制了枳实生幼苗根系的生长,使根系生物量显著低于对照。其中缺磷除对主根直径有增粗的作用外,对侧根的发生和伸长也有一定的促进作用,这可能是因为缺磷处理使根系的横向扩展速率超过纵向伸长速率所致; 缺钾对主根长度无显著影响,但其主根直径显著小于对照,且侧根数量和根系分枝显著减少; 而缺钙处理仅使根系分枝减少(图2)。
2.3 缺素处理对枳实生幼苗不同直径根系生长的影响
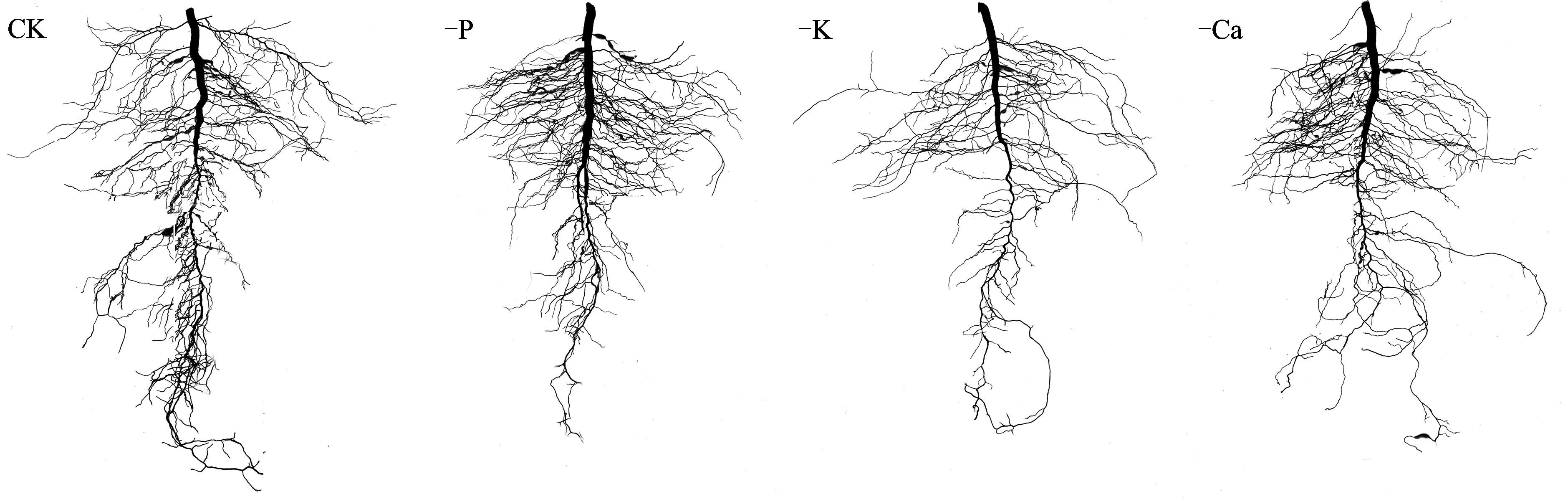
图2 不同缺素处理下枳实生幼苗的根系形态Fig.2 The root morphology of Poncirus trifoliata seedlings grown in different nutrients deficiency media
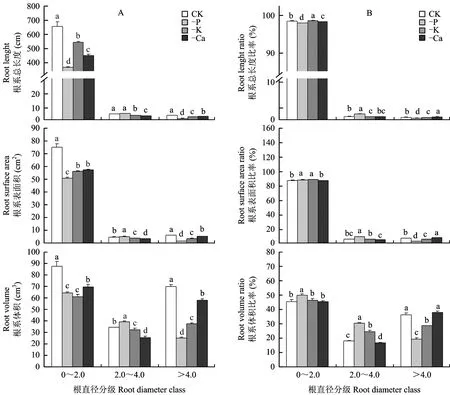
图3 缺素处理枳幼苗不同直径根系的根长、 根表面积、 根体积(A)及其所占总量比例(B)Fig.3 The volume, length, surface area (A) and the ratios (B) of root fractions of Poncirus trifoliata seedlings under different nutrient difficiency
2.4 不同缺素处理对枳幼苗根系和地上部营养元素含量的影响
从表3可以看出,与对照相比,缺磷条件下枳实生苗根系和地上部Ca含量显著升高,根系Cu浓度升高而Fe浓度显著降低,地上部Zn浓度显著升高而K浓度显著降低; 缺钾时植株根系和地上部Ca、 Cu和Zn含量显著高于对照而K和Fe含量显著降低,根系Mg显著高于对照; 缺钙处理的枳实生苗根系和地上部Fe和Zn浓度显著增加,根系Cu含量显著降低,地上部K含量显著增加而Ca含量显著降低。
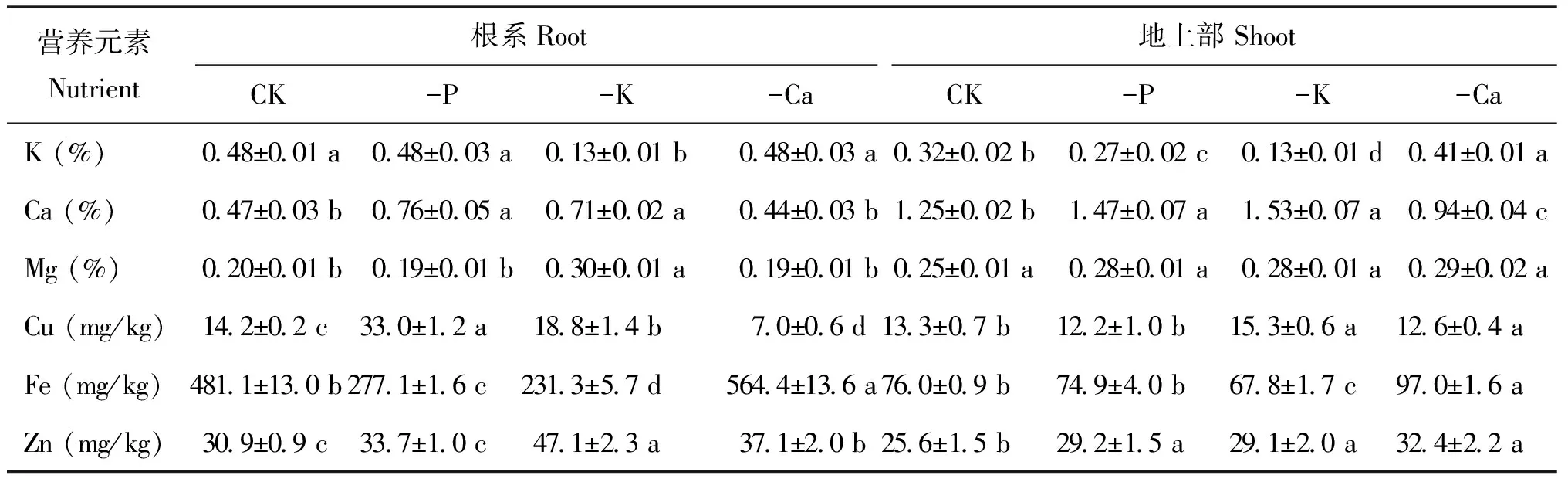
表3 不同缺素条件下枳实生幼苗根系和地上部营养元素的浓度
3 讨论
根系总体积由总表面积和平均直径共同决定,而根系总长和总表面积与其吸收能力关系密切[15]。磷、 钾和钙是保持植物正常生长发育所必需的营养元素,在根区环境缺素时,根系生长发育必然受到抑制。缺磷、 钾、 钙处理后,枳实生幼苗的根表面积、 根体积等根系参数与对照相比均显著降低,根系构型发生了不同程度的变化。从3种缺素处理的枳幼苗根系整体形态来看,其变化与枳植株整体生长有较显著的相关性。3种缺素处理的枳幼苗根系生长受抑,其植株总生长量也显著低于对照。缺素处理植株根长、 根系表面积和体积显著降低,导致有效吸收面积和吸收范围缩小,根系吸收能力下降,而对照苗根系相对发达,在基质中分布较均匀,加强了对养分和水分的吸收。缺磷、 缺钾和缺钙枳实生苗表现出缺素的特征,但三者对根系形态和生长状况的影响不尽相同。缺磷处理对枳幼苗地上部生长抑制作用较根系强,缺钾处理与之相反,对整个根系的生长发育均有显著的抑制作用,缺钙对植株整体生长的抑制作用较缺磷和缺钾小。
根系形态可影响枳幼苗根系吸收养分的能力,从而影响植株的生长。不同直径根系的吸收能力存在差别, 一般认为细根的吸收能力强于粗根[16],缺磷处理不仅抑制了枳幼苗根系生长,且使细根总量和比例降低,导致根系吸收能力降低进而抑制了植株的生长。结果表明,缺磷处理使枳实生苗地上部锌浓度增高,这可能是由于缺磷导致地上部生物量的绝对下降,相对增加了Zn的浓度。植株地上部锌浓度增高能促进生长素的合成,生长素的极性运输使根部生长素浓度显著增加,从而促使地上部碳水化合物调运至根部[17],而且生长素增加能促发根尖细胞周期蛋白基因的表达,驱动侧根发生和伸长[17],笔者之前的研究也发现缺磷和生长素处理能促进枳侧根发生和根毛发育[14,18],这可能是缺P胁迫对枳地上部生长的抑制作用强于根系的原因之一。与缺磷相似的是缺钙时植株地上部锌浓度增高,这有利于光合产物向根部调运,同时地上部和根系铁浓度也显著增加,这保证了植株光合作用的进行,因此铁和锌浓度的增加可能是导致缺钙(与缺磷和钾相比)对枳生长影响较小的原因。
钾虽然不参与细胞组成,但能够调节与光合、 代谢作用等相关的多种酶的活性且在细胞渗透调节中有重要功能。缺钾处理的枳幼苗K浓度显著降低,钾以离子形式存在于植物体内,具有高度的移动性[19]。已有研究表明,K影响根系生长的作用机制与 N、 P 等不同, K浓度的改变能引起整个根系的系统响应[20], 这可能与其高度移动性有关。此外,低钾胁迫对枳幼苗养分浓度有较大的影响(根系中所测的6种元素均与对照差异显著),其中根系和地上部Fe浓度均显著降低。Fe、 K直接参与植物光合作用、 调节光合酶活性且与光合产物的运输密切相关, Fe浓度降低使地上部分光合受阻[21],从而可能显著降低了植株叶片光合产物合成和转运能力,导致根系生长受抑[22-23]。因此,缺钾处理可能通过影响植株K、 Fe等的含量,调节光合产物的合成和运输以及一些相关酶的活性,改变枳幼苗根系形态进而影响整个植株的生长。
参考文献:
[1] 束怀瑞, 罗新书, 吴光林. 果树栽培生理学[M].北京: 中国农业出版社, 1993.
Shu H R, Luo X S, Wu G L. Pomology cultivation physiology[M]. Beijing: China Agriculture Press, 1993.
[2] 马献发, 宋凤斌, 张继舟. 根系对土壤环境胁迫响应的研究进展[J].中国农学通报, 2011, 27(5): 44-48.
Ma X F, Song F B, Zhang J Z. Advances of research of roots responses to environmental stress on soil[J]. Chinese Agricultural Science Bulletin, 2011, 27(5): 44-48.
[3] Armengaud P, Zambaux K, Hills Aetal. EZ-Rhizo: integrated software for the fast and accurate measurement of root system architecture[J]. The Plant Journal, 2009, 57(5): 945-956.
[4] Bot J L, Serra V, Fabre Jetal. DART: a software to analyse root system architecture and development from captured images[J]. Plant and Soil, 2010, 326 (1-2): 261-273.
[5] 梁泉, 廖红, 严小龙. 植物根构型的定量分析[J].植物学通报, 2007, 24(6): 695-702.
Liang Q, Liao H, Yan X L. Quantitative analysis of plant root architecture[J]. Chinese Bulletin of Botany, 2007, 24(6): 695-702.
[6] Kroon H D, Hendriks M, Ruijven J Vetal. Root responses to nutrients and soil biota: drivers of species coexistence and ecosystem productivity[J]. Journal of Ecology, 2012, 100(1): 6-15.
[7] Remans T, Thijs S, Truyens Setal.Understanding the development of roots exposed to contaminants and the potential of plant-associated bacteria for optimization of growth[J]. Annals of Botany, 2012,110(2): 239-252.
[8] Péret B, Clément M,Nussaume Letal. Root developmental adaptation to phosphate starvation: better safe than sorry[J]. Trends in Plant Science, 2011, 16(8): 442-450.
[9] Sato A, Miura K. Root architecture remodeling induced by phosphate starvation[J]. Plant Signaling & Behavior, 2011, 6(8): 1122-1126.
[10] 张志勇, 王清连, 李召虎, 等.缺钾对棉花幼苗根系生长的影响及其生理机制[J].作物学报, 2009, 35(4): 718-723.
纯文学处境尴尬,意味着纯文学一枝独秀的时代早已终结,文学领域出现多声部合唱。譬如陕西省合阳县路井镇农民侯永禄仅有初中文化程度,但他坚持写了六十年日记,总计200余万字,真实记录了自己和村民的生存经历,记录了抗日战争到改革开放等不同时代背景下的“三农”问题。经过编辑处理,20余万字的《农民日记》2006年由中国青年出版社出版后,数次再版。
Zhang Z Y, Wang Q L, Li Z Hetal. Effect of potassium deficiency on root growth of cotton (GossypiumhirsutumL.) seedlings and its physiological mechanisms involved[J]. Acta Agronomica Sinica, 2009, 35(4): 718-723.
[11] 潘艳花, 马忠明, 吕晓东, 等.不同供钾水平对西瓜幼苗生长和根系形态的影响[J].中国生态农业学报, 2012, 20(5): 536-541.
Pan Y H, Ma Z M, Lv X Detal. Effects of different potassium nutrition on growth and root morphological traits of watermelon seedling[J]. Chinese Journal of Eco-Agriculture, 2012, 20(5): 536-541.
[12] Schmidt W, Tittel J, Schikora A. Role of hormones in the induction of iron deficiency responses inArabidopsisroots[J]. Plant Physiology, 2000, 122(4): 1109-1118.
[13] Mei L, Sheng O, Peng S-Aetal. Growth, root morphology and boron uptake by citrus rootstock seedlings differing in boron-deficiency responses[J]. Scientia Horticulturae, 2011, 129(3): 426-432.
[14] 曹秀, 夏仁学, 张德建, 等.水培条件下营养元素对枳幼苗根毛发育及根生长的影响[J].应用生态学报, 2013, 24(6): 1525-1530.
Cao X, Xia R X, Zhang D Jetal. Effects of nutrients on the seedlings root hair development and root growth ofPoncirustrifoliataunder hydroponics condition[J]. Chinese Journal of Applied Ecology, 2013, 24(6): 1525-1530.
[15] 林雅茹, 唐宏亮, 申建波. 野生大豆根系形态对局部磷供应的响应及其对磷吸收的贡献[J]. 植物营养与肥料学报, 2013, 19(1): 158-165.
[16] Sullivan W M, Jiang Z, Hull R J. Root morphology and its relationship with nitrate uptake inKentuckybluegrass[J]. Crop Science, 2000, 40(3): 765-772.
[17] 孙海国, 张福锁.小麦根系生长对缺磷胁迫的反应[J].植物学报, 2000, 42(9): 913-919.
Sun H G, Zhang F S. Grouth response of wheat roots to phosphrus deficiency[J]. Acta Botanica Sinica., 2000, 42(9): 913-919.
[18] 张德建, 夏仁学, 曹秀, 等.生长调节剂和基质对枳根毛发育的影响[J].应用生态学报, 2011, 22(6): 1437-1442.
Zhang D J, Xia R X, Cao Xetal. Effects of growth regulators and media on root-hair development ofPoncirustrifoliata[J]. Chinese Journal of Applied Ecology, 2011, 22(6): 1437-1442.
[19] 李俊林, 高南, 刘春生, 等.水稻钾离子通道基因OsKAT1.1的克隆,表达载体的构建及其电生理功能[J]. 基因组学与应用生物学, 2011, 30(4): 346-351.
Li J L, Gao N, Liu C Setal. Cloning, constraction of expression vectors and electrophysiological fonction of potassium channel geneOsKAT1.1 in rice[J]. Genomics and Applied Biology, 2011, 30(4): 346-351.
[20] 陈波浪, 盛建东, 蒋平安, 等.钾营养对水培棉花生长发育的影响[J].中国农学通报, 2008, 24(11): 267-271.
Chen B L, Sheng J D, Jiang P Aetal. Effect of potassium nutrition on growth of cotton in liquid culture[J]. Chinese Agricultural Science Bulletin, 2008, 24(11): 267-271.
[21] 刘延吉, 田晓艳, 曹敏建. 低钾胁迫对玉米苗期根系生长和钾吸收特性的影响[J].玉米科学, 2007, 15(2): 107-110.
Liu Y J, Tian X Y, Cao M J. The effects of low potassium stress on root growth and potassium absorption of maize seedling[J]. Journal of Maize Sciences, 2007, 15(2): 107-110.
[22] Zia-ul-Hassan, Arshad M. Cotton growth under potassium deficiency stress is influenced by photosynthetic apparatus and root system[J]. Pakistan Journal of Botany, 2010, 42(2): 917-925.
[23] Gerardeaux E, Jordan-Meille L, Constantin Jetal. Changes in plant morphology and dry matter partitioning caused by potassium deficiency inGossypiumhirsutumL.[J]. Environmental and Experimental Botany, 2010, 67(3): 451-459.
