饲料蛋白水平对泥鳅幼鱼生长和饲料利用的影响
2014-03-29叶文娟韩冬朱晓鸣杨云霞解绶启
叶文娟韩 冬朱晓鸣杨云霞解绶启
(1. 中国科学院水生生物研究所, 淡水生态与生物技术国家重点实验室, 武汉 430072; 2. 中国科学院大学, 北京 100049)
饲料蛋白水平对泥鳅幼鱼生长和饲料利用的影响
叶文娟1,2韩 冬1朱晓鸣1杨云霞1解绶启1
(1. 中国科学院水生生物研究所, 淡水生态与生物技术国家重点实验室, 武汉 430072; 2. 中国科学院大学, 北京 100049)
蛋白质是鱼类的第一营养要素, 是鱼类生长的物质基础, 鱼体组织干物质的65%—75%都由蛋白质组成。蛋白质在维持鱼体新陈代谢、促进鱼体生长、发育、繁殖等各阶段中都发挥着至关重要的作用。食物蛋白经消化吸收后以氨基酸的形式进入体内, 氨基酸参与体内绝大部分酶、含氮激素(如生长激素、胰岛素)和含氮维生素(大部分B族维生素)的合成[1]。摄食配合饲料是养殖鱼类获取蛋白质的最重要途径, 鱼用配合饲料的蛋白水平一般在25%—55%[2]。饲料蛋白水平是决定鱼类生长和饲料成本的关键因素, 蛋白源成本可占饲料总成本的50%以上。饲料蛋白水平过低会限制鱼类生长, 无法保证养殖鱼类在一定的时间内达到上市规格, 降低水产养殖的经济效益。过高的饲料蛋白水平并不能进一步提高鱼类的生长性能,反而会增加鱼体的氮代谢负荷。饲料中多余的蛋白质被鱼体分解用于机体供能而非用于体蛋白的储积, 从而降低了鱼体对饲料蛋白质的利用率[3—5]。为了保证良好的养殖经济效益, 人工配合饲料中的蛋白水平必须根据养殖鱼类的实际蛋白需求来进行合理的设定。鱼类对蛋白质的需求量通常包含两层含义: ①维持鱼体蛋白动态平衡即维持体内蛋白质现状所必需的蛋白质量, 即鱼类的维持蛋白需求量; ②鱼类达到最大生长或鱼体内蛋白质积蓄达到最大量时所需的最低蛋白质量, 即鱼类对饲料蛋白的最适需求量。营养学研究通常采用为期至少8周的生长实验来探讨鱼类对蛋白质的需求, 根据鱼类生长速度和饲料蛋白水平的关系(剂量—反应曲线)来确定鱼类的蛋白需求量[2]。
泥鳅(Misgurnus anguillicaudatus)隶属鲤形目鳅科泥鳅属, 是一种广布小型底栖杂食性鱼类。由于具有特殊的肠呼吸功能, 泥鳅耐低氧易运输, 环境适应能力强, 几乎能在各种水域中进行养殖。泥鳅的市场需求量比较稳定,出口需求量较大, 市场价格高, 经济效益高。但近年来随着化肥、农药、工业废水等污染加剧, 泥鳅的野外生态环境进一步恶化, 加上人类的过度捕捞, 导致野生泥鳅资源锐减。野生泥鳅的产量已经远远不能满足市场需求, 发展集约化养殖是满足人类需求的有效途径。配合饲料是发展集约化养殖的基础, 然而国内外关于泥鳅营养需求的研究很少, 数据不足以支持泥鳅专用配合饲料的开发。本实验旨在探讨饲料蛋白水平对泥鳅幼鱼生长和饲料利用的影响以及泥鳅幼鱼达到最大生长时的饲料最适蛋白添加量, 为开发泥鳅幼鱼专用配合饲料提供数据支持。
1 材料与方法
1.1 养殖条件及实验鱼
生长实验在室内塑料水族箱(水体体积为: 90 cm× 45 cm×15 cm)中进行, 静水养殖, 每天更换曝气自来水约1/3。实验用泥鳅幼鱼购自湖北省洪湖渔场, 在实验条件下驯养2周, 暂养期间每天投喂等量混合的实验饲料。实验开始前将暂养鱼集中禁食24h, 选择体质健壮、规格均匀的泥鳅, 随机放入21个养殖缸中, 每缸15尾, 平均体重(1.72±0.04 ) g。实验共设置7个处理, 每个处理3个重复。实验期间每天9:00和18:00投喂, 采用循环投喂模式, 投喂时在1.5h内分多次投入, 每次投喂量以实验鱼20min内能吃完为宜。实验结束后, 称一定量的饲料放入无鱼的实验缸中, 10min后回收于70℃烘干后称重, 计算饲料的溶失率以校正实际摄食量。实验期间水温变动范围是25—28 , pH℃ 为6.5左右, 氨氮≤0.5 mg/L , 余氯≤0.05 mg/ L, 光照周期为12L∶12D。
1.2 实验饲料
以白鱼粉和豆粕为主要蛋白源, 鱼油和大豆油为主要脂肪源设7组不同蛋白水平的等脂等能实验饲料。饲料蛋白水平实测值分别为22.82%、28.28%、33.24%、38.23%、42.71%、48.49%和51.97% (分别用23、28、33、38、43、48和52表示), 饲料蛋白能量比为(11.91—26.64) mg/kJ,实验饲料配方和基本生化组成见表1。所有实验原料过30目筛, 使用实验室小型制粒机(SLP-45, 中国上海渔业机械研究所)进行制粒, 粒径为1 mm。湿颗粒饲料置于烘箱中70℃烘至半干后, 使用密封袋分装后置于–20℃冰箱中保存备用。

表1 实验饲料配方和基本成分Tab. 1 Formulation and chemical composition of experimental diets
1.3 实验取样和样品分析
实验开始前, 随机取3组泥鳅幼鱼, 每组约20 g, 抹干称重后于−4℃冰箱中保存, 用于实验初始样本的基本组分分析。生长实验结束后经过24h禁食, 称重, 计算实验鱼的生长。每缸随机选取约10尾泥鳅, 抹干称重后于−4℃保存, 用于实验中终末样本的基本组分的测定。
实验饲料和鱼体的基本组分包括水分、灰分、粗蛋白和粗脂肪, 测定方法参照AOAC(1984)[6]。样品水分含量通过在 105℃烘箱中烘至恒重进行测定, 灰分含量通过在马弗炉中 550℃充分灼烧测定。粗蛋白含量采用凯氏定氮法, 使用全自动凯氏定氮仪(2300 Kjeltec Analyzer Unit, FOSS Tecator, Haganas, Sweden)测定样品含氮量, 再乘以6.25得蛋白含量。粗脂肪含量则以乙醚为溶剂采用索氏抽提仪(Soxtoc system HT6, Tecator, Haganas, Sweden)进行提取。样品能值用氧弹测热仪(Phillipson microbomb calorimeter, Gentry Instruments Inc., Aiken, USA)测定。每个样品至少测定两个平行样。
1.4 数据处理和统计分析
实验鱼存活率(Survival)、摄食率(Feeding rate, FR)、增重率(Weight gain, WG)、特定生长率(Specific growth rate, SGR)、饲料系数(Feed conversion ratio, FCR)、蛋白质储积率 (Protein retention efficiency, PRE)和能量储积率(Energy retention efficiency, ERE)的计算公式如下:
存活率(%)= 100×终末尾数/初始尾数
摄食率(% BW/d) = 100×摄食量/[养殖天数×(初始体重+终末体重)/2]
增重率(%)=100×(终末体重–初始体重)/初始体重
特定生长率(%/d)=100×((ln终末体重– ln初始体重 )/投喂天数)
饲料系数=摄食量/(终末体重–初始体重)
蛋白质储积率(%)=100×(终末鱼体蛋白质重–初始鱼体蛋白质重)/蛋白质摄入量
能量储积率(%) =100×(终末鱼体能值–初始鱼体能值)/能量摄入量
所有实验数据用Statistica 6.0软件进行单因素方差分析(One-way ANOVA), 差异显著时进行Duncan’s多重比较。根据增重率(WG)与饲料蛋白水平的相关性, 通过折线法(Broken-1ine)分析获得泥鳅幼鱼最大生长时的饲料蛋白需要量。
2 结果与讨论
2.1 存活、摄食和生长
饲料蛋白水平对泥鳅幼鱼存活、摄食、生长和饲料利用的影响见表2。经过8周的生长, 泥鳅幼鱼从平均初始体重1.72 g生长到 4.50 g。实验鱼存活率为82.2%—97.8%, 并随着饲料蛋白水平上升呈上升趋势(P<0.05)。实验鱼摄食率在低蛋白组(23%和 28%)最高(P<0.05), 在其余各处理组之间没有显著差异(P>0.05)。Wilson[1]研究表明鱼类摄食是为了满足自身对能量和营养素的需求, 在本实验中实验饲料均为等脂等能饲料, 低蛋白组实验鱼较高的摄食率可能是为了满足鱼体对营养素的需求而做出的补偿性调节。由于摄食率是单位体重的摄食量, 摄食率的变化既受到摄食量的影响, 同时也和各实验处理组生长差异相关。

表2 饲料蛋白水平对泥鳅幼鱼生长和饲料利用的影响 (平均值±标准误)Tab. 2 Effect of dietary protein on growth performance and feed utilization of juvenile Misgurnus anguillicaudatus (means ± SE)
泥鳅幼鱼的终末体重(FBW)、增重率(WG)和特定生长率(SGR)均随着饲料蛋白水平的升高而显著上升(P<0.05), 43%—52%饲料蛋白处理组的生长差异不显著(P>0.05)。在本实验中鱼类生长随着饲料蛋白水平的上升而先上升,然后进入平台期, 这一生长曲线符合鱼类和饲料蛋白水平之间的生长-剂量关系[7], 鲿在对短体下眼 (Horabagrus brachysoma)[8]、墨西哥原银汉鱼(Menidia estor)[9]和黑线鳕(Melanogrammus aeglefinus)[10]的蛋白需求研究中也发现类似的现象。在其他学者的一些研究中则发现高水平饲料蛋白会抑制鱼类生长, 生长曲线后期会随着饲料蛋白水平的上升而出现下降[11—13]。
基于增重率(WG)的折线模拟结果(图 1 )表明, 泥鳅幼鱼(1.72 g)达到最大生长的饲料蛋白水平为45.5%(占饲料干物质)。在其他研究中发现, 杂食性鲤(Cyprinus carpio)幼鱼阶段的蛋白需求量为 43%—47%[2], 草食性草鱼(Ctenopharyngodon idella)幼鱼阶段的饲料蛋白适宜含量为 41%—43%[14], 肉食性青鱼(Mylopharyngodon piceus)夏花(1—1.6) g的最适饲料蛋白含量为41%[15], 杂食性中华倒刺耙 (Spinibarbus sinensis)幼鱼的最适蛋白需求为39.6%—42.2%[16]。泥鳅属于鲤科杂食性鱼类, 本实验得出的泥鳅幼鱼的蛋白需要量和相近规格鲤的饲料蛋白需求值近似。在无鳞鱼的研究中发现, 黄颡鱼(Pelteobagrus fulvidraco)幼鱼(10 g)饲料蛋白需求约为 44%—45%[17],美洲鳗鲡(Anguilla rostrata)幼鱼(8 g)饲料蛋白需求约为47%[18], 长吻(Leiocassis longirostris)鱼种(12 g)最适饲料蛋白需求量为 46.9%—49.5%[19], 乌鳢(Channa striata)幼鱼(0.5 g)的最适蛋白需求为55%[20]。泥鳅幼鱼的蛋白需求和黄颡鱼幼鱼的饲料蛋白需求一致, 略低于肉食性的美洲鳗鲡、长吻和乌鳢。鱼类的饲料蛋白质需要量受多种因素的影响, 如鱼类的种类(品系)、规格、养殖环境水温、溶氧、盐度和饲料蛋白源、投喂水平等, 食性、习性和规格相近的鱼类具有更为接近的饲料蛋白需求值[21]。
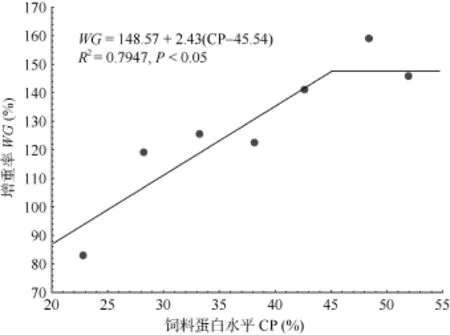
图1 饲料蛋白水平对泥鳅幼鱼增重率的影响Fig.1 Effect of dietary protein on weight gain (WG) of juvenile Misgurnus anguillicaudatus
2.2 饲料利用
在本实验中泥鳅幼鱼的饲料系数(FCR)随着饲料蛋白含量的增加而下降, 23%和28%饲料蛋白组的饲料系数较高(P<0.05), 其余各蛋白水平处理组之间饲料系数差异不显著(P>0.05)。摄食高蛋白饲料的实验鱼表现出较低的饲料系数, 意味着鱼类对高蛋白饲料具有较好的饲料利用, 在河豚(Takifugu rubripes)[22]、石斑鱼(Epinephelus coioides)[23]和胭脂鱼(Myxocyprinus asiaticus)[24]等的研究中也有类似的发现。饲料系数是衡量饲料品质最重要的指标之一, 在一定程度内提高饲料蛋白水平可以获得较低的饲料系数, 降低养殖成本。
鱼体蛋白储积率(PRE)随着饲料蛋白水平的上升而呈现下降趋势, 蛋白储积率在低饲料蛋白水平23%—38%没有显著差异(P>0.05), 显著高于高蛋白水平饲料处理组43%—52% (P<0.05)。当饲料蛋白水平过高时, 泥鳅幼鱼的蛋白储积率明显下降。这和很多其他鱼类的研究结果一致, 说明超过鱼类需求的饲料蛋白并不能有效的被应用于增加体蛋白的储积, 而是更多的被分解用作能量物质的消耗, 从而降低了饲料蛋白的利用率[22,25]。
实验饲料为等脂等能饲料, 蛋白水平对鱼体能量储积率的影响较小。泥鳅幼鱼的能量储积率(ERE)随着饲料蛋白水平呈现上升趋势, 但在33%—53%饲料蛋白水平之间没有显著差异(P>0.05)。在Kim和Lall[10]对黑线鳕(Melanogrammus aeglefinus L.)的研究和Sá, et al.[4]对重牙鲷(Diplodus sargus)的研究中, 能量储积率也都随着饲料蛋白水平的上升呈上升趋势但并没有显著性差异。
2.3 鱼体组成
泥鳅幼鱼的全鱼基本组分包括水分、灰分、粗蛋白和粗脂肪均没有受到饲料蛋白水平的显著影响(P>0.05,表3)。Lee和Kim[26]研究报道马苏大马哈鱼(Oncorhynchus masou Brevoort)的全鱼蛋白和灰分含量没有受到30%—50%饲料蛋白水平的影响, Lee和 Cho[27]报道大菱鲆(Scophthalmus maximus L.)的全鱼水分、灰分、蛋白和脂肪含量不受饲料蛋白水平(29%—57%)的影响。Martinez-Palacios, et al.[9]研究报道墨西哥原银汉鱼(Menidia estor)的全鱼水分、灰分、粗蛋白和粗脂肪含量也没有受到25%—55%饲料蛋白水平的影响。另外的一些研究则发现鱼体粗蛋白水平随饲料蛋白水平的上升而升高, 鱼体脂肪水平随着饲料蛋白水平的上升而下降[5,18,23]。
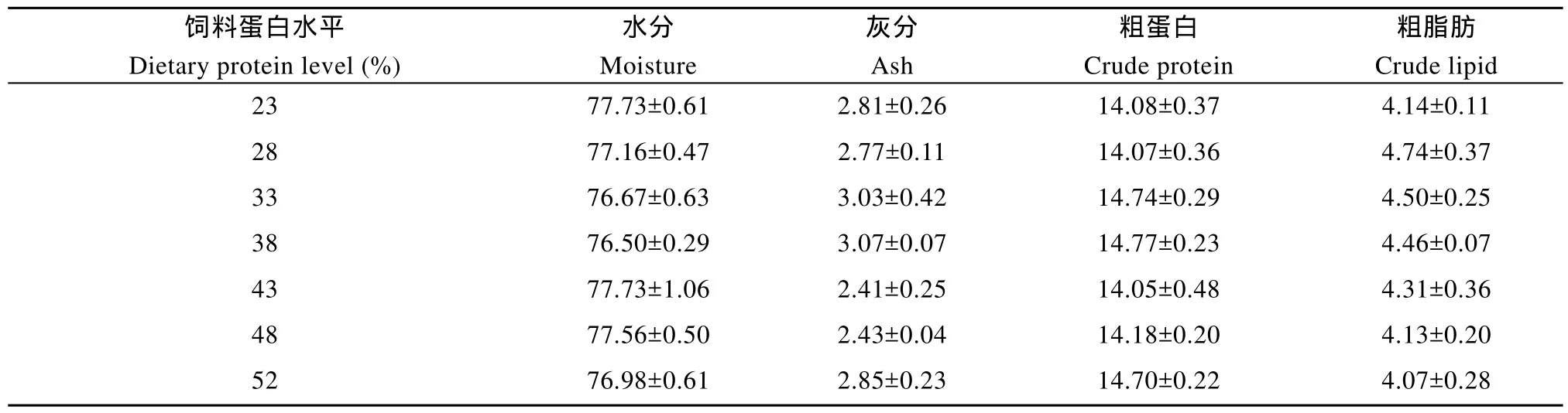
表3 饲料蛋白水平对泥鳅幼鱼全鱼组分的影响(平均值±标准误)Tab. 3 Effect of dietary protein on whole body composition of juvenile Misgurnus anguillicaudatus (means ± SE)
3 结论
以白鱼粉和豆粕为主要蛋白源, 饲料脂肪水平为11.66%—11.98%, 饲料蛋白能量比为11.91—26.64 mg/kJ,水温为 25—28℃的实验条件下, 通过增重率(WG)和饲料蛋白水平的折线法分析表明, 泥鳅幼鱼(1.72 g)达到最大生长的饲料蛋白水平为约占饲料干物质的45.5%。
[1] Wilson R P. Protein and amino acid requirements of fishes [J]. Annual Review of Nutrition, 1986, 6(1): 225—244
[2] National Research Council (NRC). Nutrient Requirements of Fish [M]. National Academy Press, Washington, DC. 1993, 16
[3] Shyong W J, Huang C H, Chen H C. Effects of dietary protein concentration on growth and muscle composition of juvenile Zacco barbata [J]. Aquaculture, 1998, 167(1): 35—42
[4] Sá R, Pousao-Ferreira P, Oliva-Teles A. Dietary protein requirement of white sea bream (Diplodus sargus) juveniles [J]. Aquaculture Nutrition, 2008, 14(4): 309—317
[5] Yang S D, Liou C H, Liu F G. Effects of dietary protein level on growth performance, carcass composition and ammonia excretion in juvenile silver perch (Bidyanus bidyanus) [J]. Aquaculture, 2002, 213(1):363—372
[6] AOAC. Official Methods of Analysis [A]. 14th edn. Washington D C: Association of Official Analytical Chemists. 1984, 152—163
[7] Robbins K R, Norton H W, Baker D H. Estimation of nutrient requirements from growth data [J]. The Journal of Nutrition, 1979, 109(10): 1710—1714
[8] Giri S S, Sahoo S K, Paul B N, et al. Effect of dietary protein levels on growth, feed utilization and carcass composition of endangered bagrid catfish Horabagrus brachysoma, (Gunther 1864) fingerlings [J]. Aquaculture Nutrition, 2010, 17(3): 332—337
[9] Martinez-Palacios C A, Rios-Duran M G, Ambriz-Cervantes L. Dietary protein requirement of juvenile Mexican Silverside (Menidia estor Jordan 1879), a stomachless zooplanktophagous fish [J]. Aquaculture Nutrition, 2007, 13(4): 304—310
[10] Kim J D, Lall S P. Effects of dietary protein level on growth and utilization of protein and energy by juvenile haddock (Melanogrammus aeglefinus) [J]. Aquaculture, 2001, 195(3): 311—319
[11] Jauncey K. The effects of varying dietary protein level on growth, food conversion, protein utilization and body composition of juvenile tilapia [J]. Aquaculture, 1982, 27(1): 43—45
[12] Elangovan A, Shim K F. Growth response of juvenile Barbodes altus fed isocaloric diets with variable protein levels [J]. Aquaculture, 1997, 158(3): 321—329
[13] Ozorio R O A, Valente L M P, Correia S, et al. Protein requirement for maintenance and maximum growth of two-banded seabream (Diplodus vulgaris) juveniles [J]. Aquaculture Nutrition, 2009, 15(1): 85—93
[14] Dabrowski K. Protein requirements of grass carp fry (Ctenopharyngodon idella Va1.) [J]. Aquaculture, 1977, 12: 63—73
[15] Yang G H, Li J, Guo L J, et al. Optimum level of protein in diet for black carp fingerlings [J]. Journal of Fisheries in China, 1981, 5(1): 49—55 [杨国华, 李军, 郭履骥, 等. 夏花青鱼饵料中的最适蛋白含量. 水产学报, 1981, 5(1): 49—55]
[16] Lin X Z, Xie X J, Luo Y P, et al. Dietary protein requirement for juvenile Spinibarbus sinensis [J]. Acta Hydrobiologica Sinica, 2009, 33(4): 674—681 [林小植, 谢小军, 罗毅平,等. 中华倒刺 鲃幼鱼饲料蛋白质需求量的研究. 水生生物学报, 2009, 33(4): 674—681]
[17] Gan L, Yuan H. Nutrient requirements of yellow catfish, Pelteobagrus fulvidraco: a review [J]. China Fishery, 2006, 9: 62—64 [甘炼, 袁华. 黄颡鱼营养需求研究进展. 中国水产, 2006, 9: 62—64]
[18] Tibbetts S M, Lall S P, Anderson D M. Dietary protein requirement of juvenile American eel (Anguilla rostrata) fed practical diets [J]. Aquaculture, 2000, 186(1): 145—155
[19] Zhang Z Y, Zhang J T, Chen X J. On protein requirement of Leiocassis longirostris juvenile [J]. Southwest China Journal of Agriculture Science, 1990, 3(4): 87—90 [张泽芸, 张季涛,陈先均. 长吻(Leiocassis longirostris)鱼种阶段蛋白质最适需求量的研究. 西南农业学报, 1990, 3(4): 87—90]
[20] Mohanty S S, Samantaray K. Effect of varying levels of dietary protein on the growth performance and feed conversion efficiency of snakehead Channa striata fry [J]. Aquaculture Nutrition, 1996, 2(2): 89—94
[21] Qian X Q, Cui Y B, Xie S Q, et al. A review of dietary protein requirement for aquaculture fishes [J]. Acta Hydrobiologica Sinica, 2002, 26(4): 410—416 [钱雪桥, 崔奕波, 解绶启, 等. 养殖鱼类饲料蛋白需要量的研究进展. 水生生物学报, 2002, 26(4): 410—416]
[22] Kim S S, Lee K J. Dietary protein requirement of juvenile tiger puffer (Takifugu rubripes) [J]. Aquaculture, 2009, 287(1): 219—222
[23] Luo Z, Liu Y J, Mai K S, et al. Optimal dietary protein requirement of grouper Epinephelus coioides juveniles fed isoenergetic diets in floating net cages [J]. Aquaculture Nutrition, 2004, 10(4): 247—252
[24] Yuan Y C, Gong S Y, Luo Z, et al. Effect of dietary protein to energy ratios on growth and body composition of juvenile Chinese sucker, Myxocyprinus asiaticus [J]. Aquaculture Nutrition, 2010, 16(2): 205—212
[25] Shiau S Y, Huang S L. Optimum dietary protein level for hybrid tilapia (Oreochromis niloticus × O. aureus) reared in seawater [J]. Aquaculture, 1989, 81(2): 119—127
[26] Lee S M, Kim K D. Effects of dietary protein and energy levels on the growth, protein utilization and body composition of juvenile masu salmon (Oncorhynchus masou Brevoort) [J]. Aquaculture Research, 2001, 32(s1): 39—45
[27] Lee J K, Cho S H, Park S U, et al. Dietary protein requirement for young turbot (Scophthalmus maximus L.) [J]. Aquaculture Nutrition, 2003, 9(4): 283—286
EFFECT OF DIETARY PROTEIN LEVEL ON GROWTH AND FEED UTILIZATION OF JUVENILE MISGURNUS ANGUILLICAUDATUS
YE Wen-Juan1,2, HAN Dong1, ZHU Xiao-Ming1, YANG Yun-Xia1and XIE Shou-Qi1
(1. State Key Laboratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China)
泥鳅; 蛋白需求; 饲料利用
Misgurnus anguillicaudatus; Protein requirement; Feed utilization
S965.1
A
1000-3207(2014)03-0571-05
10.7541/2014.80
2013-04-10;
2013-12-25
国家自然科学基金专项基金(31123001)资助
叶文娟(1986—), 女, 重庆璧山人; 博士; 研究方向为鱼类生理生态学。E-mail: yewenjuan1986@yahoo.com.cn
解绶启(1969—), 男, 研究员; E-mail: sqxie@ihb.ac.cn
