咸宁地区部分桂花品种的ISSR分析
2014-03-27范付华夏辉王振启陈洪国
范付华,夏辉,王振启,陈洪国
(1.湖北科技学院核技术与化学生物学院,湖北咸宁437100;2.咸宁职业技术学院,湖北咸宁437000)
0 引言
桂花(Osmanthus fragrans Lour.)为木犀科(Oleaceae)木犀属(Osmanthus Lour.)植物,是我国特有的常绿小乔木或灌木经济树种,原产于我国长江流域至华南、西南各地,在长期的人工栽培和演化过程中,形成了历史上著名的桂花五大产区:江苏苏州、浙江杭州、湖北咸宁、四川成都和广西桂林.桂花栽培范围广,自然环境复杂,由此也产生了许多的性状变异,形成了丰富的品种资源.长期以来,科研工作者以形态学基础对桂花品种进行分类,但由于对品种概念、分类方法及标准的认识不尽相同,20世纪80年代以后,有多人提出了不同的分类标准[1-4],导致了部分桂花品种分类较为混乱,记载不统一.咸宁作为“中国桂花之乡”,从桂花数量、产量到品种都保持全国领先地位,对咸宁桂花遗传多样性的研究将有助于其品种及其相关产业的长远发展.
分子标记技术已广泛用于植物遗传多样性分析、亲缘关系和品种鉴定。尽管利用分子标记技术研究桂花亲缘关系、品种分类等已见诸多报道[5-10],其中咸宁地区部分种质亲缘关系的分析也有涉及[7,9],但所选品种较少,不足以反映咸宁桂花品种遗传多样性的实质.本研究中以咸宁地区48份桂花主要栽培品种为供试材料,利用ISSR技术分析鉴定,旨在进一步揭示咸宁地区桂花亲缘关系和品种多样性,为地方桂花资源保护和遗传改良提供更充实的理论基础.
1 材料与方法
1.1 材料 选取咸宁地区48份桂花主要栽培品种作为研究对象。试验材料均采集于湖北咸宁潜山国家森林公园。采集幼嫩叶片,液氮处理后迅速放入-80℃超低温冰箱保存.试验所用的材料如表1.

表1 供试桂花品种名称及其资源类别
1.2 DNA的提取及检测 取超低温保存下的幼嫩桂花叶片,利用DNA secure Plant Kit(DP320-02)试剂盒(天根)提取DNA.经1%琼脂糖凝胶电泳检测其质量完好后,用TE缓冲液稀释DNA至约20ng/μL,放入-20℃冰箱备用.
1.3 PCR扩增及检测 参照范付华[11-12]方法,稍作修改.选取UBC公司公布的50条ISSR引物序列.筛选出合适的引物及每个引物的最佳退火温度.最适ISSR扩增反应体系为(15μL):7.5μL Mix,1.2μL引物(10μmol/L),1.5μL模板DNA和4.8μL ddH2O.PCR反应程序为94℃预变性4min;94℃变性1min,复性1min,温度随引物而定(表2),72℃延伸1min,30个循环;最后72℃延伸7min.ISSR扩增产物,用2%琼脂糖凝胶电泳检测.
1.4 数据处理及分析 每个引物重复扩增2次,选择可重复且清晰可见的谱带进行统计.将扩增所得到的谱带按“引物号-片段长度”进行指纹记录,以按谱带的有无分别赋值“1”和“0”建立数据库,利用NTSYSpc 2.10e软件计算供试材料间的相似性系数,并构建亲缘关系树状图.
2 结果与分析
2.1 ISSR扩增多态性 采用筛选出的11条扩增稳定性高、谱带清晰的ISSR引物,对48份供试桂花材料进行PCR扩增分析.结果表明(表2):所有11条引物共扩增出清晰可辨的谱带81条,平均每个引物可扩增7.4条,其中多态性谱带共70条,多态性比率达到86.4%;扩增出谱带最多的引物为825,共10条带;819扩增的谱带最少,仅为5条;823、842和844扩增的谱带多态性比率都达到了100%;引物891获得多态性谱带的比率最低,仅为71.4%.所有引物扩增出的DNA片段大小大多在250~2 000bp之间,并且谱带清晰(图1),可以用于桂花亲缘关系及遗传多样性分析.上述ISSR引物检测到较高的多态性谱带比率,说明了桂花品种间的遗传变异较大.
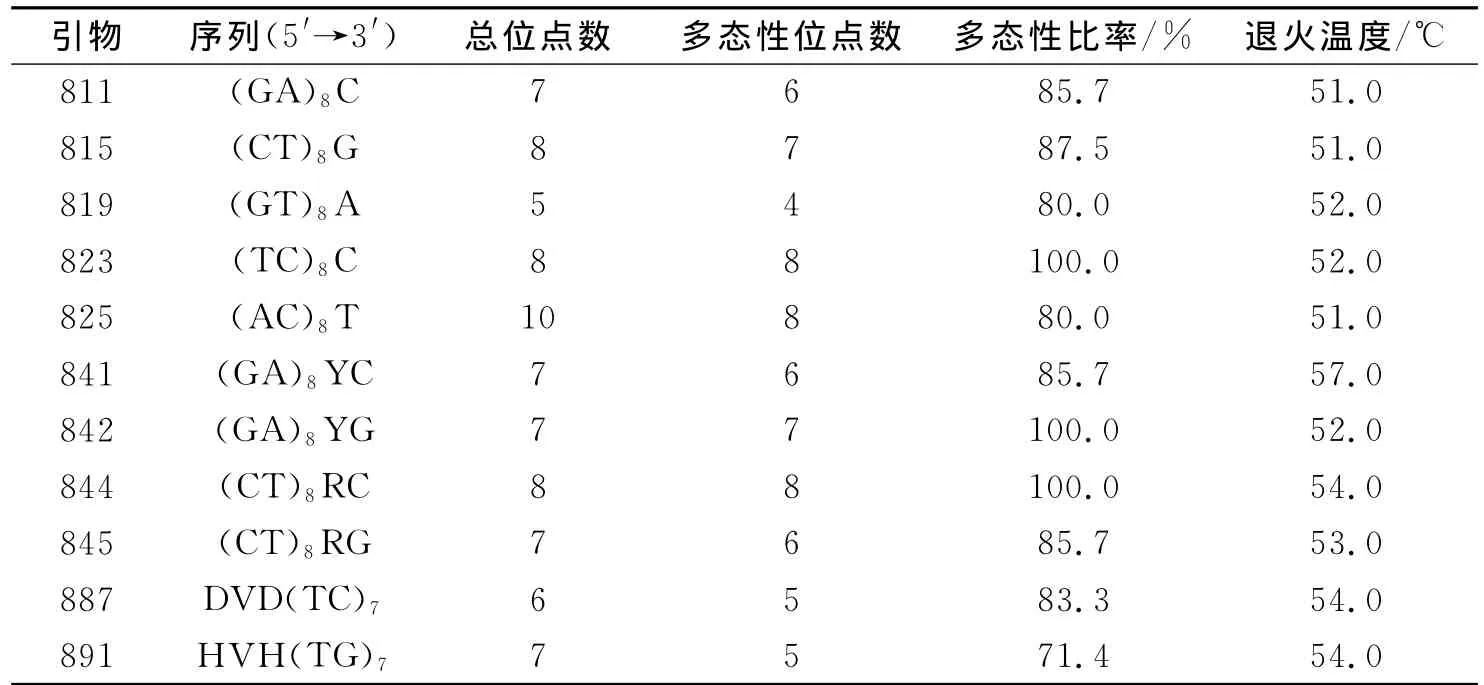
表2 用于PCR扩增的ISSR引物及其扩增结果

图1 引物842扩增48份供试材料的电泳图样品1~48的含义同表1
2.2 遗传相似系数分析 供试材料的SIMQUAL相似系数在0.46~0.97之间.29号“满条红”在湖北咸宁地区称作“红花”,自古享有盛名,是当地常见的品种,其与同为丹桂品种群的22号“大花丹桂”的相似系数最大,达到了0.97.相似系数最小的是来自四季桂品种群的3号和6号“小叶四季桂”和“天女散花”与丹桂品种群的24号“平脉红”,仅为0.46,说明这两个品种间具有较大的遗传差异,推测亲缘关系较远.对各个品种群内样品之间的遗传相似系数分析,四季桂品种群平均值为0.79,丹桂品种群为0.76,银桂品种群为0.75,金桂品种群为0.74.由此可知,咸宁地区四季桂品种群内种质间的遗传差异相对较小,遗传多样性不如其他品种群,其中金桂品种群遗传多样性相对最为丰富.
2.3 供试桂花品种的聚类 采用UPGMA法对供试桂花品种进行聚类分析,建立树状分支图(图2).48份材料在以相似系数0.75为阈值的情况下被分为5类,第Ⅰ大类包含所有的9个四季桂品种;第Ⅱ类全部为秋季开花的22个品种(包括银桂、丹桂和金桂品种群);丹桂12个品种及金桂1个品种聚为第Ⅲ大类.金桂品种群36号“大叶黄”与供试材料遗传距离较远,单独聚为第Ⅳ类;第Ⅴ大类中包括2个丹桂品种“平脉红”、“朱砂丹桂”和1个金桂品种“丛中笑”.从聚类图中可见,四季桂品种群与秋桂品种可以明显分开,而四季桂较之其他品种群童期短、花期长,一年多次开花,说明桂花亲缘关系的远近与其开花习性具有一定的相关性.由图2可知,四季桂品种单独被聚为1类,银桂品种群10个品种聚为1类,丹桂品种群15个品种分别聚入不同的3类,而金桂品种群14个品种被聚入不同的4类.反映了咸宁地区金桂品种群的遗传多样性比其他品种群丰富,这与遗传相似系数的结论一致.
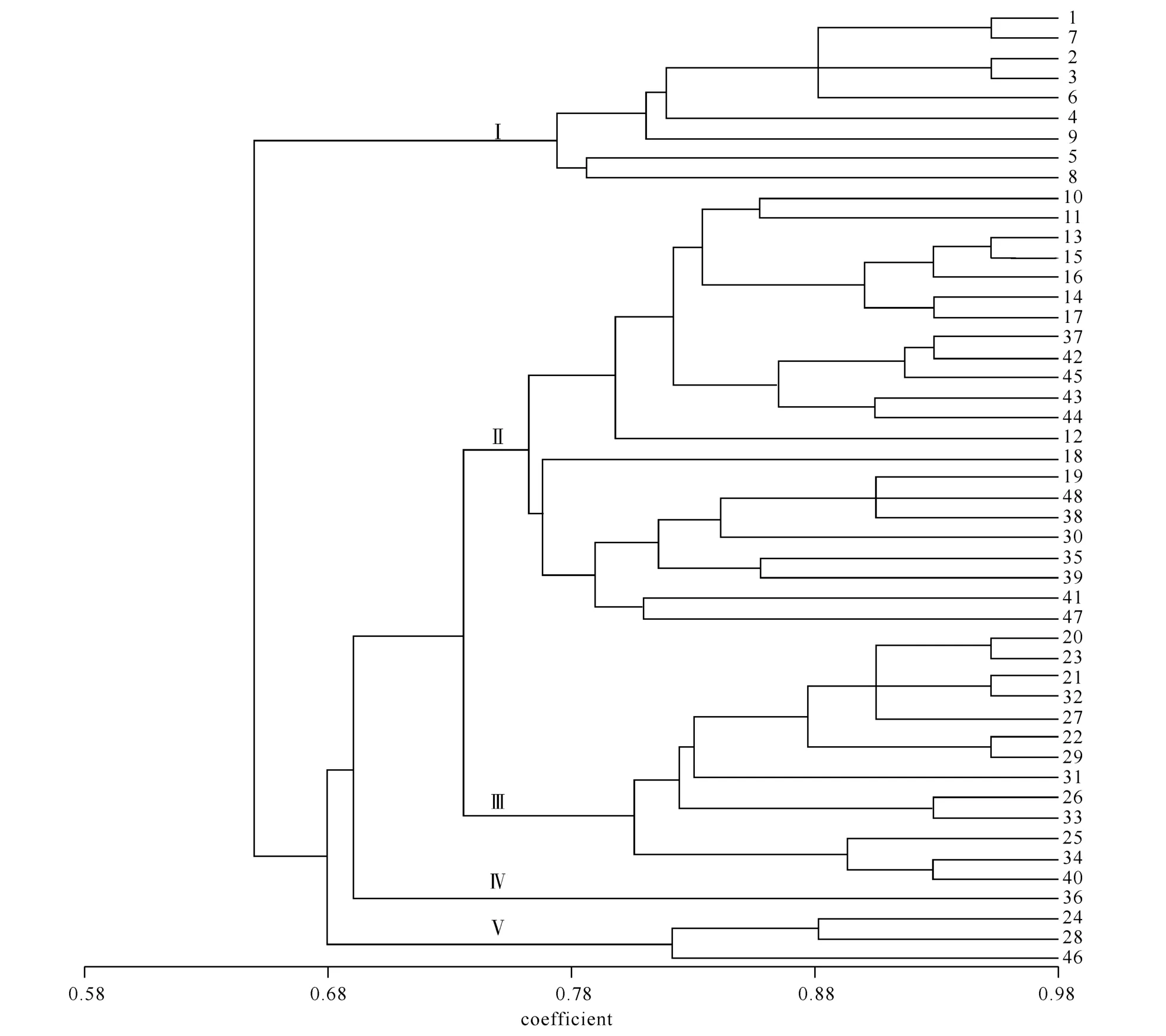
图2 48个咸宁桂花品种的UPGMA聚类分析图
3 讨论
从ISSR引物扩增谱带的多态性比率和供试品种间遗传相似性系数可知,咸宁地区桂花品种间具有较大变异,存在丰富的遗传多样性,这与长期的自然选择和人工干预密切相关.咸宁也以其资源的多样性、栽培的广泛性、桂花产量和品质的优良性于2000年被国家命名为唯一的“中国桂花之乡”.桂花各品种群平均遗传相似性系数依次为四季桂品种群 > 丹桂品种群 > 银桂品种群 > 金桂品种群,这与李梅等[8]研究结果相似又不完全相同,仅在银桂和金桂品种群间有所区别.种群间遗传多样性比率显示咸宁地区桂花大部分种内变异存在于秋桂(金桂、银桂、丹桂)之间.桂花品种资源遗传多样性的分析,为当地种质资源管理和育种提供重要参考.
聚类分析结果(图2)显示,多季开花的四季桂品种全部聚在一起,与3个秋桂类品种群遗传距离较远.基于ISSR分子标记的聚类结果与按传统形态学的一级分类标准结果相符,说明开花习性是桂花品种划分的依据之一,具有重要的分类学意义.由聚类图可见,秋桂中各个品种群的大多数品种可以先聚在一起,部分不同品种群的品种也会聚为一个分支,但多为花色相近的品种,特别是花色为橙红色的丹桂品种群的品种几乎聚在1类,而其他品种也是先聚为同一品种群后再与别的品种群有所交集.这与赵小兰等[13]的同工酶分析、尚富德等[5]的RAPD分析、韩远记等[7]的AFLP分析,以及李梅等[8]的ISSR分析的结论基本相似,说明将桂花花色作为品种分类标准具有一定合理性,但本研究中金桂与银桂、金桂与丹桂部分品种往往也聚在一起,因此,仅以花色作为桂花品种分类的二级标准并不能完全反映品种间的亲缘关系.综上所述,今后应以传统形态分类学为基础,结合酶学、分子标记及解剖学等多种分类方法,相互印证[14],以期提出更科学合理的桂花品种分类系统.
ISSR是Zietkiewicz等[15]创建起来的以PCR技术为基础且不要求预知基因组序列信息的分子标记,该项技术兼具RAPD和SSR优点,DNA用量少、多态性水平高、可重复性好、稳定性高,是一种快速、可靠、可提供丰富位点信息的DNA指纹技术[16],已在多种果树[17]、林木[18]上得到广泛应用,在亲缘关系和品种鉴定方面有较高的可靠性.本研究中发现,ISSR标记揭示出桂花不同品种及不同品种群间的多态性较丰富,存在较明显的遗传差异.并非每一个供试桂花品种都具有特征标记,但通过不同的ISSR引物组合,能将所有样品区分开,且与蔡宇良等[19]对樱桃种质资源的分析中认为的随着引物数量增加而区分更多材料的结论一致.对供试材料相似性系数及聚类显示,丹桂品种群中22号“大花丹桂”和29号“满条红”遗传差异极小,本研究中可视为同一品种,对于是否是引物太少导致的此类结果,将结合品种的形态特征,利用更多的分子标记方法进行深入研究.
[1]朱长山,李瑞符,袁建都,等.河南桂花品种的分类研究[J].河南农业大学学报,1992,26(2):194-201.
[2]刘玉莲.桂花品种分类及木犀属种质资源的利用[J].植物资源与环境,1993,2(2):44-48.
[3]高本年,张思平,曹广树.对桂花品种6级分类的探讨[J].江苏林业科技,1997,24(3):22-24.
[4]向其柏,刘玉莲.中国桂花品种图志[M].杭州:浙江科学技术出版社,2008.
[5]尚富德,伊艳杰,张彤.河南17个桂花品种的RAPD分析[J].园艺学报,2004,31(5):685-687.
[6]邱英雄,胡绍庆,陈跃磊,等.ISSR-PCR技术在桂花品种分类研究中的应用[J].园艺学报,2004,31(4):529-532.
[7]韩远记,董美芳,袁王俊,等.桂花品种资源的遗传多样性分析[J].植物学通报,2008,25(5):559-564.
[8]李梅,侯喜林,单晓政,等.部分桂花品种亲缘关系及特有标记的ISSR分析[J].西北植物学报,2009,29(4):674-682.
[9]胡春芳,袁录霞,张凡,等.咸宁部分桂花品种亲缘关系的SRAP分析[J].湖北农业科学,2012(9):1907-1911.
[10]Duan Y F,Wang X R,Xiang Q B,et al.Genetic diversity of androdioecious Osmanthus fragrans(Oleaceae)cultivars using microsatellite markers[J].Applications in Plant Sciences,2013,1(6):1200092.
[11]范付华,杨勇胜,李庆宏,等.利用ISSR分析技术鉴定贵州枇杷新种质的研究[J].北京林业大学学报,2012,34(4):52-57.
[12]范付华,宋莎,洪怡,等.贵州野生枇杷资源遗传多样性的ISSR分析[J].中国南方果树,2012,41(3):36-39.
[13]赵小兰,姚崇怀,王彩云.桂花品种的同工酶研究[J].华中农业大学学报,2000,19(6):595-599.
[14]楚爱香,汤庚国.我国观赏植物的品种分类方法[J].林业科技开发,2008,22(4):1-5.
[15]Zietkiewiez E,Rafalski A,Labuda D.Genome fingerprinting by simple sequence repeat(SSR)-anchored polymerase chain reaction amplification[J].Genomics,1994,20:176-183.
[16]Bornet B,Branchard M.Nonanchored inter simple sequence repeat(ISSR)markers:reproducible and specific tools for genome fingerprinting[J].Plant Molecular Biology Reporter,2001,19:209-215.
[17]张青林,罗正荣.ISSR及其在果树上的应用[J].果树学报,2004,21(1):54-58.
[18]杨玉玲,马祥庆,张木清.ISSR分子标记及其在树木遗传育种研究中的应用[J].亚热带农业研究,2006,2(1):18-24.
[19]蔡宇良,曹东伟,李珊,等.甜樱桃品种及其砧木的RAPD分析[J].西北植物学报,2006,26(6):1125-1132.
