石羊河下游退耕区次生草地自然恢复过程及土壤酶活性的变化
2014-03-26王理德姚拓何芳兰韩福贵郭春秀王方琳魏林源
王理德,姚拓,何芳兰,韩福贵,郭春秀,王方琳,魏林源
(1.甘肃农业大学草业学院,甘肃 兰州730070;2.甘肃民勤荒漠草地生态系统国家野外观测研究站,甘肃 民勤733300;3.甘肃省荒漠化与风沙灾害防治国家重点实验室培育基地,甘肃 武威733000;4.草业生态系统教育部重点实验室,甘肃 兰州730070)
退耕区是绿洲农业中不再实施耕作措施的这部分土地所占的区域(空间与面积)。主要是由于水土流失严重、粮食产量低而不稳、风沙危害严重、盐渍化程度高或者水资源缺乏等原因,不宜作为耕地,从而退出耕种的坡耕地、沙化地或者其他耕地。
农业大词典对次生草地的定义是在农田退耕放荒、森林采伐、草原放牧、割草、火烧、病虫害等外界因素作用后,自然演替形成草本植物群落为主的草地。在中国北方,干旱条件下发育形成的次生草地,是真旱生的多年生草本植物占优势、旱生小半灌木起明显作用的植被性草地,也可称为荒漠草原或漠境草原[1]。
次生草地土壤是一个不断进行着复杂生物化学反应的体系,土壤酶是土壤中的生物催化剂,参与许多重要的生物化学过程,与土壤肥力的形成和转化有密切关系[2-9]。土壤酶活性的研究作为土壤肥力指标而受到土壤学家的普遍重视[2,10]。土壤酶在土壤生物化学循环中具有重要地位,是土壤功能的直接体现。随着土壤酶的研究进展,土壤酶活性作为农业土壤质量和生态系统功能的生物活性指标已被广泛研究[11-14]。高寒灌丛草地土壤酶季节性动态特征[15]及高寒草甸草场不同载畜量下土壤酶与土壤肥力的关系研究[3]也有报道,但在西北干旱条件下,退耕区次生草地自然恢复过程中土壤酶活性变化的研究至今未见报道。
石羊河流域中下游地区,曾经是植被茂密、物种丰富、水草特别丰美的大片绿洲[16]。但是,由于石羊河上游农业生产规模不断扩大对水资源的需求逐年增加,流入石羊河中下游绿洲区内的水量急剧减少[17-18]。为了维持原有的生产、社会方式和保证粮食的基本需求,在地表水量急剧减少的情况下,一度靠过量开采地下水来满足灌溉需水[19-20]。在过去30多年中,地下水位由2m左右下降到现在的10m多,部分地区达到几十米,造成天然植被死亡、尾闾湖沼干涸、盐碱化和沙漠入侵等[21],造成土地大面积弃耕而撂荒,成为次生草地。据统计,民勤县用河水灌溉播种的面积,由历史上的6万多hm2减少至1995年的2.7万hm2,坝区2万多hm2耕地原可在苗期利用河水灌溉1~2次,现在已全部改为井灌。湖区历史上延续下来的约0.67万hm2水歇地因缺水全部弃耕;1.2万hm2耕地改为轮休种植或部分弃耕。民勤全县近8万hm2耕地,1995年实际播种面积只有4万余hm2。近年来在实施国家生态建设项目《甘肃石羊河流域重点治理规划》的过程中,政府采取“关井压田”和退耕政策,这样退耕区次生草地的面积又有所增加,这部分次生草地如果不及时保护,长时间大面积撂荒,就会风蚀沙化,成为新的沙尘源,形成恶性循环,使次生草地土壤生态系统遭到破坏,其结果加剧和导致石羊河中下游成为全国的沙尘源区和特级告急的生态危机区,不利于石羊河流域综合治理工程的实施。因此,如何保护石羊河流域中下游绿洲退耕区次生草地,促进该地区生态环境的保护、修复和改善是亟待解决的新课题。
本文采用时空替代法,对石羊河下游不同年限(1~31a)退耕区次生草地自然恢复过程中的群落类型及演替规律进行了调查与分析,并对植被自然恢复过程中土壤酶的活性变化及分布特征进行研究,旨在为退耕区次生草地自然恢复重建和保护提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于石羊河下游的西渠镇,海拔1300~1311m,地理坐标为39°01′30″~39°03′28″N、103°35′57″~103°37′56″E,研究区均为退耕区次生草地,附近居民已经搬迁或者移民。年日照时数2832.1h;年均温度7.4℃,极端最高气温38.1℃,极端最低气温-28.8℃,昼夜温差大;年均降水量110mm,年内分布不均匀,73%的降雨量集中在7-9月;年均蒸发量2644mm;年平均风速2.3m/s;土壤类型为灰棕漠土。灌木种主要有盐爪爪(Kalidiumfoliatum),苏枸杞(Lyciumruthenicum),红砂(Reaumuriasoongorica)等;草本植物有藜(Chenopodiumalbum),骆驼蒿(Peganumnigellastrum),盐生草(Halogetonarachnoideus),骆驼蓬(Peganumharmala),田旋花(Convolvulusarvensis)等。
1.2 样地布设与植被调查
1.2.1 样地布设 2011年11月,走访调查和查阅西渠镇煌辉村与志云村退耕土地使用记录。采用空间替代时间的方法,在集中连片分布的退耕区次生草地上,选择该区域植物生长均匀、微地形差异较小。并且,土壤没有因自然因素而导致地形的变迁或因人为因素而引起的土壤物质再分配的地段,在保证样地沙土母质基本相同的情况下,分别选择退耕年限为1,2,3,4,5,8,15,24和31a的样地9个(表1),并用 GPS定位,样地面积为1hm2。在每个样地中以S形方法布设10m×10m样方5个,在每个样方内的固定样点上做好标记。试验区各样地之间相对高差只有9m(表1),海拔对植被演替不会造成影响。

表1 不同退耕年限样地状况Table 1 The status of abandoned lands sample with different durations
1.2.2 植被调查 2012年6月27-30日,调查每个样地内5个10m×10m的灌木样方中每种灌木的个体数、高度、冠幅及盖度等指标;再在每个10m×10m的灌木样方内四角及中心各设置1个1m×1m草本样方,分别记录每种草本的个体数、高度与盖度。
1.3 土壤样品采集及土壤酶活性测定
1.3.1 土壤样品采集 2012年6月28-29日,在不同年限退耕区次生草地样方周边采用剖面法和5点混合法分别采集30~40cm、20~30cm、10~20cm、0~10cm深度的土壤样品,设3次重复。每个样品取重1kg左右,装入布袋中,带回实验室,用于土壤酶活性的测定。
1.3.2 土壤酶活性测定 过氧化氢酶活性的测定采用容量法[14]。取5g风干土置于100mL三角瓶中,注入40 mL蒸馏水和5mL 0.3%过氧化氢溶液。将三角瓶放在往复式振荡机上,以120r/min振荡30min。而后加入5 mL 1.5mol/L的硫酸,以稳定未分解的过氧化氢。再将瓶中悬液用慢速型滤纸过滤。然后吸取25mL滤液,用0.02mol/L高锰酸钾滴定至淡粉红色为终点。
结果计算:用于滴定土壤滤液所消耗的高锰酸钾量(毫升数)为B,用于滴定25mL原始的过氧化氢混合液所消耗的高锰酸钾量(毫升数)为A。(A-B)×T即为过氧化氢酶活性。以30min后1g土壤的0.02mol/L高锰酸钾的毫升数表示。T为高锰酸钾滴定度的矫正值,即高锰酸钾标定后的真实浓度与试验所需配制的高锰酸钾的浓度(即0.02mol/L)的比值。
脲酶活性的测定采用靛酚蓝比色法[22]。标准曲线绘制:吸取稀释的10μg/mL氮的标准液1,3,5,7,9,11,13mL,移于50mL容量瓶中,然后加蒸馏水至20mL。再加4mL苯酚钠溶液和3mL次氯酸钠溶液,随加随摇匀。20min后显色,定容。1h内在分光光度计上于波长578nm比色。根据光密度值与溶液浓度绘制标准曲线。
取5g风干土置于50mL三角瓶中,加1mL甲苯。15min后加入10mL 10%尿素液和20mL pH 6.7柠檬酸盐缓冲液。摇匀后在37℃恒温箱中培养24h。过滤后取3mL滤液注入50mL容量瓶中,然后按绘制标准曲线显色方法进行比色测定。
结果计算:脲酶活性以24h后1g土壤中NH3-N的毫克数表示。
NH3-N(mg)=a×V×n/m,a为从标准曲线中查得的 NH3-N毫克数;V为显色液体积(50mL);n为分取倍数;m为烘干土的质量。
蔗糖酶活性采用3,5-二硝基水杨酸比色法[22]。标准曲线绘制:取0.01~0.5mg/mL不同浓度的葡萄糖工作液1mL,并按与测定蔗糖酶活性同样的方法进行显色,比色后以光密度值为纵坐标,葡萄糖浓度为横坐标绘制成标准曲线。
取5g风干土置于50mL三角瓶中,注入15mL 8%蔗糖溶液,5mL pH 5.5磷酸缓冲液和5滴甲苯。摇匀混合物后,放入恒温箱,在37℃下培养24h。到时取出,迅速过滤,从中吸取滤液1mL,注入50mL容量瓶中,加3,5-二硝基水杨酸,并在沸腾的水浴锅中加热5min,随后将容量瓶移至自来水流下冷却3min,溶液因生成3-氨基-5-硝基水杨酸而呈橙黄色,最后用蒸馏水稀释至50mL,并在分光光度计上于波长508nm处进行比色。
结果计算:蔗糖酶活性以24h后1g土壤葡萄糖的毫克数表示。
葡萄糖(mg)=a×V×n/m,a为从标准曲线中查得的葡萄糖毫克数;V为显色液体积(50mL);n为分取倍数;m为烘干土的质量。
磷酸酶活性用磷酸苯二钠比色法[22]。标准曲线绘制:取1,3,5,7,9,11和13mL酚工作液,置于50mL容量瓶中,每瓶加入5mL硼酸盐缓冲液和4滴氯代二溴对苯醌亚胺试剂,显色后稀释至刻度,30min后比色测定。以光密度为纵坐标,浓度为横坐标绘制成标准曲线。
取5g风干土置于200mL三角瓶中,加2.5mL甲苯,轻摇15min后加入20mL 0.5%磷酸苯二钠(测定酸性磷酸酶用醋酸盐缓冲液;碱性磷酸酶用硼酸盐缓冲液;中性磷酸酶用柠檬酸盐缓冲液。0.5%磷酸苯二钠溶液必须用相应的缓冲液配制),仔细摇匀后放入恒温箱,在37℃下培养24h。后于培养液中加100mL 0.3%硫酸铝溶液并过滤。吸取3mL滤液于50mL容量瓶中,然后按绘制标准曲线所述方法显色。用硼酸缓冲液时,呈蓝色,在分光光度计上于660nm处比色。
结果计算:磷酸酶活性,以24h后1g土壤中释出的酚的毫克数表示。
酚(mg)=a×V×n/m,a为从标准曲线中查得的酚毫克数;V为显色液体积(50mL);n为分取倍数;m为烘干土的质量。
1.4 数据处理
首先使用Excel软件进行统计整理,然后将整理好的数据用SPSS 16.0软件进行分析处理。对不同年代退耕样地土壤酶活性差异显著性采用单因素方差分析(one-way ANOVA)检验。
2 结果与分析
2.1 不同年限退耕区次生草地植被自然恢复变化特征
石羊河下游退耕区次生草地在31a的植被恢复过程中,9个样方内共出现植物34种,不同退耕年代物种组成存在很大差异。退耕1a的样方内出现22种植物,主要以黎、田旋花、五星蒿等一年生草本植物为主(表1);退耕2a的样方内出现17种植物,群落中一年生草本植物种类明显减少,苦苣菜、独行菜、骆驼蒿等多年生草本成为优势种;退耕3~5a的样方内物种数量减少为10种左右,一年生草本植物逐渐消失,顶羽菊、盐生草、骆驼蒿、骆驼蓬等成为群落优势种;退耕8~15a的样方内物种数量由12种减少为9种,与退耕3~5a样方内的物种数量相比较,变化不大,但优势种逐渐由木本植物代替草本植物;退耕24a的样方内,物种数量只有6种,群落中草本植物只有少量的芦苇(Phragmitesaustralis),其余都是灌木植物,盐爪爪、苏枸杞等成为群落优势种;退耕31 a后的样方内,植物种类只有5种,草本植物全部退出,形成以苏枸杞和盐爪爪为优势种的灌木植被群落。
退耕区次生草地自然恢复的31a间,退耕区次生草地上的物种构成表现为多数种属于少数科(藜科、禾本科、蒺藜科3科共出现18种植物占物种总数的52.94%),少数种属于多数科(十字花科、茄科、旋花科等11科只有16种植物,占物种总数的47.06%)。并且,很多种为单科单属种(蓼科、蓝雪科、毛茛科等7科,均只有一种植物出现),这一现象与杨自辉[23]、彭鸿嘉等[24]和何芳兰等[25]研究相一致,也符合西北荒漠区的植物区系特征[26]。
2.2 不同退耕年限次生草地土壤酶活性与分布特征
2.2.1 过氧化氢酶 过氧化氢酶活性均随土层深度的增加而显著降低,表层(0~10cm)过氧化氢酶活性显著高于其他各土层(表2)。0~10cm,10~20cm,20~30cm,30~40cm土层两两之间酶活性大部分差异均达到显著水平(P<0.05),除退耕1,3与5a外,其余退耕年限0~10cm土层中过氧化氢酶活性与10~20cm,20~30cm,30~40cm土层间的差异均达到极显著的水平(P<0.01)。另外,表层过氧化氢酶活性在4层该酶活性中占有较大的比例。
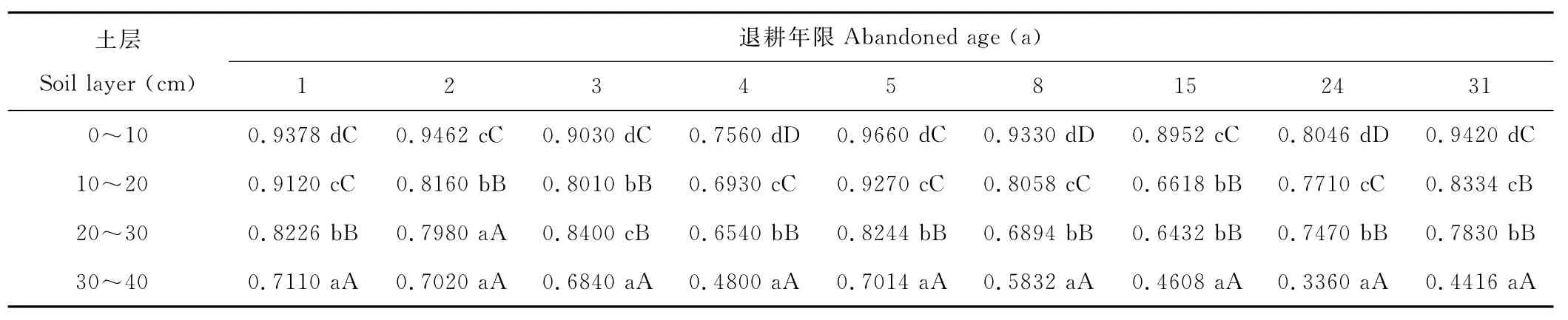
表2 不同土层不同退耕年限土壤过氧化氢酶分布Table 2 The distribution of soil catalase in different soil layer and abandoned age mL/(g·20min)
不同年限退耕区次生草地相同土层间过氧化氢酶呈波动式变化(表2)。退耕1~3a间变化不大,到4a时,显著减小,退耕第5a时,突然增加,达到最大值,然后缓慢减小,退耕24a后出现上升的现象。总体来看,过氧化氢酶随着退耕区次生草地自然恢复其活性变化不大,特别位于表层(0~10cm)相对稳定,除退耕4a土壤酶活性为0.7560mL/(g·20min)外,退耕1~31a土壤酶活性都在0.80~0.97mL/(g·20min)之间。
2.2.2 蔗糖酶 在0~40cm土层中,0~10cm土层蔗糖酶活性明显高于30~40cm土层(表3)。除退耕2a的20~30cm与30~40cm、退耕3a的10~20cm与20~30cm之间土壤蔗糖酶活性外,其余所有不同年限退耕区次生草地各土层间蔗糖酶活性差异均达到显著水平。不同年限退耕区次生草地土壤蔗糖酶活性分布随土层深度的增加而逐渐降低的趋势比较明显。
不同年代退耕区次生草地蔗糖酶活性总体表现出波动式增大,再波动式减小的趋势(表3)。当退耕4a时,蔗糖酶活性达到同一层的最大值(21.7955,20.8402,17.3809和10.0598mg/g·d),然后随着退耕区次生草地植被的自然恢复逐渐减小,当退耕时间达到15a时,表层蔗糖酶活性显著增强,10~20cm土层间的蔗糖酶活性相对平稳,但20~30cm及30~40cm土层间蔗糖酶的活性继续减小,分别降低到该层31a的最小值2.0557和1.6776mg/(g·d)。

表3 不同土层不同退耕年限土壤蔗糖酶的分布Table 3 The distribution of soil sucrase in different soil layer and abandoned age mg/(g·d)
2.2.3 脲酶 在0~40cm土层中,表层土壤脲酶活性明显高于下层(表4)。除退耕1和4a的土壤脲酶外,其余各土层间脲酶活性差异均达到显著水平(P<0.05),退耕4a的土壤脲酶各层间虽然也表现出从上到下逐渐降低的趋势,但各层间相差较小(0.0114,0.0322和0.0003mg/g·d);退耕24a的土壤脲酶各层间相差较大(0.2063,0.0849和0.0541mg/g·d),分别是退耕4a的18.1,2.6和180倍,均达到极显著水平(P<0.01)。不同年限退耕区次生草地土壤脲酶活性表现出上层大于下层的特点。这与尹伟等[27]对巴音布鲁克不同建植期人工草地土壤酶活性研究相一致。

表4 不同土层不同退耕年限土壤脲酶分布Table 4 The distribution of soil urease in different soil layer and abandoned age mg/(g·d)
由表4可以看出,脲酶活性呈现出随着退耕年限的增加而波动式减小趋势,最后趋于稳定。波动幅度比较平稳,退耕3a时,明显减小,退耕4a时,恢复到平稳状态,随着植被的恢复,退耕15a的时候相对平稳。
2.2.4 磷酸酶 磷酸酶活性(表5)在0~10cm土层中明显高于其他3层,不同年限退耕区次生草地磷酸酶分布均随土层深度的增加而逐渐降低,并且表层土壤磷酸酶活性在4层总酶活性中占有较大的比例(分别占39%,37%,32%,35%,38%,38%,30%,35%,30%),退耕1a到5a的土壤表层磷酸酶活性与以下各层间均表现出极显著的差异,退耕8a的土壤表层磷酸酶与10~20cm土层间表现出显著差异,退耕15a后,0~10cm与10~20 cm土层间磷酸酶活性差异不显著。
磷酸酶活性呈现出随着退耕年限的增加而波动式增大,再波动式减小趋势(表5)。其活性的波动趋势与蔗糖酶相似,在退耕1到3a间,变化相对平稳,到第4a的时候显著增大,达到了最大值3.6030mg/(g·d),随着退耕区次生草地植被的恢复,退耕8a时,逐渐减小,退耕15a,磷酸酶活性相对平稳,无论是不同年限还是不同土层深度,其酶的活性与最大值都相差2.0257~3.1897mg/(g·d)。

表5 不同土层不同退耕年限土壤磷酸酶分布Table 5 The distribution of soil phosphatase in different soil layer and abandoned age mg/(g·d)
3 讨论
石羊河流域下游退耕区次生草地植被在31a的自然恢复过程中,植物物种数量由退耕1a样方内的22种,减少为退耕31a样方内的5种;植被群落由一年生草本为建群种,多年生草本与个别灌木植物为伴生种,逐渐演替为多年生草本植物,最后演替为单一的灌木群落,趋于稳定。表明物种丰富度指数与多样性指数逐渐减小,均匀度指数增大,退耕区次生草地植被的演替具有递进性;在植被自然恢复的31a间,不同年限退耕区次生草地样方内均出现灌木植物苏枸杞,在退耕1~15a的样方内都出现了骆驼蒿、骆驼蓬及盐生草等草本植物,在退耕8~31a的样方内都出现了白刺(Nitrariasibirica)、盐爪爪、红砂等灌木植物,显示出次生草地植被在自然恢复过程中主要物种具有较强的连续性。总体来说,植被演替速度比较缓慢。
随着退耕区次生草地自然恢复,土壤过氧化物酶、蔗糖酶、脲酶和磷酸酶活性均表现出随土壤深度的增加而逐渐减小。同时也发现,0~10cm土层的酶活性在4层总酶活性中占有较大的比例。该结果与邰继承等[28]、文都日乐等[29]、秦燕等[30]、朱丽等[31]研究得出的土壤酶活性垂直变化的特点相一致。究其原因,由于石羊河下游土壤肥力较差,只有表层有少量的枯枝落叶和腐殖质,可以支持微生物的生长,表层温度条件和通气状况良好,一旦遇到降雨,微生物旺盛生长,代谢活跃,使表层的土壤酶活性提高。研究区干旱少雨,自然降雨只能贮藏于土壤表层,随着土壤剖面的加深,土壤水分显著减少,土壤温度的降低,限制了土壤微生物的正常活动及代谢产酶能力。由于这些因素的综合作用,使得土壤酶活性随着土层的加深而逐渐降低,而且,表层土壤酶活性所占比例较大。
随着退耕区次生草地植被演替年限的增加,土壤中酶活性总的趋势是逐渐减小,但各有不同。过氧化氢酶活性呈现出逐渐减小,再增大,最后趋于稳定,达到相对平衡状态。退耕初期由于耕作施肥刚刚停止,有机质和其他相关因子含量较高,1a生草本植物迅速生长,占据优势,土壤微生物生长旺盛,过氧化氢酶的活性较强;随着退耕年限的增加,土壤湿度减小,土壤中有机质和其他因子含量下降,限制了土壤微生物的正常活动,过氧化氢酶的活性也随之减小,退耕4a时,达到极小平均值(0.6458mL/g·20min);退耕5a后,植被逐渐由草本植物演替为灌木植物,一年生或多年生草本植物死亡根系逐步增多,经过腐烂,增加土壤有机物质,有利于土壤微生物的代谢产酶能力的提高,促进了过氧化氢酶的活性,出现各土壤层的平均峰值(0.8547mL/g·20min),并趋于稳定,达到平衡状态。
蔗糖酶活性总体表现出波动式增大,再波动式减小,最后趋于相对稳定,达到相对平衡状态。当退耕4a时,蔗糖酶活性达到同一层的最大值,退耕5a时,又基本恢复到退耕3a时的状况,然后随着退耕区次生草地植被的自然演替,逐渐趋于相对稳定,特别是退耕15,24和31a的10~20cm土层间的蔗糖酶活性十分相近。蔗糖酶在土壤易溶性营养物质中起着非常重要的作用,其活性可以作为评价土壤熟化程度和土壤肥力水平的一个指标[14],说明退耕4a时,土壤的熟化程度和土壤肥力水平达到了最好状态,到退耕5a时,又恢复到退耕3a时的状况,可能与退耕最后一年种植的不同农作物有关,退耕4a的土地,最后一年种植豆科的紫花苜蓿(Medicago sativa),其余土地种植的农作物是西瓜(Citrulluslanatus)、茴香(Foeniculumvulgare)、棉花(Gossypiumspp.)等,这一现象的出现,正好与种植豆科植物可以改良土壤相吻合,说明退耕区次生草地蔗糖酶的活性与退耕最后一年种植的农作物有密切的关系。
脲酶活性呈现出随着退耕区次生草地植被恢复年限的增加而波动式减小,最后趋于稳定。波动幅度比较平稳,退耕3a时,明显减小,脲酶活性平均值由退耕1a时的0.5487mg/(g·d)下降到3a时的0.3561mg/(g·d),退耕4a时,恢复到平稳状态,随着植被的恢复,最后趋于相对平稳。这主要是退耕3a后,群落内一年生草本植物基本退出,多年生草本的种类显著增加,群落内物种总数量由退耕1a的22种减少为10种,苏枸杞、盐生草、骆驼蒿、骆驼蓬、顶羽菊等成为群落优势种,这说明草本植物的退出可能使脲酶活性降低。退耕4a时,草本植物死亡根系经过腐烂,增加了土壤有机物质,促进土壤微生物的产酶能力,提高了脲酶的活性,随后逐渐下降,达到平衡状态。
磷酸酶活性呈现出波动式增大,再波动式减小趋势。其变化趋势与蔗糖酶很相似,在退耕1到3a间,变化相对平稳,到第4a的时候显著增大,达到了最大值,随后磷酸酶活性逐渐趋于平稳。土壤磷酸酶可以加速有机磷的脱磷速度,是生物磷代谢的重要酶类,是反应土壤磷素水平的重要指标[32]。说明退耕4a时,土壤磷素水平达到最大值,这一现象的出现,有可能也与退耕最后一年种植的不同农作物有关;另外也可能与退耕4a出现的甘肃柽柳(Tamarixgansuensis)、沙生针茅(Stipaglareosa)、铁线莲(Clematisflorida)等根系分泌物有关。退耕区次生草地土壤磷酸酶的活性与退耕最后一年种植的农作物及柽柳、沙生针茅、铁钱莲等根系分泌物的相关性还需要进一步的研究。
在退耕区次生草地恢复过程中,研究参试酶的活性,虽然变化规律不同,但总的趋势与物种丰富度和多样性指数变化规律相一致,随着退耕区次生草地恢复年限的增加,呈波动式下降。退耕初期物种丰富度和多样性指数最大,土壤酶的活性也最大,随着退耕区次生草地植被恢复时间的延长,均匀度指数逐渐增大,但丰富度和多样性指数减小,土壤酶活性降低。因而,土壤酶活性与植被的恢复是一个互动过程,它们之间存在着互相回馈的响应。这就表明,土壤酶的活性在退耕区次生草地植被恢复过程中,随着植被的物种丰富度与多样性指数的增大而增加。这是由于,退耕1a时,土壤中保持良好的水分和养分条件,促进田间杂草生长,物种繁多,积累了较多的枯枝落叶,腐烂后增加了土壤中的腐殖质和有机质含量,使微生物生长代谢活跃,产酶能力增加,提高了土壤酶的活性。退耕8~15a之后,因民勤石羊河下游的特殊自然条件,风沙大、干旱少雨,年蒸发量大于降雨量,只有极少数耐干旱的物种生存下来,退耕24~31a后,耐干旱性较差的植物也逐步被淘汰,植物生长量变小,产生的枯枝落叶与根部分泌物减少,有机质随之下降,土壤微生物的活动减弱,数量减少,土壤酶活性也随之下降。由于这些因素的综合作用,使得土壤酶活性随着退耕区次生草地植被自然恢复而逐渐降低。
在退耕区次生草地的恢复过程中,土壤酶活性和植被自然恢复都是极其复杂而又缓慢的过程,其酶活性不但与植被自然演替过程中的物种丰富度、物种多样性及退耕最后一年种植的农作物有关,还与土壤的有机肥、微团聚体[33]、温度[34]等其他理化性状有关系。鉴于土壤酶对各种相关因子的敏感性,还需进一步加强相关方面的研究。
[1] 农业大词典编辑委员会.农业大词典[M].北京:中国农业出版社,1998.
[2] 周礼恺.土壤酶学[M].北京:科学出版社,1987.
[3] 焦婷,常根柱,周学辉,等.高寒草甸草场不同载畜量下土壤酶与土壤肥力的关系研究[J].草业学报,2009,18(6):98-104.
[4] 任祖淦,陈玉水,唐福钦,等.有机无机肥料配施对土壤微生物和酶活性的影响[J].植物营养与肥料学报,1996,2(3):279-283.
[5] 蒋和,翁文钰,林增泉.施肥十年后的水稻土微生物学特性和酶活性的研究[J].土壤通报,1990,21(6):265-268.
[6] 袁玲,杨邦俊,郑兰君,等.长期施肥对土壤酶活性和氮磷养分的影响[J].植物营养与肥料学报,1997,3(4):300-306.
[7] 张为政.作物茬口对土壤酶活性和微生物的影响[J].土壤肥料,1993,(5):12-14.
[8] 程国华,郭树凡,薛景珍,等.长期施用含氯化肥对土壤酶活性的影响[J].沈阳农业大学学报,1994,25(4):360-365.
[9] Paul E A,Clack F E.Soil Microbiology and Biochemistry[M].New York:Academic Press,1998:104-109.
[10] 马晓飞,李艳红,杨爱霞,等.新疆甘家湖湿地边缘带土壤酶活性研究[J].干旱区研究,2012,29(3):405-412.
[11] 张咏梅,周国逸,吴宁.土壤酶学的研究进展[J].热带亚热带植物学报,2004,12(1):83-90.
[12] 邱东,程争鸣,张元明,等.寡营养细菌对古尔班通古特沙漠土壤环境的影响[J].干旱区研究,2012,29(1):148-154.
[13] 王俊华,尹睿,张华勇,等.长期定位施肥对农田土壤酶活性及其相关因素的影响[J].生态环境,2007,16(1):191-196.
[14] 关松荫.土壤酶及其研究法[M].北京:农业出版社,1986.
[15] 杨成德,龙瑞军,陈秀蓉,等.东祁连山高寒灌丛草地土壤微生物量及土壤酶季节性动态特征[J].草业学报,2011,20(6):135-142.
[16] 冯绳武.民勤绿洲的水系演变[J].地理学报,1963,29(3):241-249.
[17] 苏志珠,卢琦,吴波,等.气候变化和人类活动对我国荒漠化的可能影响[J].中国沙漠,2006,26(3):229-334.
[18] 王世金,何元庆,赵成章.西北内陆河流域水资源优化配置与可持续利用[J].水土保持研究,2008,15(5):22-29.
[19] 李丁,马金珠,南忠仁.民勤盆地地下水位下降特征与可持续利用评价[J].中国沙漠,2004,21(6):734-739.
[20] 王琪,史基安,张中宁,等.石羊河流域环境现状及其演化趋势分析[J].中国沙漠,2003,23(1):46-52.
[21] 刘恒,钟华平,顾颖.西北干旱内陆河区水资源利用与绿洲演变规律研究[J].水科学进展,2001,12(3):378-384.
[22] 姚槐应,黄昌勇.土壤微生物生态学及其实验技术[M].北京:科学出版社,2007.
[23] 杨自辉.民勤沙井子地区40a来荒漠植被变迁初探[J].中国沙漠,1999,19(4):395-397.
[24] 彭鸿嘉,傅伯杰,陈利顶,等.甘肃民勤荒漠区植被演替特征及驱动力研究——以民勤为例[J].中国沙漠,2004,24(5):628-633.
[25] 何芳兰,李治元,赵明,等.民勤绿洲盐碱化退耕地植被自然演替及土壤水分垂直变化研究[J].中国沙漠,2010,30(6):1374-1380.
[26] 党荣理,潘晓玲.西北干旱荒漠区种子植物科的区系分析[J].西北植物学报,2002,22(1):24-32.
[27] 尹伟,胡玉昆,柳妍妍,等.巴音布鲁克不同建植期人工草地土壤生物学特性研究[J].草业学报,2010,19(5):218-226.
[28] 邰继承,杨恒山,张庆国,等.不同生长年限紫花苜蓿人工草地土壤酶活性及分布[J].草业科学,2008,25(4):76-78.
[29] 文都日乐,李刚,张静妮,等.呼伦贝尔不同草地类型土壤微生物量及土壤酶活性研究[J].草业学报,2010,19(5):94-102.
[30] 秦燕,牛得草,康健,等.贺兰山西坡不同类型草地土壤酶活性特征[J].干旱区研究,2012,29(5):870-877.
[31] 朱丽,郭继勋,鲁萍,等.松嫩羊草草甸羊草、碱茅群落土壤酶活性比较研究[J].草业学报,2002,11(4):28-34.
[32] 薛立,邝立刚,陈红跃,等.不同林分土壤养分、微生物与酶活性的研究[J].应用生态学报,2003,40(2):280-285.
[33] 林天,何园球,李成亮,等.红壤旱地中土壤酶对长期施肥的响应[J].土壤学报,2005,42(4):682-686.
[34] 秦纪洪,张文宣,王琴,等.亚高山森林土壤酶活性的温度敏感性特征[J].土壤学报,2013,50(6):174-178.
