花生生理和农艺性状对冠层温度的影响
2014-03-26任学敏王长发王小立
任学敏,朱 雅,王长发,王小立
(1 南阳师范学院 a生命科学与技术学院,b 图书馆,河南 南阳 473061;2 西北农林科技大学 农学院,陕西 杨凌 712100)
近年来,冠层温度已成为大田作物研究的热点问题之一,其在农田水肥管理[1-2]、耐热基因型作物筛选[3]、抗旱基因型作物筛选[4]等方面得到了深入的研究和广泛应用,对于农业生产实践的指导显示出了强大的优越性。冠层温度能够用于指导农业生产实践是因为:(1)在不同环境条件下(高温、干旱、肥力等),同一基因型作物品种的冠层温度表现不同;(2)在相同环境条件下,同种作物的不同基因型冠层温度存在差异。有关同种作物不同基因型冠层温度的分异,已在多种作物上发现,如小麦[5]、水稻[6]、棉花[7]、花生[8]、大豆[9]、谷子[10]和豌豆[11]等。同时还发现,冠层温度偏低的品种在生理特性、产量、品质等方面比冠层温度偏高的品种有更优良的表现[12]。作物冠层温度的差异主要受两大方面因素影响,即外因和内因以及二者的相互作用[12]。对于外因,目前已有大量研究,主要是外部环境条件的影响,如第一热源(太阳辐射)、第二热源(地面辐射)、CO2浓度、肥料[2,12-14]等;对于内因,一些学者认为是作物自身生物学特性的影响,在这方面研究较多的是蒸腾作用[9,15],而对于其他生物学性状(如生理和农艺性状)鲜有研究。本试验以12种北方大花生为研究材料,探索了一些生理和农艺性状对冠层温度的影响,以期为揭示不同基因型花生品种冠层温度差异机理提供理论依据。
1 材料与方法
1.1 供试材料
供试花生品种12个,均为大花生,其中开农41、豫花2号、豫花153、豫花7号、豫花9331、标花2号、濮花9506、濮科花2号、豫花9327和豫花11号引自河南,鲁花11号和鲁花9号引自山东。
1.2 试验设计
试验地位于陕西关中平原中部的渭河谷地头道塬,属北方大花生区。该区域为暖温带半湿润气候,年均降水量约700 mm,90%以上集中在4-9月。试验于2007年进行,采用随机区组排列,4次重复,试验地前茬空茬,采用起垄地膜覆盖种植法,垄距0.8 m,垄高0.1 m,垄面宽0.55 m,畦沟宽0.26 m。04-24人工开沟带尺点播。每小区3垄6行,行长2.4 m,行距 0.35 m,株距0.16 m,每穴播种2粒,播深3~5 cm。播种前以尿素200 kg/hm2、二胺300 kg/hm2的标准施底肥。田间管理参照北方大花生生产区的管理方法进行。
1.3 冠层温度的测定
采用BAU-Ⅰ型红外测温仪对冠层温度进行测定。观测时间为各材料冠层温度差异最明显的午后(13:00-15:00)。自花生结荚开始,每隔7~9 d观测1次。选择各花生品种群体生长均匀一致且有代表性的部位作为测点, 避免红外线照射裸露地面。观测时, 感应头距花生冠层约20 cm, 探棒倾角约30°。对每个品种的4次重复进行往返观测,其平均值作为该次的冠层温度值。
1.4 测定指标及方法
1.4.1 生理指标的测定 在花生冠层温度测定的当天,对每个品种的4个重复分别取样,取样部位为花生功能叶片(主茎顶3叶)[8]。所取样品放入冰壶带回实验室,用于测定过氧化氢酶(Catalase,CAT)、超氧化物歧化酶(Superoxide dismutase,SOD)、硝酸还原酶(Nitrate reductase,NR)活性及丙二醛(Malondialdehyde,MDA)、可溶性蛋白、可溶性糖和叶绿素含量。CAT活性用紫外吸收法测定[16],SOD活性参照王爱国等[17]的方法测定,NR活性参照文献[16]的方法测定,MDA含量用硫代巴比妥酸法[18]测定,可溶性蛋白含量用考马斯(G-250)亮蓝法[19]测定,可溶性糖含量用蒽酮法[19]测定,叶绿素含量采用体积分数80%丙酮低温浸提法[20]测定。
1.4.2 农艺性状调查方法 花生收获时每小区随机选取3株,4 次重复共12株,分别调查分枝数,并测量侧枝长、主茎高,结果取平均值。
1.5 数据处理
对花生生理和农艺性状与冠层温度的关系,均分别采用相关分析和通径分析方法进行分析。所有分析均用SAS8.0统计软件[21]完成。图1用Origin7.5完成。
2 结果与分析
2.1 不同基因型花生品种冠层温度的差异
图1显示了不同基因型花生品种冠层温度与所有品种冠层温度平均值的差值。从图1可以看出,豫花153冠层温度最低,比平均冠层温度低1.08 ℃;开农41冠层温度最高,高于平均冠层温度0.84 ℃;其他10个花生品种冠层温度由高到低依次为标花2号、濮花9506、濮科花2号、豫花7号、豫花11号、鲁花11号、鲁花9号、豫花2号、豫花9327和豫花9331,其中豫花2号、鲁花9号、鲁花11号和豫花11号与平均冠层温度差异不明显。

图1 不同基因型花生品种冠层温度与平均冠层温度的差值
2.2 花生冠层温度与生理和农艺性状的相关分析
花生冠层温度与各生理指标以及各生理指标之间的相关系数如表1所示。由表1可知,花生冠层温度与CAT活性、可溶性蛋白含量、叶绿素含量、NR活性和SOD活性存在负相关关系,其中与前三者相关达极显著水平(P≤0.01),相关系数r分别为-0.400,-0.378和-0.515;与NR活性相关达显著水平(P≤0.05),相关系数r为-0.282,与SOD活性相关不显著(P>0.05)。冠层温度与MDA和可溶性糖含量呈正相关关系,其中与前者的相关达显著水平(P≤0.05),相关系数r为0.243;与可溶性糖含量的相关不显著(P>0.05)。从这些相关系数可知,叶绿素含量、CAT活性、可溶性蛋白含量、NR活性和MDA含量是影响花生冠层温度的主要生理指标,且其影响强度依次减小。
花生冠层温度与分枝数、侧枝长和主茎高均呈负相关(表2),相关系数r分别为-0.604,-0.290和-0.661,其中与分枝数和主茎高的相关性达显著水平。3个指标对冠层温度的影响强度依次为:主茎高>分枝数>侧枝长。
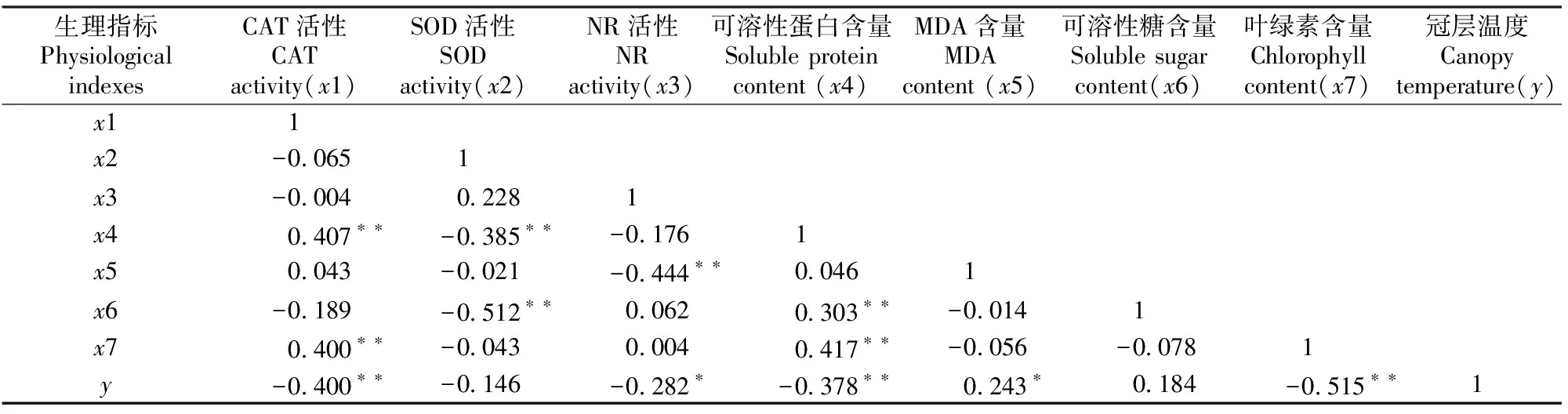
表1 花生生理指标与冠层温度的相关系数(n=72)
2.3 花生冠层温度与生理和农艺性状的通径分析
花生的一些生理指标之间以及农艺性状指标之间存在显著的相关关系(表1,表2),这可能使得一些指标对冠层温度的作用通过另一些指标来实现。因此,简单相关分析中生理和农艺性状指标与冠层温度的相关系数仅仅反映了各指标对冠层温度的总影响力。为了确定各指标对冠层温度的直接影响力和间接影响力,进而确定各指标的相对重要性,有必要采用通径分析对简单相关分析中的相关系数进行剖析。花生生理指标与冠层温度的通径分析结果如表3所示。由表3可以看出,根据直接通径系数的绝对值,各指标对冠层温度的直接影响力大小为:叶绿素含量>SOD活性>NR活性>可溶性蛋白含量>CAT活性>MDA含量>可溶性糖含量。
由间接通径系数(表3)可知,花生各生理指标通过其他指标不同程度地影响冠层温度。CAT活性主要通过可溶性蛋白和叶绿素含量对冠层温度起作用,通过二者所起的作用(通径系数之和为-0.239)超过了其本身;SOD活性对冠层温度的影响较大,仅次于叶绿素含量,但其通过NR活性和可溶性蛋白含量对冠层温度的影响与其自身对冠层温度的影响方向相反,相互抵消后使得SOD活性的总影响力大大降低;NR活性主要通过自身影响冠层温度(直接通径系数绝对值较大),但也通过SOD活性、可溶性蛋白和MDA含量对冠层温度起一定的间接作用;可溶性蛋白含量通过间接作用影响冠层温度的指标主要有CAT活性、SOD活性、NR活性和叶绿素含量,但总的间接作用(-0.143)小于可溶性蛋白的直接作用(-0.250),说明可溶性蛋白含量也是主要通过自身影响冠层温度;MDA含量对冠层温度的直接影响较小,与其通过NR活性对冠层温度的间接影响相近,而通过其他指标的间接影响均很小;可溶性糖含量对冠层温度的直接通径系数是0.026,比其通过CAT活性、SOD活性、可溶性蛋白含量和叶绿素含量的间接通径系数均小,说明可溶性糖含量对冠层温度的作用不大;叶绿素含量对冠层温度的直接通径系数是-0.343,是所有生理指标中最大的,其通过CAT活性和可溶性蛋白含量对冠层温度也有较大的间接作用。

表2 花生农艺性状与冠层温度的相关系数(n=12)
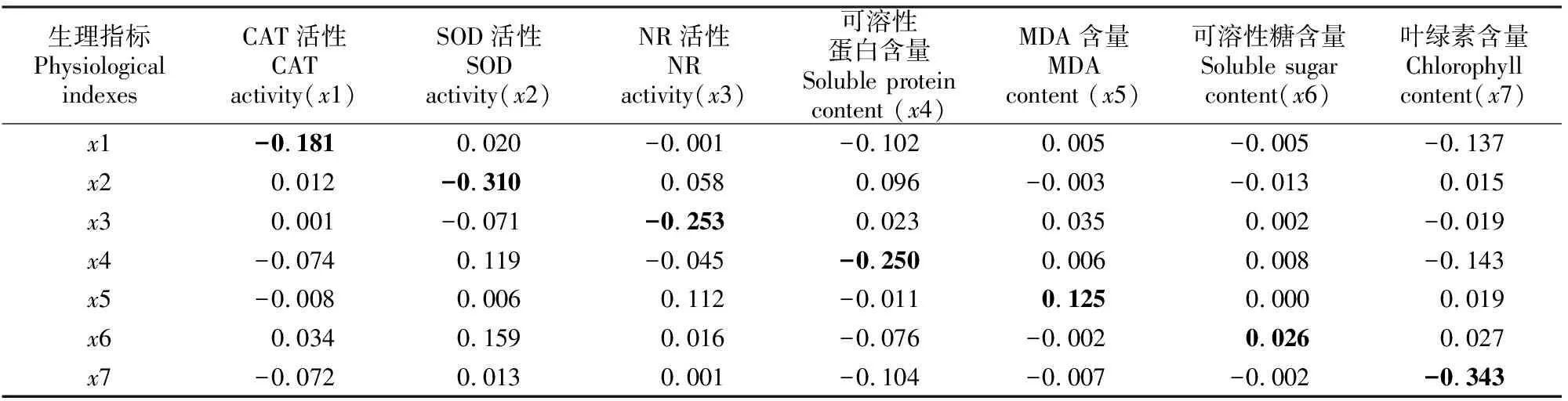
表3 花生生理指标与冠层温度的通径分析
花生农艺性状与冠层温度的通径分析结果(表4)显示,主茎高直接通径系数的绝对值最大,因此是影响花生冠层温度的最主要的农艺性状,其次是侧枝长,分枝数的影响最小。

表4 花生农艺性状与冠层温度的通径分析
分枝数对冠层温度的作用主要是直接作用,但由于其与主茎高存在正相关(r=0.227),通过主茎高还可使分枝数对冠层温度的作用有较高提升;侧枝长对冠层温度的直接通径系数是0.579,而其通过主茎高的间接通径系数是-0.835,说明侧枝长是通过主茎高间接作用于冠层温度的;主茎高对冠层温度的直接作用最大,作用方向为负向,其通过侧枝长对冠层温度的作用为正向,相互抵消后主茎高的总效应依然较大(-0.566),说明主茎高确实是影响冠层温度的主要因素。
3 讨 论
张嵩午和王长发[22]对小麦冠层温度的研究认为,内因和外因共同决定了冠层温度。在较小尺度下,植株形态和生理特性等内因的差异对小麦冠层温度具有决定意义,外因主要是第二热源对冠层温度起作用[22]。生理特性决定着植物蒸腾作用的强弱,而蒸腾作用对冠层温度有重要影响。由农田热量平衡方程式[23]可知,净辐射(R)用于蒸腾作用的能量多,则必然使得用于加热株茎的能量(QT)和叶片的能量(QA)减少。因此,蒸腾作用强的品种其冠层温度必然较低,反之则较高[13]。

第二热源是影响作物冠层温度的最主要的外在因素,而第二热源对冠层的加热作用与作物高度密切相关。张嵩午[13]在对小麦的研究中发现,第二热源对冠层的加热作用由植株基部向上逐渐减弱,因此,植株越高第二热源的加热作用越小,冠层温度相应越低。本研究虽然没有直接测定第二热源的加热作用,但仍发现花生主茎高与冠层温度存在显著的负相关(P≤0.05),且相关系数的绝对值最大;进一步的通径分析结果表明,主茎高对花生冠层温度的直接作用最大,说明主茎高确实是影响冠层温度的主要因素,其影响机理可能涉及到其对第二热源的影响。本研究还发现,分枝数和侧枝长对冠层温度也有一定的直接影响,但二者通过主茎高对冠层温度的间接作用均较大,尤其是侧枝长,通过主茎高的间接作用大于直接作用,并使得总作用的方向与直接作用相反,这进一步说明了主茎高对花生冠层温度影响的重要性。
影响作物冠层温度的因素包括外在和内在多个方面以及各方面的相互作用。到目前为止,即便是研究最为广泛和深入的小麦,有关其冠层温度的影响因素尤其是内因依然没有完全清楚。本研究仅以花生1种作物为试验材料,探索其生理和农艺性状对冠层温度的影响,所得结果不可避免地具有一定的局限性和片面性,若要全面揭示作物冠层温度的形成机理,尤其是内在因素的作用,还需要进行更广泛(如拓展到更多作物)、更深入(如从分子水平上探索决定冠层温度的基因或系列基因)地研究。
[参考文献]
[1] Durigon A,de Jong van Lier Q.Canopy temperature versus soil water pressure head for the prediction of crop water stress [J].Agricultural Water Management,2013,127:1-6.
[2] 周春菊,张嵩午,王林权,等.施肥对小麦冠层温度的影响及其与生物学性状的关联 [J].生态学报,2005,25(1):18-22.
Zhou C J,Zhang S W,Wang L Q,et al.Effect of fertilization on the canopy temperature of winter wheat and its relationship with biological characteristics [J].Acta Ecologica Sinica,2005,25(1):18-22.(in Chinese)
[3] Kumari M,Pudake R N,Singh V P,et al.Association of staygreen trait with canopy temperature depression and yield traits under terminal heat stress in wheat (TriticumaestivumL.) [J].Euphytica,2013,191(1):87-97.
[4] Guendouz A,Guessoum S,Maamri K,et al.Canopy temperature efficiency as indicators for drought tolerance in durum wheat (TriticumdurumDesf.) in semi-arid conditions [J].Journal of Agriculture and Sustainability,2012,1(1):23-38.
[5] 张嵩午.小麦温型现象研究 [J].应用生态学报,1997,8(5):471-474.
Zhang S W.Temperature type phenomenon of wheat [J].Chinese Journal of Applied Ecology,1997,8 (5):471-474.(in Chinese)
[6] 高继平,韩亚东,王晓通,等.水稻齐穗期冠层温度分异及其相关特性的研究 [J].沈阳农业大学学报,2011,42(4):399-405.
Gao J P,Han Y D,Wang X T,et al.Canopy temperature difference and the related characteristics at heading stage in rice [J].Journal of Shenyang Agricultural University,2011,42(4):399-405.(in Chinese)
[7] 韩 磊,王长发,王 建,等.棉花冠层温度分异现象及其生理特性的研究 [J].西北农业学报,2007,16(3):85-88.
Han L,Wang C F,Wang J,et al.The study on differences of the canopy temperature and physiological characteristics of cotton [J].Acta Agriculturae Boreali-Occidentalis Sinica,2007,16(3):85-88.(in Chinese)
[8] 任学敏,王长发,秦晓威,等.花生群体冠层温度分异现象及其生理特性研究初报 [J].西北农林科技大学学报:自然科学版,2008,36(6):68-72.
Ren X M,Wang C F,Qin X W,et al.Preliminary study on canopy temperature difference of peanut varieties population and physiological characteristics [J].Journal of Northwest A&F University:Nat Sci Ed,2008,36(6):68-72.(in Chinese)
[9] 李永平,王长发,赵 丽,等.不同基因型大豆冠层冷温现象的研究 [J].西北农林科技大学学报:自然科学版,2007,35(11):80-83,89.
Li Y P,Wang C F,Zhao L,et al.Low temperature phenomena of soybean of different genotype [J].Journal of Northwest A&F University:Nat Sci Ed,2007,35(11):80-83,89.(in Chinese)
[10] 秦晓威,王长发,任学敏,等.谷子冠层温度分异现象及其生理特性研究 [J].西北农业学报,2008,17(2):101-105.
Qin X W,Wang C F,Ren X M,et al.Study on canopy temperature and physiological characteristics of different millet genotypes [J].Acta Agriculturae Boreali-Occidentalis Sinica,2008,17(2):101-105.(in Chinese)
[11] 王 一,王长发,邹 燕,等.豌豆冠层温度分异现象及其生理特性 [J].西北农业学报,2009,18(4):133-136.
Wang Y,Wang C F,Zou Y,et al.Study on canopy temperature and physiological characteristics of different peas [J].Acta Agriculturae Boreali-Occidentalis Sinica,2009,18(4):133-136.(in Chinese)
[12] 张嵩午.冷型小麦的概念特性未来 [J].中国科学基金,2006(4):210-214.
Zhang S W.Cold wheat’s concept,characteristic and future [J].Bulletin of National Natural Science Foundation of China,2006(4):210-214.(in Chinese)
[13] 张嵩午.小麦群体的第二热源及其增温效应 [J].生态学杂志,1990,9(2):1-6.
Zhang S W.The second heat source of wheat populations and it’s heating effects [J].Journal of Ecology,1990,9(2):1-6.(in Chinese)
[14] Kimball B A.Lessons from FACE:CO2effects and interactions with water,nitrogen and temperature [M]//Kimball B A.Handbook of climate change and agroecosystems:Impacts,adaptation,and mitigation.London:Imperial College Press,2011:87-107.
[15] Monteith J L,Szeicz G.Radiative temperature in the heat balance of nature surface [J].Quarterly Journal of the Royal Meteorological Society,1962,88(378):496-507.
[16] 孙 群,胡景江.植物生理学研究技术 [M].陕西杨凌:西北农林科技大学出版社,2006.
Sun Q,Hu J J.Research technology of plant physiology [M].Yangling,Shaanxi:Northwest A&F University Press,2006.(in Chinese)
[17] 王爱国,罗广华,邵从本,等.大豆种子超氧物歧化酶的研究 [J].植物生理学报,1983,9(1):77-83.
Wang A G,Luo G H,Shao C B,et al.A study on the superoxide dismutase of soybean seeds [J].Acta Phytophysiologia Sinica,1983,9(1):77-83.(in Chinese)
[18] Heath R L,Packer L.Photoperoxidation in isolated chloropla-sts:Ⅰ.Kinetics and stoichiometry of fatty acid peroxidation [J].Archives of Biochemistry and Biophysics,1968,125(1):189-198.
[19] 汪沛洪,陈毓荃,马仁懿,等.基础生物化学实验指导 [M].西安:陕西科学技术出版社,1986.
Wang P H,Chen Y Q,Ma R Y,et al.Experimental directions of fundamental biochemistry [M].Xi’an:Shaanxi Science and Technology Press,1986.(in Chinese)
[20] 高俊凤.植物生理学试验技术 [M].西安:世界图书出版社,2000.
Gao J F.Plant physiology testing techniques [M].Xi’an:World Book Press,2000.(in Chinese)
[21] 胡小平,王长发.SAS 基础及统计实例教程 [M].西安:西安地图出版社,2001.
Hu X P,Wang C F.Course of SAS foundation and statistical examples [M].Xi’an:Xi’an Map Press,2001.(in Chinese)
[22] 张嵩午,王长发.小麦低温基因型的研究现状和未来发展 [J].中国农业科学,2008,41(9):2573-2580.
Zhang S W,Wang C F.Research progress and tendency in wheat low temperature genotypes [J].Scientia Agricultura Sinica,2008,41(9):2573-2580.(in Chinese)
[23] 翁笃鸣,陈万隆,沈觉成,等.小气候和农田小气候 [M].北京:农业出版社,1981:69-74.
Weng D M,Chen W L,Shen J C,et al.Microclimate and farmland climate [M].Beijing:China Agricultural Press,1981:69-74.(in Chinese)
[24] Patterson T G,Moss D N,Brun W A.Enzymatic changes during the senescence of field-grown wheat [J].Crop Science,1980,20(1):15-18.
