rhKD/APPvar毕赤酵母分泌表达质粒的构建及其重组蛋白的表达和纯化
2014-03-23王心童王虹蛟孟威宏颜炜群任立群
王心童,王虹蛟,王 强,孟威宏,颜炜群,任立群
(1.吉林大学中日联谊医院神经内科,吉林 长春 130033;2.解放军第461医院内科,吉林 长春 130021;3.沈阳军区总医院心血管内科,辽宁 沈阳 110015;4.吉林大学再生医学科学研究所再生医学系,吉林 长春 130021)
牛胰蛋白酶抑制剂(bovine pancreatic trypsin inhibitor,BPTI)是Kunitz家族的丝氨酸蛋白酶抑制剂,已用于急、慢性肝损伤的研究[1-6]。但BPTI是1种从牛肺中提纯的碱性蛋白酶抑制剂,为异体蛋白,具有抗原性,长期反复应用能够产生特异性抗体,引起机体免疫反应。KD/APP是人Kunitz型蛋白酶抑制剂(Kunitz protease inhibitor,KPI),具有抑制丝氨酸蛋白酶的活性,是较为理想的BPTI替代品。虽然KD/APP与BPTI在结构、生物活性等方面有很多相似之处,但由于KD/APP活性中心的氨基酸残基是Arg15-Ala16-Met17(RAM),BPTI是Lys15-Ala16-Arg17(KAR),两者在酶的专一性、亲和常数和抑制酶谱等方面存在较大的差异。本研究利用KD/APP为基本骨架,用BPTI活性中心氨基酸残基KAR替代KD/APP活性中心的RAM,构建新的重组KD/APP变异体(rhKD/APPvar),并构建rhKD/APPvar真核表达质粒,实现其在毕赤酵母中的分泌表达,建立大规模发酵、纯化rhKD/APPvar的工艺,为筛选具有与BPTI在生物活性(抑制酶谱)和药效学类似的新的抑制剂提供物质基础。目前国内外尚未见相关报道。
1 材料与方法
1.1 菌株和主要试剂 表达质粒pPICZα和酵母菌株X-33购自美国Invitrogen公司;感受态菌XL1-Blue购自北京鼎国生物公司;LA Taq酶、T质粒、限制性内切酶和T4DNA连接酶购自日本Takara公司;感受态菌XL1-Blue和DNA快速纯化/回收试剂盒购自北京鼎国生物公司;质粒提取试剂盒购自美国Promega公司;SP Sepharose XL阳离子交换树脂购自瑞典Pharmacia公司;rhKD/APP-pPICZα重组质粒由吉林大学再生医学科学研究所提供。
1.2 构建rhKD/APPvar-pPICZα表达质粒 利用已构建好的rhKD/APP-pPICZα表达质粒,在KD/APP活性中心两侧设计2个酶切位点(ApaⅠ和SacⅡ),将KD/APP的活性中心RAM替换成BPTI的活性中心KAR。①在KD/APP活性中心5′端引入ApaⅠ位点:根据在KD/APP序列,设计引物扩增5′端引入ApaⅠ限制性内切酶位点。上游引物:5′-GGTCTGCAGTGAACAAGC-CGAGACTGGGCCCTG-3′(下划线部分为酶切位点);下游引物:5′-GAAGTCTAGATTAAATGGCGCTGCCACACA-3′。以KD/APP-pPICZα重组质粒为模板,行PCR反应,PCR反应程序:94℃变性4min;94℃变性30s,55℃退火30s,72℃延伸1min,共30个循环;72℃再延伸5min。回收PCR产物约200bp片段,经PstⅠ和XbalⅠ双酶切后,连接经PstⅠ和XbalⅠ双酶切的pPICZ质粒。②在KD/APP活性中心3′端引入SacⅡ位点:上游引物,5′-ATGGGGCCCTG-TAGAGCAATGATCCCGCGGTGG-3′(下划线部分为酶切位点);下游引物,5′-GAAGTCTAGATTAAATGGCGCTGCCACACA-3′。以①中获得的重组质粒为模板,PCR反应程序:94℃变性4min;94℃变性30s,55℃退火30s,72℃延伸1min,30个循环;72℃再延伸5min。回收PCR产物200bp片段,经PstⅠ和XbalⅠ双酶切后,连接同样经PstⅠ和XbalⅠ双酶切的pPICZ质粒,构建重组质粒Kas。用SacⅠ和SacⅡ双酶切鉴定,经酶切鉴定正确的重组质粒Kas经ApaⅠ和SacⅡ双酶切,去除KD/APP活性中心的RAM。③活性中心的替换:合成编码BPTI活性中心KAR的短链DNA,序列为5′-CTGTAAAGCTAGAATCTCGC-3′和5′-GAGATTCTAGCTTTACAGGGCC-3′。引物退火后获得的黏性末端与ApaⅠ和SacⅡ双酶切后的黏性末端连接。利用此特性将重组质粒Kas中的活性中心RAM与BPTI活性中心KAR的DNA进行替换,获得rhKD/APPvar-pPICZα重组表达质粒。经酶切鉴定正确的重组质粒转化大肠杆菌感受态XL1-Blue,大量扩增,并用质粒快速提取试剂盒提取质粒用于酶切鉴定及核酸序列分析。
1.3 rhKD/APPvar-pPICZαz转化毕氏酵母 将20μg rhKD/APPvar-pPICZα质粒用SacⅠ酶切,酚氯仿抽提,用10μL无菌水溶解。与80μL新鲜制备的X33感受态混匀,加入0.2cm电转杯,冰浴5min,以Bio-Rad Gene Pulser电转仪于1500V、25μF、200Ω条件下转化,同时加入1.0mL山梨醇,将菌液涂布于YPD(含100mg·L-1的Zeocin)的选择培养板上,30℃培养2~3d,筛选具有Zeocin抗性的生长良好的表达菌株。
1.4 酵母工程菌的诱导表达 取上述鉴定结果阳性的克隆接种于10mL BMGY(pH 6.0)培养基中,30℃震荡培养24h,至吸光度(A600)值达2.0~6.0收集细胞。等体积(10mL)BMMY(pH 6.0)重悬细胞沉淀,30℃震荡培养,诱导表达。在诱导过程中,每24h补充一次甲醇至终浓度为0.5%,同时补充灭菌超纯水,使发酵液总体积保持不变。在培养0、24、48、72、96和120h等时间点各取0.5mL发酵液,离心取上清用于聚丙烯酰胺凝胶电泳(SDS-PAGE)分析。
1.5 毕赤酵母表达rhKD/APPvar最佳pH值的确定 将生长培养基的菌体等量接入pH值分别为3.0、3.5、4.0、4.5、5.0、5.5和6.0的诱导培养基内进行培养。在培养0、24、48、72、96和120h等时间点各取0.5mL发酵液,离心取上清用于SDS-PAGE分析。
1.6 rhKD/APP的纯化 将rhKD/APPvar发酵液离心,取上清,用3倍体积水稀释并加入1/10体积的200mmol·L-1醋酸盐缓冲液(pH 4.0),用HCl调节其pH值为3.5。用10倍柱体积20mmol·L-1醋酸盐缓冲液(pH3.5)平衡SP Sepharose FF阳离子树脂。加样品rhKD/APPvar于平衡好的SP Sepharose FF阳离子树脂柱,用波长280nm监测。待穿透液有目的蛋白出现时,停止加样,用20mmol·L-1醋酸盐缓冲液冲洗树脂至A280值降至基线。用含1mol·L-1NaCl的20mmol·L-1醋酸盐缓冲液洗脱,并分步收集蛋白峰。用超滤纯化装置浓缩纯化后的rhKD/APPvar,并用水倍比稀释3~5次以除盐。应用SDS-PAGE分析确定纯化后蛋白的纯度和浓度,Bradford法精确定量。
2 结 果
2.1 重组质粒Kas的鉴定 利用已构建好的rhKD/APP-pPICZα表达质粒,在KD/APP活性中心两侧通过PCR引入2个酶切位点(ApaⅠ和SacⅡ),连接pPICZ质粒后,小提质粒进行酶切鉴定。用SacⅠ和SacⅡ双酶切重组质粒结果:重组质粒成功切下1000bp的片段,而对照组质粒未见此片段,证明2个酶切位点(ApaⅠ和SacⅡ)成功引入KD/APP活性中心两侧,且PCR获得的目的片段成功插入pPICZ质粒,构建了重组质粒Kas。见图1。
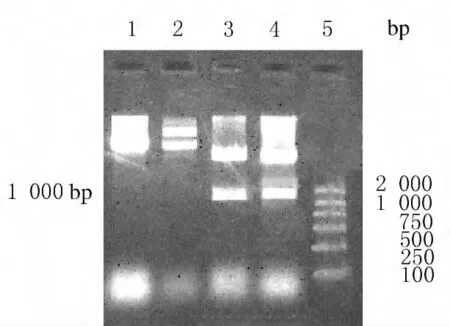
图1 SacⅠ和SacⅡ双酶切质粒产物琼脂糖凝胶电泳图Fig.1 Agarose gel electrophoregram of plasmid digested by SacⅠ和SacⅡ
2.2 活性中心替换的鉴定 合成编码BPTI活性中心KAR的短链DNA,退火后获得的黏性末端可与ApaⅠ和SacⅡ双酶切后的黏性末端连接。利用此特性将重组质粒Kas与BPTI活性中心KAR的DNA序列连接,获得rhKD/APPvar-pPICZα表达质粒。连接BPTI活性中心KAR后,SacⅡ位点消失,可以作为鉴定依据。经SacⅠ和SacⅡ双酶切,活性中心替换成功的KD/APPvar-pPICZα重组表达质粒无1000bp片段(图2)。
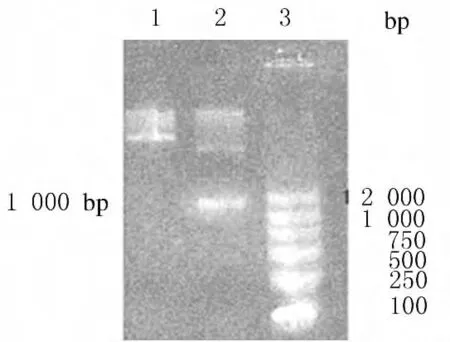
图2 活性中心替换鉴定的琼脂糖凝胶电泳图谱Fig.2 Agarose gel electrophoregram of substitute identification of active site
2.3 rhKD/APPvar-pPICZαz重组质粒测序rhKD/APPvar-pPICZαz重组质粒测序结果显示DNA序列与预期相符。见图3。

图3 替换后质粒DNA测序图谱Fig.3 Sequencing diagram of vector DNA after substituting
2.4 rhKD/APPvar蛋白表达 最适表达时间的优化:SDS-PAGE结果:甲醇诱导表达后在相对分子质量约6700处出现蛋白条带,诱导表达24h有蛋白分泌,随着诱导时间的延长其表达水平升高,120h即达到高峰。发酵液上清rhKD/APPvar表达水平可达285mg·L-1。见图4。毕赤酵母表达rhKD/APPvar的最佳pH值:毕赤酵母在pH 2.5~6.0培养基均能很好地生长,本实验在pH 6.0时发酵外源蛋白表达水平最高。见图5。

图4 rhKD/APPvar经时变化的SDS-PAGE图谱(考马斯亮蓝染色)Fig.4 SDS-PAGE graph of ongoing changes of rhKD/APPvar(Coomassie blue staining)
2.5 rhKD/APPvar的纯化 rhKD/APPvar发酵液离心取上清,经Sepharose SP FF阳离子交换层析,得到1个活性洗脱主峰,除盐浓缩后rhKD/APPvar在15%SDS-PAGE中显示1条考马斯亮蓝区带。纯化后纯度可达95%以上。见图6。
3 讨 论

图5 rhKD/APPvar SDS-PAGE图谱(考马斯亮蓝染色)Fig.5 SDS-PAGE graph of rhKD/APPvar(Coomassie blue staining)

图6 大规模纯化rhHKD/APPvar的SDS-PAGE图谱(考马斯亮蓝染色)Fig.6 SDS-PAGE graph of rhKD/APPvar with large-scale purification(Coomassie blue staining)
蛋白酶抑制剂在机体中参与凝血、纤溶、细胞迁移、细胞分化、凋亡和炎症反应等,在临床上主要应用于细胞凋亡、凝血、骨质疏松和肿瘤等疾病治疗[7]。目前丝氨酸蛋白酶抑制剂是研究最多、最为清楚的一类抑制剂,其中Kunitz家族的牛胰蛋白酶抑制剂(BPTI)的结构、功能及药理学方面的研究比较深入,已广泛应用于临床。BPTI是一种广谱的丝氨酸蛋白酶抑制剂,其相对分子质量为6500,是由58个氨基酸残基组成的碱性蛋白,分别在Cys14-Cys38、Cys5-Cys55和Cys30-Cys51连接半胱氨酸形成3对二硫键。其中1对二硫键(Cys14-Cys38)位于分子表面,其使2段肽链(32~42,9~21)连接在一起,这1对二硫键被还原和重新氧化后不影响抑制剂活力。另外2对二硫键(Cys5-Cys55,Cys30-Cys51)位于分子内部,还原后抑制剂失活。Kunitz家族蛋白酶抑制剂分子空间构相高度保守,有许多共同特征,BPTI整个分子呈梨型,由2条反平行的β-折叠、2段α-螺旋、β-转角和一些环组成,其最主要的结构特征是含有2片180度扭曲的反平行β-折叠,这个折叠分别和β转角与1对二硫键(Cys14-Cys38)相连。这个分子的主要片段1~14和38~58残基沿着β-折叠来回弯曲折叠,以使末端残基1和残基58能够在β转角处相互靠近,二硫键的共价连接也使这两端靠拢并和β转角(30~51残基组成)靠近。N端的残基形成了一个短的螺旋段,C端形成了1个更长的、有规律的α螺旋(48~56残基)。分子的自然折叠和半胱氨酸以独特的方式共价连接使整个分子变得紧凑、稳定[8]。自Kunitz[9]发现BPTI可以抑制激肽酶和胰蛋白酶以来,许多科研工作者就致力于BPTI的研究。1993年12月美国食品药品监督管理局(FDA)批准将BPTI应用于临床心肺胸外科手术中[10],用于减少出血和渗血以及有凝血障碍可能出现大出血情况的患者治疗中。目前国产的BPTI制品主要是从牛肺中提取,BPTI作为异源蛋白具有免疫原性,大量使用可出现过敏反应等不良临床反应。每千克的牛肺组织中仅可提取1mg的BPTI,且提取工艺复杂[11]、产量低、产品的纯度不高。
本实验选用了甲基营养型酵母——毕赤巴斯德酵母(Pichia.pastoris)。毕赤酵母系统的广泛应用,除该系统具有一般酵母所具有的特点外,还具有以下几个优点[12-14]:①具有醇氧化酶AOX1基因启动子,该启动因子是目前调控机制最严格的启动子之一;②表达质粒能在基因组的特定位点以单拷贝或多拷贝的形式稳定整合;③菌株易于进行高密度发酵,外源蛋白表达量高;④毕赤酵母中存在过氧化物酶体,表达的蛋白贮存其中,可免受蛋白酶的降解,而且减少对细胞的毒害作用。经近十年发展毕赤巴斯德酵母基因表达系统已成为较完善的外源基因表达系统,具有易于高密度发酵、表达基因稳定整合在宿主基因组中、能使产物有效分泌并适当糖基化和培养方便经济等特点。毕赤巴斯德酵母表达系统在生物工程领域将发挥越来越重要的作用,促进更多外源基因在该系统的高效表达,提供更为广泛的基因工程产品[15]。
本实验采用pPICZα质粒构建KD/APPvar的分泌型表达质粒,电转化毕赤酵母X-33菌种,构建了高效分泌表达rhKD/APPvar的工程菌,并对毕赤酵母大规模表达rhKD/APPvar的发酵条件进行了优化。纯化过程中,采用pH 3.5条件下进行阳离子交换层析可很好地避免核酸和内毒素的污染,且得到纯度较高的rhKD/APPvar,除盐浓缩后,可直接用于生物活性的鉴定和药效学实验,该纯化方法简单、成本低,适合于大规模纯化。
[1]Posadas SJ,Caz V,Caballero I,et al.Effects of mannoprotein E1in liquid diet on inflammatory response and TLR5expression in the gut of rats infected by Salmonella typhimurium[J].BMC Gastroenterol,2010,10(1):58-65.
[2]Greif F,Ben-ari Z,Taya R,et al.Dual effect of erythropoietin on liver protection and regeneration after subtotal hepatectomy in rats[J].Liver Transpl,2010,16(5):631-638.
[3]Ellett JD,Atkinson C,Evans ZP,et al.Murine Kupffer cells are protective in total hepatic ischemia/reperfusion injury with bowel congestion through IL-10[J].J Immunol,2010,184(10):5849-5858.
[4]王虹蛟,王心童,王 强,等.抑肽酶对实验性慢性肝损伤大鼠肝细胞增殖能力的影响[J].吉林大学学报:医学版,2011,37(1):18-20.
[5]王 强,王心童,王虹蛟,等.抑肽酶对实验性急性肝损伤模型小鼠肝组织的保护作用[J].吉林大学学报:医学版,2010,36(6):1039-1042.
[6]王虹蛟,孟威宏,王 强,等.抑肽酶对实验性慢性肝损伤的保护作用[J].中国实验诊断学,2008,12(1):4-7.
[7]Hocman G.Chemoprevention of cancer:protease inhibitor[J].Int J Biochem,1992,24(3):1365-1375.
[8]Haghara Y,Shirali K,Nakamura T,et al.SCreening for stable mutants with amino acid pairs substituted for the disulfide bond between residues 14and 38of bovine pancreatic trypsin inhibitor(BPTI)[J].J Biol Chem,2002,277(52):51043-51048.
[9]Kunitz M.Isolation of crystalline protein compound of trypsin and of soybean tryps ininhibitor[J].J Gen Physiol,1947,30(4):311-320.
[10]Chesshyre JA,Kraunsoe JA,Lowe G.Production of bovine-pancreatic-trypsin-inhibitor homologues in Escherichia coli and their characterization[J].Biotechnol Appl Biochem,1995,22(3):269-280.
[11]张振龙,李 津.发酵工艺放大的优化[J].国外医学:预防、诊断、治疗生物制品分册,2003,26(2):61-63.
[12]李 晶,赵晓祥,沙长青,等.甲醇酵母基因表达系统的研究进展[J].生物工程进展,1999,19(2):17-20.
[13]Gurramkonda C,Zahid M,Nemani SK,et al.Purification of hepatitis B surface antigen virus-like particles from recombinant Pichia pastoris and in vivo analysis of their immunogenic properties[J].J Chromatogr B Analyt Technol Biomed Life Sci,2013,940(1):104-111.
[14]Schmidt C,Ploier B,Koch B,et al.Analysis of yeast lipid droplet proteome and lipidome[J].MethodsCell Biol,2013,116(1):15-37.
[15]彭 毅,杨希才,康良仪.影响甲醇酵母中外源蛋白表达的因素[J].生物技术通报,2000,30(4):33-36.
