金叶马褂木叶片特性及其RAPD分子鉴定
2014-03-22王立如徐绍清沈生初王忠华
王立如,徐绍清,沈生初,施 佩,王忠华
(1. 浙江省慈溪市林特技术推广中心,浙江 慈溪 315300; 2. 浙江万里学院 生物技术研究所,浙江 宁波 315100)
马褂木,又称为鹅掌楸(LiriodendronchinenseSarg.),为木兰科(Magnoliaceae)鹅掌楸属树种,是珍稀的第三纪孑遗树。该属目前仅存马褂木和北美鹅掌楸(L.tulipifera)两种[1]。在中国的12个省(区)84县(市)有天然分布[2],被列入同级二级珍稀濒危保护树种[3]。另外还有中国著名林木育种学家叶培忠教授通过人工杂交方法育成的杂交马褂木(L.chinense× L.tulipifera)[4]。
北美鹅掌楸为落叶乔木,树高可达60 m,胸径可达3.5 m以上[1]。该树种绿树浓荫,叶形奇特,具有极高的观赏价值。另外,该树种材淡红褐色,轻软适中,纹理清晰,结构细致,轻而强韧,硬度适中,是胶合板的理想原料,也是制家具、缝纫机板、收音机壳与室内装修的良材。
近20年来,国内外众多学者对鹅掌楸属植物进行了大量的研究。国外主要是对北美鹅掌楸进行的研究相对比较深入,无论在表型性状(果长、叶形)、材性、生长量等,还是在蛋白质、DNA分子水平上都做了深入的研究[1,5]。国内对于马褂木的研究主要集中在地理分布特征[2,3]、生殖生物学[6,7]、交配系统[8]、遗传结构[9,10]、材性[11,12]、育种与繁殖[13-17]、光合作用[18,19]和病原菌[20]等方面。以DNA为基础的多态性检验是遗传多样性检测的强有力工具,RAPD分子标记技术是常用方法之一,简单、快速、灵敏。罗光佐等[21]利用RAPD标记,对部分群体马褂木的遗传多样性进行了研究。
本课题组自2000年开始引种鹅掌楸属树种,包括马褂木、北美马褂木、金边马褂木和杂交马褂木等,其中金边马褂木是北美马褂木的栽培品种。金叶马褂木是2002年从金边马褂木的嫁接苗中发现的全叶为金黄色的芽变材料。目前存圃的8年生金叶马褂木平均树高11 m,胸径12 cm,长势健壮。本研究以8年生金叶马褂木、金边马褂木、北美马褂木为实验材料,对金叶马褂木的主要生物学特性进行比较分析,并结合RAPD技术进行分子鉴定,为确认金叶马褂木的遗传变异提供科学依据。
1 材料和方法
1.1 材料
实验材料分别为8年生金叶马褂木、金边马褂木、北美马褂木,样品采自浙江省慈溪市白沙街道农业园区马褂木种植基地。
1.2 方法
1.2.1 生物学特性调查
在金叶马褂木等品种生长初期开始,间隔一定时间用数显游标卡尺和直尺测量各品种在不同时期叶片的宽度、长度,并以相机采集植株叶片图像。用叶绿素测定仪SPAD-502(日本生产)测定各品种在不同时期的叶绿素含量,进行比较分析。
1.2.2 预处理
将新鲜材料洗净擦干后剪碎,放入预冷的研钵中,加入适量的PVP提取液以及2%的β-巯基乙醇进行快速研磨。研磨成均匀的糊状后,取适量分装入1.5 mL的Eppendorf管中,待用。
1.2.3 基因组DNA提取
3个品种的样品基因组DNA的提取采用改良的CTAB法进行[22],其步骤如下:1)向上面待用的EP管中加入800 μL预热的CTAB提取缓冲液,混匀后65℃水浴1 h,12000 r/min离心15 min;2)取上清,加入等体积的氯仿/异戊醇(24∶1)轻轻混匀,12000 r/min离心15 min,重复此步骤1次;3)取上清,加入2/3体积的异丙醇,放入-20 ℃冰箱40 min,沉淀DNA;4)加入200 μL 70%乙醇洗涤DNA 2~3次;5)将1.5 mL Eppendorf管倒置于通风柜里,使DNA尽量干燥;6)加入适量的TE,溶解DNA;7)置于4℃冰箱保存,备用。
1.2.4 DNA纯度和浓度检测
DNA 浓度与纯度的定性检测采用琼脂糖凝胶电泳的方法。取5 μL DNA 样品,在1%的琼脂糖下电泳。DNA 浓度与纯度的定量检测采用紫外分光光度法,DNA 的紫外吸收峰在260 nm 波长处。纯的DNA 的OD260/OD280应该≥1.8。
1.2.5 PCR扩增
选用上海生物工程公司RAPD扩增随机引物,引物长度10 bp,经初步筛选后,选用12个随机引物,引物名称与序列见表1。

表1 PCR反应引物
本实验中,RAPD 反应总体积为20 μL,反应体系见表2。

表2 PCR反应体系
反应程序如下:94 ℃预变性3 min,94 ℃变性45 s,36 ℃退火45 s,72 ℃延伸90 s,共45个循环,最后72 ℃延伸10 min。
1.2.6 PCR产物电泳及显色
取5 μL PCR扩增产物,用1.2 %琼脂糖凝胶上电泳,电泳条件为80 V稳压电泳1 h,溴酚兰作电泳指示剂,缓冲液为1×TBE,紫外凝胶成像仪观察,用凝胶成像设备摄像并通过与DNA标准分子量Marker比较分析结果。
1.3 数据分析
用Excel 2003 统计软件分析和制图,采用DPS软件显著性测定[23]。
2 结果与分析
2.1 叶片大小变化
如图1和图2 所示,3个鹅掌楸属品种叶片生长发育过程基本相同,3月底开始萌发,至5月上旬,为鹅掌楸属品种叶片增大期,5月中旬开始进入缓慢生长期,6月8日金叶马褂木、金边马褂木、北美马褂木叶片宽度、长度分别为13.14 cm/11.62 cm、13.21 cm/11.55 cm、13.85 cm/12.2 cm。由此表明3个鹅掌楸属品种叶片大小差异不大,其中金叶马褂木、金边马褂木叶片比北美马褂木叶片略小。从图1和图2还可看出,在4月16日,北美马褂木叶片长度和宽度都显著高于金叶马褂木和金边马褂木,叶片长度F﹡﹡=32.573≥F0.01=4.02,叶片宽度F﹡﹡=23.457≥F0.01=4.02,而后两者差异不大。这与各品种的基因型差异有关。
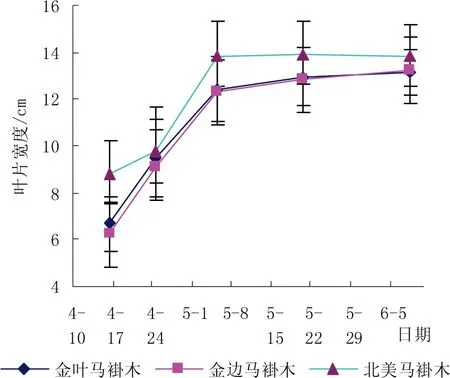
图1 金叶马褂木叶片宽度变化

图2 金叶马褂木叶片长度变化
2.2 叶片叶绿素含量和色泽变化
如图3所示,3个鹅掌楸属品种叶片叶绿素含量变化过程基本相同。即从萌芽展叶至5月上旬,3个鹅掌楸属品种叶片的叶绿素含量都处于递增期,5月中旬开始进入稳定期,5月20日达到最高值。从对三者的比较可看出,金叶马褂木叶片叶绿素含量与金边马褂木叶片周边部分叶绿素含量差异不大,而金叶马褂木叶片叶绿素含量明显少于金边马褂木叶片中间部分和北美马褂木叶片的叶绿素含量,4月16日F**=611.677≥F0.01=3.96;4月24日F**=661.99≥F0.01=3.97;5月5日F**=325.476≥F0.01=3.96;5月9日F**=84.622≥F0.01=4.03;5月20日F**=62.399≥F0.01=4.01;6月8日F**=196.754≥F0.01=3.94,差异达到极显著性水平。

图3 金叶马褂木叶绿素含量变化
A1—2013年4月11日金叶马褂木叶色;A2—2013年4月11日金边马褂木叶色;B1—2013年4月16日金叶马褂木叶色;B2—2013年4月16日金边马褂木叶色;C1—2013年4月24日金叶马褂木叶色;C2—2013年4月24日金边马褂木叶色;D1—2013年5月9日金叶马褂木叶色;D2—2013年5月9日金边马褂木叶色。
图4金叶马褂木与金边马褂木的叶色变化对比
Fig 4 The difference of leaf color changes between tulip tree with golden leaves and with golden edge
2.3 3个鹅掌楸属品种的RAPD分子鉴定
在12个RAPD随机引物PCR扩增结果发现,只有2个RAPD引物OPO16和OPAU19在3个鹅掌楸属品种中扩增出差异性条带(图5)。由图5可见,上述两种引物在金叶马褂木中扩增出与北美马褂木、金边马褂木不同的特异性条带。因此,RAPD引物OPO16和OPAU19可初步作为区分金叶马褂木与北美马褂木、金边马褂木的DNA指纹标记。
1~3泳道分别为北美马褂木(1)、金叶马褂木(2)和金边马褂木(3)样品;M为标记。
图5 RAPD引物OPO16和OPAU19的PCR扩增结果
Fig 5 PCR amplification with primers OPO16 and OPAU19
3 结论与讨论
鹅掌楸属树种树形端正,叶形奇特,是优美的庭荫树和行道树种,其中马褂木与悬铃木(Platanus×acerifolia)、椴树(Tiliasp.)、银杏(Ginkgobiloba)、七叶树(Aesculuschinensis)并称“世界五大行道树种”。花淡黄绿色,美而不艳,最宜植于园林中的安静休息区的上。秋叶呈黄色,很美丽,可独栽或群植,在江南自然风景区中可与木荷(Schimasuperba)、山核桃(Caryacathayansis)、板栗(Castaneamollissima)等行混交林式种植。因其花形酷似郁金香(Tulipagesneriana),故被称为“中国的郁金香树(Chinese Tulip Tree)”,是一种非常珍贵的盆景观赏植物,对SO2等有毒气体有抗性,可在大气污染较严重的地区栽植。
本试验所选的金叶马褂木是金边马褂木的芽变种,其叶片特性与后者有较大的区别,即全叶为金黄色。其叶片大小、发育状况与金边马褂木类似,但叶片叶绿素含量明显低于后者。由此表明,金叶马褂木极有可能是一种新的马褂木种质资源。
RAPD技术可快速、高效地获得许多个或基因型的多位点DNA序列多态性信息,是极其有效的DNA遗传标记系统之一。本试验采用该技术对金叶马褂木进行了初步的分子验证,已成功获得了差异性标记RAPD引物OPO16和OPAU19,为金叶马褂木作为新的种质提供了有力的分子证据。但RAPD技术的扩增结果受到多种因素的影响,如模板DNA、Mg2+、Taq酶、引物以及dNTPs等[24],因此有必要开展马褂木RAPD分子鉴定技术的参数优化研究,为马褂木的种质资源评价与应用提供技术支撑。
参考文献:
[1]王章荣,等编著. 鹅掌楸属树种杂交育种与利用[M]. 北京:中国林业出版社, 2005, 16-36.
[2]郝日明,贺善安,汤诗杰,等.鹅掌楸在中国的自然分布及其特点[J].植物资源与环境学报.1995,4(1):1-6.
[3]贺善安,郝日明.中国鹅掌楸自然种群动态及其致危生境的研究[J].植物生态学报,1999,23(1):88-96.
[4]赵书喜.杂交马褂木的引种与杂种优势利用[J].湖南林业科技,1989(2):20-21.
[5]王章荣.鹅掌楸属杂交育种成就与育种策略[J].林业科技开发,2008,22(5):1-4.
[6]徐 进,王章荣.鹅掌楸属种间杂种与亲本花粉壁的超微结构的比较观察[J].植物学通报,2005,22(1):35-38.
[7]黄双全,郭友好,陈家宽.渐危植物鹅掌楸的授粉率及花粉管生长[J].植物分类学报,1998,36(4):23-29.
[8]李周岐,王章荣.用RAPD标记检测鹅掌楸属种间杂交的花粉污染[J].植物学报,2001,43(12):1271-1274.
[9]董 纯,谭德仁,汪长江,等.马褂木(鹅掌楸)地理种源试验研究报告[J].湖北林业科技,1999(01):3-10.
[10]李 斌,顾万春,夏良放,等.鹅掌楸种源遗传变异和选择评价[J].林业科学研究,2001,14(3):237-244.
[11]李 斌,顾万春,谭德仁,等.鹅掌楸种源材性遗传变异与选择[J].林业科学,2001,37(2):42-50.
[12]季孔庶,王章荣,温小荣.杂交鹅掌楸生长表现及其木材胶合板性能[J].南京林业大学学报:自然科学版,2005,29(1):71-74.
[l3]王章荣.中国马褂木遗传资源的保存与杂交育种前景[J].林业科技通讯,1997(09):8-10.
[14]王章荣.鹅掌楸属(Liriodendron)杂交育种回顾与展望[J].南京林业大学学报:自然科学版,2003,27(3):76-78.
[15]程忠生,毛根松.马褂木种子发芽试验简报[J].浙江林业科技,1997,17(3):20-25.
[16]陈金慧,张艳娟,李婷婷,等.杂交鹅掌楸体胚发生过程的起源及发育过程[J].南京林业大学学报: 自然科学版,2012,36(1):16-20.
[17]叶金山,季孔庶.杂种马褂木无性系插条生根能力的遗传变异[J].南京林业大学学报:自然科学版,1998,22(2):71-74.
[18]杨秀艳,季孔庶,王章荣,等.杂交鹅掌楸苗期光合特性的研究[J].西北林学院学报,2005,20(02):39-43.
[19]郭志华,张宏达,李志安,等.鹅掌楸(Liriodendronchinense)苗期光合特性的研究[J].生态学报,1999,19(2):22-27.
[20]王记祥,马良进,张立钦,等.马褂木褐斑病病原的鉴定[J].林业科学,2013,49(6):189-191.
[21]罗光佐,施季森,尹终明,等.利用RAPD标记分析北美鹅掌楸与鹅掌楸种间遗传多样性[J].植物资源与环境学报,2000,9(2):9-13.
[22]王廷华,景 强,Pierre D.PCR理论与技术[M].北京:科学出版社,2006, 13.
[23]唐启义,冯明光编著.实用统计分析及其DPS数据处理系统[M]. 北京:科学出版社, 2002:48-53.
[24]李建民,周志春,吴开云,等.RAPD标记研究马褂木地理种群的遗传分化[J].林业科学,2002,38(4):61-66.
