青海湖区大型底栖动物群落结构与空间分布格局
2014-03-17孟星亮何玉邦宋卓彦熬鸿毅张虎蒋小明
孟星亮何玉邦宋卓彦熬鸿毅张 虎蒋小明
(1.中国科学院水生生物研究所, 武汉 430072; 2. 中国科学院大学, 北京 100049; 3. 青海湖国家级自然保护区管理局, 西宁 810008)
青海湖区大型底栖动物群落结构与空间分布格局
孟星亮1,2何玉邦3宋卓彦1,2熬鸿毅1张 虎3蒋小明1
(1.中国科学院水生生物研究所, 武汉 430072; 2. 中国科学院大学, 北京 100049; 3. 青海湖国家级自然保护区管理局, 西宁 810008)
为了解青海湖大型底栖动物的群落结构特征、空间分布格局及关键驱动力, 于2011—2012年分4次对青海湖区主体湖、3个子湖和5条入湖河流进行了系统调查。共采集到底栖动物28个分类单元, 隶属于3门5纲15科, 主体湖区、咸水和淡水子湖及入湖河流分布有9、5、9和15种; 它们的密度分别为455.8、1562.7、741.3和331.2 ind./m2; 生物量为1.18、3.70、2.75和4.53 g/m2。优势度分析显示, 喜盐摇蚊为主体湖区绝对优势种(占总密度的83.32%)。Surfer模拟显示, 主体湖区东南部沿岸带的密度和生物量最高, 湖心区最低; 高峰值主要出现在布哈河、菜挤河等大型河流入湖的河口附近。CCA分析表明: 青海湖区大型底栖动物的分布主要受水体pH、水深、总氮和盐度等环境因子的驱动。
大型底栖动物; 空间格局; 青海湖; 多因子分析
青海湖是我国最大的高原咸水湖, 由于其独特的高原环境和丰富的生物多样性, 在1992年被列入拉姆莎尔保护公约中著名的湿地保护区, 同时分别在1997、2006和2007年被国务院和国家建设部批准为国家级自然保护区、国家自然名录以及国家级水产种质资源保护区[1]。然而近50年来, 由于受气候变暖和人类活动影响, 青海湖出现了一系列的生态问题, 如水位下降、沙漠化加剧、草场面积退化、鱼类资源骤减等, 这促使国内外学者对青海湖现状愈加关注[2]。目前为止, 国内有关青海湖的研究主要集中在气候变化[3,4]、水位状况[5,6]、鱼类资源[7]和水生植物[8]等领域。但迄今为止, 有关该湖底栖动物的研究却相对较少, 现有研究也大多仅集中于种类描述[1,9], 而罕有关于底栖动物群落结构和格局的研究。
大型底栖动物是湖泊生态系统中的重要生态类群, 在促进水体生态系统的营养物质循环和能量流动、调节底质与水体的物质交换以及加速水体净化、改善水体微环境等方面起着十分重要的作用[10—13]。此外, 底栖动物作为鱼类、鸟类等动物的天然饵料,其群落结构的变化直接影响着高营养级动物的生长和繁殖[14]。鉴于其高原咸水湖泊的特殊性及大型底栖动物的未知现状, 本研究于2011年6月初和8月底以及2012年6月中旬和8月底, 分4个时间段对青海湖及其周边水体的底栖动物进行了调查, 以期阐明该湖区底栖动物群落结构和空间分布现状, 为青海湖湖区管理提供科学依据。
1 材料与方法
1.1 样点设置与湖泊概况
湖泊概况 青海湖 (N36°32′-37°15′, E99°36′-100°47′) 又名“库库淖尔”, 位于青海省东北部青海湖盆地。主体湖水域面积 4340.0 km2(平均高程3193.92 m), 平均湖长为109.0 km, 平均湖宽39.8 km (最大宽67.0 km), 平均水深17.9 m (最大水深27.0 m)。湖区气候属高原高寒半干旱气候, 年均气温 1.2 , ℃多年平均降水量336 mm。青海湖于每年11月进入冰封期至次年3月开始解冻, 冰期历程100—129d。湖水主要依赖地表径流和湖面降水补给, 主要入湖河流有布哈河、巴戈乌兰河、沙柳河、黑马河、甘子河、倒淌河等, 其径流量约占入湖总径流量的95%。布哈河最大, 长约286 km, 流域面积14384 km2,其径流量约占入湖总径流量的45%[15]。其中金沙湾部分与主体湖区季节性连通, 尕海与主体湖完全隔离且无明显河流补给, 因而现在这两个子湖仍然呈现咸水湖泊特征; 洱海与主体湖完全隔离且有倒淌河补给, 如今已演变成为一个淡水子湖。

图1 青海湖及其附近水体底栖动物和水体理化参数的样点分布Fig. 1 Sampling sites of benthic organisms and physical-chemical parameters in Qinghai Lake
样点设置 样点布设上综合考虑了主体湖区、2 个咸水子湖、1 个淡水子湖及入湖河流4 类不同盐度类型生态区域。主体湖区大体沿用了1962年样点 (图 1A), 布设 17个采样点(QHH01-18, 除JSW14外)。入湖河流设置了5个采样点, 其中黑马河、布哈河、泉吉河、沙柳河和菜挤河样点分别对应HMH20、BHH21、QJH22、SLH23和CJH24。子湖设置了3个采样点, JSW14、GH19和EH25分别对应金沙湾、尕海和洱海3个子湖 (图1B)。
1.2 样品的采集与处理
主体湖区使用 1/16 m2的改良 Peterson 采泥器进行底栖动物样品采集, 每个样点采集2次, 泥样经40目的筛网筛洗后, 置于密封袋内带于室内, 当天置入白色解剖盘中分拣。随后置入50 mL样品瓶中,并用 10%的福尔马林固定, 带回实验室进行种类鉴定、计数和称湿重(其中软体动物为带壳湿重)。入湖河流和子湖则使用0.09 m2的Surber网进行, 将采得样品置于40目的分样筛中筛洗, 此后处理方法同上。
1.3 水体理化指标的测定
用5 L采水器采取表层水样(0.5 m), 当天带回实验室测定。碱度(ALK)、化学需氧(COD)、总氮(TN)、氨氮(NH4-N)、亚硝酸氮(NO2-N)、硝氮(NO3-N)、总磷(TP)、磷酸盐(PO4-P)测定方法参照《水和废水监测分析方法》[16]。其他指标温度(T)、pH、溶解氧(DO)、水深(H)、电导率(EC)、盐度(SAL)在现场使用便携式水质分析仪测定, 透明度(SD)用Secchi盘测定。
1.4 数据处理
以底栖动物相对密度≥5%作为确定优势种的标准[17, 18]。
使用单因素方差分析(One-way ANOVA)比较4类水体的理化因子、底栖动物密度、生物量、物种数和多样性指数间的差异。为提高数据的正态性和方差齐性, 理化因子(pH、溶解氧和氨氮除外)与Shannon-Wiener 和Margalef指数分别进行lg(x + 1)转换。若单因素方差分析的组间差异显著, 则进行多重比较; 如果方差同质, 则进行 LSD (the least significant difference)检验; 如果方差不具同质性, 就选用Games-Howell检验[19], 以上处理均在SPSS 13.0软件中完成。
底栖动物 Shannon-Wiener和 Margalef 多样性指数计算采用以下公式:

Margalef 多样性指数: Margalef index (d) = (S–1)/lnN,其中, H ′: Shannon-Wiener 多样性指数; S: 样本中总分类单元数; Pi: 物种 i在样本中的相对丰度; N:样本中总个体数。
本次调查主要在主体湖区, 因而作者着重分析了主体湖区底栖动物的密度和生物量空间分布格局,空间分布图使用普通克里格插值法, 在Surfer 8.0软件中完成[20]。
探讨底栖动物分布和水环境因子的关系采用 R软件(2.15版本), “vegan” 软件包完成。首先, 对25个样点4次水体理化因子和物种数据分别进行平均,然后分别对非正态分布的环境因子(pH、氨氮和矿化度除外)分别进行 lg (x+1)转换。接着利用函数permutest( )进行 Monte Carlo 转置, 检验环境因子与物种分布间的相关性。其次使用envfit( )涵数检验每个环境因子的相关性, 然后将物种丰富度与环境因子不相关的(P>0.05)环境因子剔除, 同时利用vif.cca( )函数剔除较高变异波动系数(>20)的变量。随后分别对物种数据进行 lg (x+1) 转换, 保留在各样点出现的频度>10%且相对密度≥1%的物种, 以消除稀有种的影响[21—22]。最后使用decorana( )函数对底栖动物群落分布进行除趋势对应分析 (DCA),显示单峰模型 (gradient length > 4 standard units) 更为适合群落组成与环境因子之间关系的分析,因此采用cca( )函数进行了典范对应分析(CCA)[23,24]。
2 结果
2.1 青海湖湖区环境因子特征
在4类水体中, 多数环境因子差异显著(表1)。其中, SAL、EC、MIN、ALK表现为咸水子湖>主体湖>淡水子湖和河流; 而 SiO2、TP、NO3-N则为入湖河流>淡水子湖>主体湖区和咸水子湖; TN 和CODMn在淡水子湖出现最高值, 河流和咸水子湖居中, 而在主体湖区最低。总体而言, 主体湖区的营养盐浓度处于贫营养水平[25]; 而 3个子湖和入湖河流的营养盐浓度相对较高。
2.2 青海湖湖区底栖动物群落结构
本文共记录28个分类单元, 隶属于3门5纲15科, 其中主体湖区9种, 淡水子湖9种, 咸水子湖5种, 入湖河流 15种(表 2)。主体湖、咸水子湖、淡水子湖和入湖河流的平均密度分别为455.8、1562.7、741.3和 331.2 ind./m2; 平均生物量分别为 1.18、3.70、2.75和4.53 g/m2(图2)。
密度、生物量和物种数在4类水体间差异不显著 (P > 0.05)。相对而言, 在密度方面咸水子湖最高,淡水子湖次之, 河流和主体湖最低; 生物量方面河流和咸水子湖较高, 主体湖最低, 淡水子湖居中;物种数河流和淡水子湖较高, 主体湖与咸水子湖较低(图 2)。

表1 2011—2012年青海湖湖区主要水质参数Tab. 1 The water quality parameters in Qinghai Lake area from 2011 to 2012 (mean ± SD)
群落组成方面 4类水体差异较大, 主体湖区以喜盐摇蚊(Chironomus salinarius)、前突摇蚊(Procladius sp.)和钩虾(Gammarus sp.)为优势种, 三者相对密度分别为83.32%、10.58%和5.11%, 占总生物量的百分比分别为 81.49%、7.91%、10.6%。3个子湖主要以水生昆虫为主, 其中优势种类均为蠓(Ceratopogonidae spp.)及水甲(Hygrobiidae spp.), 二者在咸水和淡水子湖相对密度分别为 80.55%、18.77%和79.82%、19.47%。而入湖河流的优势种为
钩虾、扁蜉(Heptageniidae spp.)、特氏直突摇蚊(Orthocladious thienemanni)和大蚊 (Tipulidae spp.),相对密度分别为62.80%、9.18%、8.21%、7.25%。

表2 青海湖湖区底栖动物物种名录Tab. 2 List of the species of macroinvertebrates in Qinghai Lake area

图2 四类水体平均密(A)、生物量(B)和物种数(C)的比较(Mean±SD)Fig. 2 Comparison of density, biomass and the number of the species between 4 types of water bodies in Qinghai Lake area Abbreviation: MB: main body; SAL: saline adjacent lake; FAL: freshwater adjacent lake; Irs: inflowing rivers
2.3 青海湖湖区底栖动物生物多样性
4类水体的多样性较低(图 3)。Shannon-Wiener指数在4类水体之间差异不明显(P>0.05); Margalef指数 4类水体之间相比, 主体湖区、咸水子湖与河流间差异显著(P<0.05)。相对而言河流和淡水子湖多样性较高, 咸水子湖和主体湖较低。
2.4 主体湖区大型底栖动物空间分布
底栖动物的密度和生物量呈现大体一致的空间分布格局, 表现为西南部大于东北部 (图4a、b);两者从湖心区域依次向沿岸区域逐渐增加, 最高点位于西南角菜挤河河口附近, 密度和生物量分别达2000 ind./m2、4 g/m2以上, 次高点位于泉吉河河口(800 ind./m2和 2.5 g/m2); 优势种喜盐摇蚊与底栖动物总体的密度和生物量空间分布大体一致(图4c、d)。2.5 青海湖湖区底栖动物与水环境关系
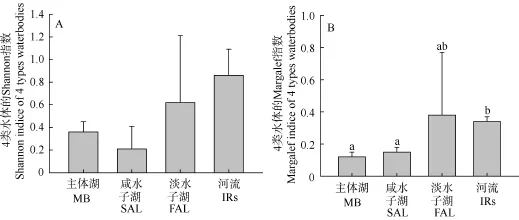
图3 青海湖区4类水体Shannon (A)和Margalef (B)指数的比较(Mean±SD)Fig. 3 Comparison of Shannon and Margalef Indice between the 4 types waterbodies in Qinghai Lake area Abbreviation: MB: main body; SAL: saline adjacent lake; FAL: freshwater adjacent lake; Irs: inflowing rivers

图4 青海湖大型底栖动物总体的密度(a)和生物量(b)及优势种喜盐摇蚊的密度(c)和生物量(d)的空间分布Fig. 4 Density (a) and biomass (b) spatial distribution of total macrozoobenthos and density (c) and biomass (d) space distribution of Chiroindmus salinarius in Qinghai Lake
利用 envift()函数对 15个环境因子进行筛选,最后有7个关键环境因子进入CCA模型(表3)。主要环境因子的解释率为65.2%, CCA 第一和第二排序轴的解释率分别为 24.5%和 16.6%, 前两轴累积解释率为 41.1%。其中 pH (r2=0.8683)、水深(r2= 0.8527)、总氮(r2=0.7762)和硝氮(r2=0.7542)与底栖动物群落分布显著相关, 其次是盐度、矿化度和温度(表3)。

表3 青海湖湖区底栖动物与关键环境因子Monte Carlo检验结果Tab. 3 Results of Monte Carlo test between the key environmental factors and the distribution of the zoobenthos in Qinghai Lake area
水深、pH和盐度与CCA第一排序轴呈显著负相关, 矿化度、总氮和硝氮与第一轴呈显著正相关; CCA第二排序轴则与温度和盐度呈显著正相关。物种与环境因子相互关系表明, 水深、pH和盐度显著影响了喜盐摇蚊、前突摇蚊(Procladius sp.)等物种的空间分布(图 5)。水甲(Hygrobiidae spp.)、蠓(Ceratopogonidae spp.)和梯形多足摇蚊(P. scalaenum)喜温的昆虫与温度和总氮正相关。大蚊 (Tipulidae spp.)和带襀(Taenionema sp.)等物种与盐度呈负相关,与硝氮呈正相关, 这与该两个物种偏好在淡水、营养盐类相对较为丰富的溪流中生活相吻合(图5)。
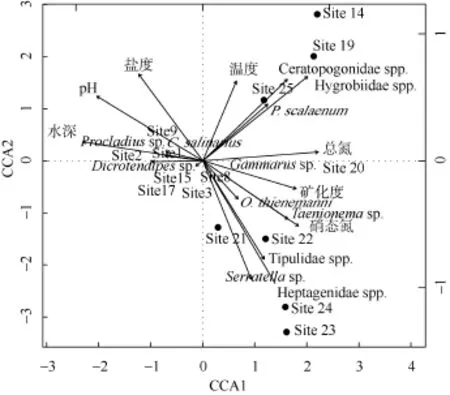
图5 青海湖底栖动物群落结构与水环境关系的CCA分析Fig. 5 The CCA analysis between macrozoobenthos and the key environmental factors in Qinghai lake
3 讨论
3.1 青海湖湖区大型底栖动物历史动态
青海湖主体湖区的底栖动物物种组成单一、群落结构也较为简单, 主要以摇蚊为主, 这与我国北方许多盐碱湖泊的底栖动物群落组成较为相似[26,34]。就主体湖区而言, 与20世纪60年代相比, 本研究底栖动物的总分类单元减少了10个, 科级水平减少了6个科, 属级水平减少了 10个属, 其中软体动物下降了3种、寡毛类下降了2种、其他水生昆虫下降了4种、摇蚊类下降了1种; 平均生物量和密度略有上升[27]; 而对摇蚊类而言, 本此调查与20世纪80年代减少了1个属5个分类单元[28](表4)。就整个青海湖区而言, 底栖动物的物种组成虽然比20世纪90年代在科级水平略有提升, 但其减少了 5属 16个分类单元[29]; 摇蚊类与20世纪80年代相比减少了9属[28](表4)。
青海湖底栖动物方面的研究资料极度匮乏, 就本文数据, 这50年来青海湖底栖动物生物多样性呈现下降的态势。这可能一方面与盐度、水域面积变化等有关, 另一方面可能与本研究对河流及子湖调查力度较为薄弱有关[30,31]。
3.2 青海湖底栖动物与水环境之间的关系
许多研究表明, 水深是影响静水生态系统底栖动物群落结构的关键因子, 水深的改变会导致一些重要理化因子(如温度和溶氧等)发生变化, 从而间接影响底栖动物的分布[32—34]。主体湖Surfer模拟结果显示, 底栖动物密度和生物量的空间分布呈现,从西北部沿岸带向湖心深水区逐渐递减的趋势, 且水深较浅的西南部底栖动物的密度和生物量远高于水深较深的东北部。此外, 湖心带相比湖泊沿岸带通常具有较高的底栖动物多样性和生产力, 因为沿岸带丰富的有机质来源为底栖动物提供了充沛的食物, 更加有利于底栖动物生存[35]。

表4 不同阶段青海湖调查数据Tab. 4 The data of Qinghai Lake in different investigation periods
pH是另一个显著影响底栖动物分布格局的环境因子, 谢祚浑等[36]研究我国北方盐碱水域中的底栖动物指出, 不同的底栖动物对酸碱度的耐受不同,如羽摇蚊(C. plumosus)、前突摇蚊和蠓科幼虫能够在pH 9—10内的水体中生存, 然而其他种类大多生活于pH 7—9内的水体内。这与本文CCA分析结果一致, 扁蜉、特氏直突摇蚊、大蚊等物种与pH呈负相关, 表明这些物种对pH的耐受范围较窄, 因而它们大多分布在pH相对较低的入湖河流中; 相反, 喜盐摇蚊、前突摇蚊及蠓等物种大多分布于主体湖和咸水子湖中。
通常在咸水及半咸水的水体中, 盐度往往成为影响底栖动物群落组成与分布的关键因素, 随着盐度的增加, 底栖动物的物种多样性将逐渐降低, 而在水体盐度值界于 5‰—8‰时, 种类组成呈现单一化的趋势[37,38]。另外, 不同种类的底栖动物对于盐度的耐受范围也差异很大: 如喜盐摇蚊作为咸水水体的广布物种, 其可在 5‰—50‰盐度范围内生存,花纹前突摇蚊(P. Choreus)和蠓科的一些种类均可耐受10‰以上的盐度; 而椭圆萝卜螺(Radix swinhoei)和扁旋螺(Gyraulus compressus) 等螺类大多只能在低于6‰ 盐度的水体中生存[36,39,40]。本文调查发现,金沙湾、尕海及主体湖盐度与河流和子湖洱海差异明显, 因而洱海和河流的物种数目、多样性指数相比主体湖及2个咸水子湖较高。青海湖是典型的高原深水型咸水湖泊, 水体中大量盐份的存在, 限制了其他物种的入侵与生存, 致使河流及附近淡水湖泊中较为丰富的底栖动物种类无法扩散到主体湖,却为喜盐的喜盐摇蚊和前突摇蚊等物种提供了良好的生存环境。因此, 盐度可能是导致青海湖底栖动物物种数目单一的重要原因。
致谢:
感谢中国科学院水生生物研究所王丑明在采样和标本鉴定工作中给予的帮助。
[1] Yao W Z. Study on the resource of feed organisms in Qinghai Lake during summer [D]. Southwest University PH.D thesis. 2011 [姚维志. 青海湖夏季饵料生物资源研究.西南大学博士学位论文. 2011]
[2] Chen G C, Peng M, Zhou L H, et al. Relationship between ecological environment change and human activity in Qinghai Lake region: A preliminary study [J]. Chinese Journal of Ecology, 1994, 13(2): 44—49 [陈桂琛, 彭敏, 周立华, 等. 青海湖地区生态环境演变与人类关系的初步研究. 生态学杂志, 1994, 13(2): 44—49]
[3] Feng S, Tang M C, Zhou L S. The water level of Qinghai Lake change in recent 600 years [J]. Journal of Lake Sciences, 2000, 12(3): 205—210 [冯松, 汤懋苍, 周陆生.青海湖近 600年的水位变化. 湖泊科学, 2000, 12(3): 205—210]
[4] Zhen E L, Shen J, Wang S M, et al. Lake sediment records of climate and environment evolution of Qinghai Lake in recent 900 years [J]. Journal of Lake Sciences, 2002, 14(1):32—38 [张恩楼, 沈吉, 王苏民, 等. 青海湖近900年来气候环境演化的湖泊沉积记录. 湖泊科学, 2002, 14(1): 32—38]
[5] Ma Y. Correlation analysis between the changing of water level and the climate elements of Qinghai Lake [J]. Journal of Lake Sciences, 1996, 8(2): 103—106 [马钰. 青海湖水位变化与湖区气候要素的相关分析. 湖泊科学, 1996, 8(2): 103—106]
[6] Yang H Y, Jia S F. Water balance and water resources allocation of Qinghai Lake [J]. Journal of Lake Sciences, 2003, 15(1): 35—40 [燕华云, 贾绍风. 青海湖水量平衡分析与水资源优化配置研究. 湖泊科学, 2003, 15(1): 35—40]
[7] Hu A, Zhu S Q, Wang J L, et al. Fish Fauna of Qinghai Lake and the Biology of Gymnocypris przewalskii [M]. Beijing: Science Press. 1975, 94 [青海省生物研究所编. 青海湖地区的鱼类区系和青海湖裸鲤的生物学. 北京: 科学出版社. 1975, 94]
[8] Chen Y D. Studies on the Potamogetonaceae in Qinghai Lake [J]. Acta Hydrobiologica Sinica, 1987, 11(3): 228—235 [陈耀东. 青海湖眼子菜科植物的研究. 水生生物学报, 1987, 11(3): 228—235]
[9] Yang J X, Qi H F, Si J Q, et al. The summer investigation of aquatic organisms in Qinghai Lake [J]. Qinghai Science and Technology, 2008, 6: 19—24 [杨建新, 祁洪芳, 史建全, 等.青海湖夏季水生生物调查. 青海科技, 2008, 6: 19—24]
[10] Covich A P, Palmer M A, Crowl T A. The role of benthic invertebrate species in freshwater ecosystems: zoobenthic species influence energy flows and nutrient cycling [J]. BioScience, 1999, 49(2): 119—127
[11] Lindegaard C. The role of zoobenthos in energy flow in deep, oligotrophic Lake Thingvallavatn, Iceland [J]. Hydrobiologia, 1992, 243(1): 185—195
[12] Lindegaard C. The role of zoobenthos in energy flow in two shallow lakes [J]. Hydrobiologia, 1994, 275(1): 313—322
[13] Bonsdorff E. Drifting algae and zoobenthos effects on settling and community structure [J]. Netherlands Journal of Sea Research, 1992, 30: 57—62
[14] Schilling E G, Loftin C S, Huryn A D. Effects of introduced fish on macroinvertebrate communities in historically fishless headwater and kettle lakes [J]. Biological Conservation, 2009, 142(12): 3030—3038
[15] Wang S M, Dou H S. Lakes in China [M]. Beijing: Science Press. 1998, 472—473 [王苏民, 窦鸿身. 中国湖泊志. 北京: 科学出版社. 1998, 472—473]
[16] The Editor of Water and Wastewater Monitoring Analysis Method. Water and Wastewater Monitoring Analysis Method (4th edition) [M]. Beijing: China Environmental Science Press. 2002, 211—279 [国家环保局, 水和废水监测分析方法编委会编. 水和废水监测分析方法(第四版)北京: 中国环境科学出版社. 2002, 211—279]
[17] Bunn S E, Edward D H, Loneragan N R. Spatial and temporal variation in the macroinvertebrate fauna of streams of the northern jarrah forest, Western Australia: community structure [J]. Freshwater Biology, 1986, 16: 67—92
[18] Xiong J, Jiang X M, Wang C M, et al. Community variation of macrozoobenthos and bioassessment of Dongqian Lake, Ningbo [J]. Research of Environmental Sciences, 2012, 25(3): 282—289 [熊晶, 蒋小明, 王丑明, 等. 宁波东钱湖大型底栖动物群落动态及水质生物学评价. 环境科学研究, 2012, 25(3): 282—289]
[19] Beckmann C F, DeLuca M, Devlin J T, et al. Investigations into resting-state connectivity using independent component analysis [J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 2005, 360: 1001—1013.
[20] Zeng Z X, Chen H X. How to use Sufer 8.0 to draw isoline map [J]. Guangdong Meteorology, 2006, 3: 64—65 [曾志雄,陈慧娴. 如何使用Sufer 8.0画等值线图. 广东气象, 2006, 3: 64—65]
[21] Flores L N, Barone R. Phytoplankton dynamics in two reservoirs with different trophic state (Lake Rosamarina and Lake Arancio, Sicily, Italy) [J]. Hydrobiologia, 1998, 369/370: 163—178
[22] Muylaert K, Sabbe K, Vyverman W. Spatial and temporal dynamics of phytoplankton communities in a freshwater tidal estuary (Schelde, Belgium) [J]. Estuarine, Coasta and Shelf Science, 2000, 50: 673— 687
[23] Leps J, Smilauer P. Multivariate Analysis of Ecological Data Using CANOCO [M]. University of Cambridge Press. 2003, 51
[24] Lai J S, Mi X C. Ordination analysis of Ecological data using Vegan package in R [J]. Advances in Biodiversity Conservation and Research in China IX, 2005, 332—343 [赖江山, 米湘成. 基于Vegan软件包的生态学数据排序分析.中国生物多样性保护与研究进展IX, 2005, 332—343]
[25] Wang M C, Liu X Q, Zhang J H. Evaluate method and classification standard on lake eutrophication [J]. Environmental Monitoring in China, 2002, 18(5): 47—49 [王明翠, 刘雪芹, 张建辉. 湖泊富营养化评价方法及分级标准. 环境科学研究, 2002, 18(5): 47—49]
[26] Lan C J, Shen Y, Wang B X, et al. Investigation of plants and benthic macroinvertebrates of in inner Mongolia-Xinjiang Plateau [J]. Journal of Lake Science, 2010, 22(6): 888—893 [兰策介, 沈元, 王备新, 等. 蒙新高原湖泊高等水生植物和大型底栖无脊椎动物调查. 湖泊科学, 2010, 22(6): 888—893]
[27] Lanzhou Institute of Geology CIS. The Report of Integrated Investigation on Qinghai Lake [M]. Beijing: Science Press. 1979, 50—54 [中国科学院兰州地质研究所等编. 青海湖综合考察报告. 北京: 科学出版社.1979, 50—54]
[28] Ye C J. The larvae of some Tendipedidae midge (Diptera) from Qinghai Lake and its’ river system [J]. Acta Biologica Plateau Sinica, 1984, 2: 67—74 [叶沧江. 青海湖及其附属水系的摇蚊幼虫. 高原生物学集刊, 1984, 2: 67—74]
[29] Yang H Z, Wang J L. Analysis on the benthos and their productivity in Qinghai Lake [J]. Qinghai Science and Technology, 1997, 4(3): 36—39 [杨洪志, 王基琳. 青海湖底栖生物及其生产力分析. 青海科技, 1997, 4(3): 36—39]
[30] Shen F, Kuang D B. Remote sensing investigation and survey of Qinghai Lake in the past 25 years [J]. Journal of Lake Sciences, 2003, 15(4): 289—296 [沈芳, 匡定波. 青海湖最近 25年变化的遥感调查与研究. 湖泊科学, 2003, 15(4): 289—296]
[31] Tian R W. The hydrologic condations of the Qinghai Lake [J]. Tansactions of Oceanology and Limnology, 1988, 4: 18—25 [田任午. 青海湖的水文概况. 海洋湖沼通报. 1988, 4: 18—25]
[32] Petridis D, Sinis A. Benthic maerofauna of Tavropos Reservoir (central Greece) [J]. Hydrobiologia, 1993, 262: 1—12
[33] Baudo R, Ochhipinti A, Nocentini A M.Benthos of Lake Orta in the year 1996 [J]. Journal of Limnology, 2001, 60(2) : 241—248
[34] Zhang C W, Zhang T L, Zhu T B, et al. Community structure of Macrozoobenthos and its relationship with environmental factors in lake Hongze [J]. Journal of Hydroecology, 2012, 33(3): 27—33 [张超文, 张堂林, 朱挺兵, 等. 洪泽湖大型底栖动物群落结构及其与环境因子关系. 水生态学杂志, 2012, 33(3): 27—33]
[35] Horne A J, Goldman C R (editors). Limnology (2nd edition) [M]. McGraw-Hill Inc, New York. 1994, 17—19
[36] Xie Z H, Zhou Y B. Zoobenthos in inland saline waters from autonomous region northern parts of China [J]. Journal of Dalian Fisheries University, 2002, 17(3): 176—186 [谢祚浑,周一兵. 中国北方盐碱水域中的底栖动物. 大连水产学院学报, 2002, 17(3): 176—186]
[37] Cartier V, Claret C, Garnier R, et al. How salinity affects life cycle of a brackish water species, Chironomus salinarius KIEFFER (Diptera: Chironomidae) [J]. Journal of Experimental Marine Biology and Ecology, 2011, 405: 93—98
[38] Williams W D. Salinity as a determinant of salt lake fauna: a question of scale [J]. Hydrobiologia, 1990, 197: 257—266
[39] Gainey Jr. L F, Greenberg M J. Physiological basis of the species abundance-salinity relationship in Molluscs: a speculation [J]. Marine Biology, 1977, 40: 41—49
[40] Wang B Q, Xue J Z, Liu L, et al. Community structure of mcacrobenthos in different salt ponds of Altay prefecture, Xinjiang [J]. Arid Land Geography, 2012, 35(2): 265—273 [王宝强, 薛俊增, 刘婧, 等. 新疆阿勒泰地区不同盐池大型底栖动物的群落结构. 干旱区地理, 2012, 35(2): 265—273]
STRUTURE AND SPATIAL DISTRIBUTION PATTERNS OF MACROZOOBENTHOS IN QINGHAI LAKE AREA
MENG Xing-Liang1,2, HE Yu-Bang3, SONG Zhuo-Yan1,2, AO Hong-Yi1, ZHANG Hu3and JIANG Xiao-Ming1
(1. Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China; 3. Qinghai Lake National Nature Reserve Administration Bureau, Xining 810008, China)
To study macrozoobenthos community structures, distributions and key underlying drive forces of the Qinghai Lake (QL), one of typical saline lakes in Tibetan Plateau, the samples of benthic animals were taken via four different times during 2011—2012 from QL area, including main body (MB), 3 adjacent lakes (ALs) and 5 inflowing rivers (IRs). We identified 28 taxa: 9 in MB, 5 in 2 saline ALs, 9 in 1 freshwater AL, and 15 in IRs. The mean density of 4 types of water bodies were 455.8, 1562.7, 741.3, and 331.2 ind./m2, respectively. Biomass of these 4 different water bodies were 1.18, 3.70, 2.75, and 4.53 g/m2respectively. Chiroinomus salinarius was the major species (83.32% of the total abundance) in MB. The general spatial distribution pattern was supported by the Surfer Model, indicating that the highest value of macrozoobenthos abundance and biomass occurred in the southeast littoral zone of MB, especially in the sites near estuaries of IRs (e.g., Buha, Caiji and Quanji Rivers), and that the lowest value was in the profundal zone of MB. The CCA analysis showed that pH, water depth, total nitrogen, and salinity were the key environmental factors to affect macrozoobenthos community variance.
Macrozoobenthos; Spatial pattern; Qinghai Lake; Multivariate analysis
Q145+.2
A
1000-3207(2014)05-0819-09
10.7541/2014.123
2013-06-26;
2013-11-28
国家科技基础条件平台重点项目“青海湖综合研究专题数据库”(BSDN2009-18)资助
孟星亮(1982—), 男, 河南汤阴人; 博士研究生; 主要从事底栖动物多样性及分类学研究。E-mail: xingliang01@163.com
蒋小明(1983—), 男, 助理研究员; 主要从事底栖动物多样性及分类学研究。E-mail: jxm123.83@163.com
