草鱼细胞系CIK酵母双杂交cDNA文库的构建及初步应用
2014-03-17闫秀英谢吉国李杰丁燏吴灶和鲁义善简纪常
闫秀英谢吉国李杰丁燏吴灶和鲁义善简纪常
(1. 广东海洋大学水产学院 广东省水产经济动物病原生物学及流行病学重点实验室暨广东省高等学校水产经济动物病害控制重点实验室,湛江 524088;2. 仲恺农业工程学院,广州 510225)
草鱼细胞系CIK酵母双杂交cDNA文库的构建及初步应用
闫秀英1谢吉国1李杰1丁燏1吴灶和2鲁义善1简纪常1
(1. 广东海洋大学水产学院 广东省水产经济动物病原生物学及流行病学重点实验室暨广东省高等学校水产经济动物病害控制重点实验室,湛江 524088;2. 仲恺农业工程学院,广州 510225)
旨在探索草鱼呼肠孤病毒与宿主细胞蛋白质间的相互作用,运用SMART技术构建了草鱼肾组织细胞系CIK(Ctenopharyngodon idellus kidney)的酵母双杂交cDNA文库。提取CIK细胞总RNA后,分离纯化mRNA,然后以mRNA为模板,反转录合成cDNA第一链,再在DNA聚合酶作用下,通过长距离PCR,扩增双链cDNA。利用SMART技术,通过同源重组的方法,在酵母株Y187中构建了草鱼CIK细胞全长cDNA文库。经检测,未扩增文库的转化率为1.6×105,文库容量为2.4×106,插入的双链cDNA片段的长度为250-2 000 bp,文库滴度为7×107CFU/mL,重组率为98%,此文库具有良好的cDNA片段多态性和完整性。利用构建的CIK酵母双杂交文库,以草鱼呼肠孤病毒的VP7和VP5蛋白作为诱饵进行筛选试验,得到VP7相互作用蛋白的阳性菌落,未得到VP5相互作用蛋白的阳性菌落。草鱼CIK细胞酵母双杂交cDNA文库的构建为研究草鱼呼肠孤病毒与宿主细胞间的互作机制提供了重要研究工具。
草鱼肾组织细胞系 酵母双杂交 cDNA文库 蛋白质互作
草鱼呼肠孤病毒(Grass carp reovirus,GCRV)是草鱼出血病的病原,给草鱼养殖业的健康发展造成了严重的损失[1]。目前,研究人员已对呼肠孤病毒的形态[2-4]、流行病学[4]、检测[5,6]、生物学特性[7]等方面进行了研究,但GCRV的感染机制和致病机制知之甚少,GCRV蛋白的功能有待进一步研究[8-10]。草鱼肾组织细胞系CIK(Ctenopharyngodon idelluskidney)来源于草鱼的肾脏组织[11],是GCRV及多种病毒的收纳细胞系[12],广泛应用于呼肠孤病毒分子生物学的各种试验研究[13-16]。
借由感染的机制,病毒利用宿主的细胞系统进行自我复制,蛋白质-蛋白质相互作用在病毒整个生命周期中发挥着极其重要的作用,研究病毒与宿主蛋白的相互作用对于揭示病毒致病机理、病毒在宿主细胞内引起的信号传导、新型药物的开发与筛选有重要意义。自酵母双杂交技术[17]建立以来,通过不断改进和优化,极大地推动了蛋白质相互作用的研究,酵母双杂交技术已应用于病毒蛋白分析鉴定、细胞周期调控、信号传导、药物靶位点筛选等领域的研究[18-22]。近年来,酵母双杂交技术作为研究蛋白质间相互作用的有力工具,已在水产动物及其病毒相关研究中得以应用[10,23-27]。Cai等[10]在对GCRV非结构蛋白NS80与其NS38、VP4、VP6蛋白相互作用研究中发现,NS80在病毒组装、加工、形成中发挥着重要的作用。GCRV VP5蛋白可能介导病毒粒子进入细胞,VP7可能通过改变蛋白构象协助VP5蛋白穿透宿主细胞膜进入宿主细胞[6],探寻VP5和VP7的相互作用蛋白。深入了解病毒与宿主间的相互作用机制,对于草鱼病毒性出血病的防治和在分子水平上揭示病毒的致病机理具有重要意义。本研究构建CIK酵母Y187双杂交cDNA文库,旨在为研究草鱼呼肠孤病毒与宿主细胞间的相互作用,识别和鉴定相关的宿主细胞因子奠定基础。
1 材料与方法
1.1 材料
1.1.1 CIK细胞 草鱼肾组织细胞系CIK(Ctenopharyngodon idelluskidney,CIK)购买于深圳检验检疫局,培育于添加10%胎牛血清的M199培养基中,28℃温育。
1.1.2 菌株与载体 酵母文库菌株Y187和载体pGADT7-Rec均购自Clontech公司。含重组质粒pGBKT7-VP7的Y2H Gold诱饵菌株和含重组质粒pGBKT7-VP5的Y2H Gold诱饵菌株由本实验构建和保存[28]。
1.2 方法
1.2.1 草鱼细胞系CIK总RNA的提取与mRNA的纯化 在25 mL培养瓶中接种生长状态良好的CIK细胞,待CIK呈单层细胞铺满培养瓶底,移去培养基,用PBS缓冲液漂洗单层细胞2次,移去缓冲液后向培养瓶中加入1 mL Trizol进行裂解,然后按RNA抽提试剂盒(上海生工)说明书进行RNA提取,提取的总RNA用RNase-free H2O溶解并检测质量。
按照mRNA纯化试剂盒(Oligotex mRNA Kit,QIAGEN)操作方法从提取的总RNA中纯化mRNA。
1.2.2 cDNA文库的构建
1.2.2.1 cDNA第一链、第二链的合成和双链cDNA的纯化 利用SMART技术合成全长cDNA,根据酵母双杂交文库构建试剂盒(Make Your Own “Mate & Plate” Library System,Clontech)操作说明合成cDNA第一链和第二链,PCR反应条件为:95℃ 30 s;95℃ 10 s,68℃ 6 min,25个循环;68℃ 5 min。
按照CHROMA SPIN TE-400 Columns说明书纯化双链cDNA。纯化产物用25 μL去离子水溶解并用1%琼脂糖凝胶电泳检测。
1.2.2.2 dscDNA、pGADT7-Rec和carrier DNA共转化酵母菌株Y187 Y187酵母感受态细胞制备和转化按照酵母转化试剂盒(YeastmakerTMYeast Transformation System 2,Clontech)操作说明进行。利用dscDNA和pGADT7-Rec同源重组共转化Y187酵母感受态细胞。
1.2.3 文库克隆子的收获 按酵母转化试剂盒(YeastmakerTMYeast Transformation System 2,Clontech)说明操作,将转化子涂布于150 mm SD/-Leu平板的克隆子菌落长到2-3 mm即可收获克隆子。
1.2.4 文库鉴定 文库收获前随机挑取克隆子菌落PCR检测cDNA插入片段捕获质粒的情况。菌落PCR检测按照全长cDNA合成PCR预混液(MatchmakerTMInsert Check PCR Mix 2,Clontech)操作说明
进行。
1.2.5 互作蛋白的筛选 诱饵菌株在30℃培养3-5 d,分别挑取直径大于2 mm的单菌落接种于50 mL SD/-Trp/Kan液体培养基中,30℃、250-270 r/min 孵育至OD600=0.8,室温1 000×g离心 5 min,弃上清,用 4 mL SD/-Trp/Kan液体培养基重悬沉淀,细胞浓度大于 1.0×108/mL。取1 mL CIK 文库菌液,取出10 μL在SD/-Trp/Kan平板上测定文库滴度(>2.0× 107CFU/mL)后,与上述4 mL诱饵菌液在2 L无菌烧瓶中融合,加 45 mL 含50 μg/mL Kan 的 2×YPDA肉汤,于 30℃慢速震荡(30-50 r/min)培养20-24 h。待观察到“三叶草”接合子形成,1 000×g离心10 min,用10 mL 0.5×YPDA/Kan溶解沉淀,确定总体积。以每平板 200 μL涂QDO/X-α-Gal/AbA平板,30℃培养3-5 d。挑取生长在QDO/X-α-Gal/AbA平板上的蓝色克隆,转移到高筛选率的QDO/X-α-Gal/AbA平板上反复筛选,以确定阳性克隆。
阳性克隆经酵母菌落PCR扩增、鉴定后,送上海生工进行测序,测序后的阳性克隆cDNA序列进行Blast分析。
2 结果
2.1 草鱼CIK细胞总RNA提取及mRNA纯化
提取的草鱼CIK细胞总RNA有3条清晰的条带(图1),分别是28S、18S和5S rRNA。所提取总RNA样品A260/A280=2.004,浓度为2.51 μg/μL,表明所提取的草鱼CIK细胞总RNA质量良好,无DNA、蛋白质污染及小分子污染,28S比18S条带亮。

图1 草鱼CIK细胞总RNA琼脂糖凝胶电泳
利用Oligotex mRNA Kit纯化的草鱼CIK细胞mRNA其大小多集中在250-2 000 bp之间且分布均匀(图2)。

图2 纯化mRNA琼脂糖凝胶电泳
2.2 cDNA第一链合成和双链cDNA纯化结果
以接头引物SMART III-modified oligo反转录合成的cDNA一链产物在100-250 bp之间有一相对集中的条带,但大多数片段主要集中在250-2 000 bp之间,在该区域处有较清晰的丰富带的产生(图3)。
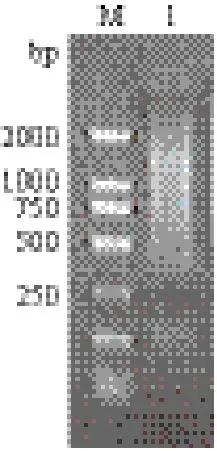
图3 合成的cDNA第一链电泳图
利用CHROMA SPIN TE-400 Columns纯化dsc-DNA,其片段大小在200 bp以上。纯化后的dscDNA大多集中在250-2 500 bp之间,弥散成明显的瀑布状(图4)。

图4 dscDNA纯化电泳图
2.3 文库的构建及鉴定
100倍稀释度时,平板克隆子个数为480个,计算得出转化率为1.6×105,文库容量为2.4×106。
文库收获后,经血球计数板计数,其滴度为7×107CFU/mL,满足试剂盒建库要求。
酵母菌落PCR检测结果显示文库质粒插入片段的大小均在250 bp以上,大多分布在500-2 000 bp(其中1 000-2 000 bp的片段占46%)之间(图5),挑取总克隆的50%,2 000 bp以上只出现一个条带,可能与挑取菌落的随机性有关,cDNA文库的重组率为98%。

图5 酵母菌落PCR电泳图
2.4 GCRV VP7和VP5相互作用蛋白的筛选
构建的pGBKT7-VP7和pGBKT7-VP5诱饵载体转入酵母Y2H Gold后,经检测无自激活性和对酵母的毒性现象。诱饵菌株与文库菌液混合后可观察到“三叶草”接合子形成,在QDO/X-α-Gal/AbA培养基上生长,再用高筛选率的QDO/X-α-Gal/AbA平板反复筛选时,获得VP7互作蛋白的阳性菌落(图6箭头所示),并没有筛选到VP5互作蛋白的阳性菌落。
将VP7阳性菌落进行PCR扩增和测序,获得阳性菌落的cDNA序列(图7)。对阳性克隆序列进行Blastn比对分析发现,在数据库中具有较大片段的同源序列,与斑点叉尾鮰40S核糖体蛋白S20具有89%的同源性,同源性最高;与其他多数生物核糖体蛋白S20的同源性均在83%以上。

图6 QDO/ X-α-Gal/AbA培养基上与VP7相互作用蛋白筛选的阳性菌落

图7 阳性菌落序列
3 讨论
全长cDNA文库不仅包含完整的开放阅读框,还需含有5'和3'非编码区序列。RNA的纯度和完整性决定着cDNA文库的构建质量即序列信息的完整性和多态性[29]。真核生物总RNA的完整性通常用28S、18S rRNA的质量来衡量,28S与18S的强度一般为1.5-2.5∶1。本研究以草鱼肾组织细胞系CIK为试验材料提取总RNA,琼脂糖凝胶电泳显示28S rRNA比18S rRNA的亮度稍高,并且条带清晰,提取的总RNA质量较好,可以进一步用于mRNA的纯化。
一般认为非哺乳动物的mRNA大多分布在0.5-3.0 kb之间,为防止出现利用总RNA构建文库产生的小片段冗余现象,本研究进行了mRNA的纯化。纯化的草鱼CIK细胞mRNA的条带大小集中在250-2 000 bp之间且分布均匀,250-500 bp之间的片段条带也相对较亮和较集中,可能是由于mRNA纯化过程中mRNA发生降解和断裂产生的。有些研究人员认为mRNA不需要纯化,纯化mRNA反而更容易造成降解[30]。本研究纯化的mRNA符合构建文库的一般规律和本试验要求,可以用于全长cDNA文库的构建。
构建高质量的全长cDNA文库是非常关键的,文库中如何区分完整的cDNA和变短的cDNA片段尚无较好的方法。由于小片段的cDNA扩增时间
短,即使小片段数量很少,在足够的循环次数下仍能大大超过全长cDNA数量,产生大量冗余。本研究利用Clontech公司Make Your Own “Mate&PlateTM”Library试剂盒合成全长cDNA。该试剂盒采用SMART(Switching mechanism at 5' end of RNA transcript)技术即模板转换技术合成全长cDNA,且试剂盒所带cDNA双链引物的5'端都携带有和文库质粒pGADT7-Rec相同的同源重组片段,cDNA全长合成后不需经过双酶切就可通过同源重组和文库质粒相连接。序列信息不完整的mRNA逆转录时由于5'端序列信息的缺失,MMLV便不会在cDNA第一链3'末端添加Oligo(C),因此不完全的cDNA第一链便不会继续合成第二链cDNA,最终获得的cDNA都是全长cDNA。同时采用YeastmakerTM酵母转化系统2,酵母转化效率高,且易于筛选转化子。本研究成功构建了草鱼细胞系CIK酵母双杂交cDNA文库,且cDNA较完整,为研究草鱼呼肠孤病毒与宿主细胞间的互作机制奠定了基础。
构建cDNA文库是研究蛋白互作的重要途径,鉴于细胞系CIK在GCRV研究方面的广泛应用[13-16],采用SMART技术成功构建了CIK的酵母Y187 AD融合cDNA文库。cDNA文库质量评价主要是检测文库的代表性和序列完整性,本研究所构建文库的原始库容量为2.4×106CFU,滴度为7×107CFU/mL,重组率为98%,达到了CLONTECH公司SMART文库构建指标(原始文库的库容大于1×106CFU,滴度大于1×107CFU/mL,重组率大于85%),此文库具有较好的cDNA片段多态性和完整性,可用于进一步的文库筛选。将GCRV VP7和VP5作为诱饵蛋白进行杂交试验,筛选到VP7的相互作用蛋白,但未筛选到VP5的相互作用蛋白(利用生物信息学技术分析预测VP5无跨膜结构域,确切的原因有待试验验证)。
VP7相互作用蛋白的阳性克隆序列Blast比对结果可以断定得到的阳性克隆序列结果相对准确,此序列经生物信息学软件分析显示无完整的阅读框,可能是由于mRNA在反转录过程中发生降解,故认为此阳性克隆序列为草鱼肾组织细胞系40S核糖体亚基S20蛋白基因的一段序列。与阳性克隆序列同源的核糖体蛋白S20除了作为组成40S核糖体亚基的组分外,还可以结合到自身mRNA特定位点抑制其自身基因翻译表达[31]。根据VP7与阳性克隆序列间的相互作用推测,VP7可能通过与草鱼核糖体蛋白S20的结合抑制了S20调节自身翻译表达的功能,从而使S20出现高表达量异常。由于未获得完整的阳性克隆序列开放阅读框,对于阳性克隆和VP7是如何发生作用的以及发生作用的蛋白质结构域无法作出预测。由于阳性克隆序列编码蛋白属于胞内蛋白,因此该结果尚不能验证目前推测的GCRV VP7蛋白的功能。病毒蛋白在宿主细胞内如何利用宿主的翻译系统;在病毒粒子的组装、释放过程中,病毒和宿主间究竟发生了怎样的相互作用,其具体机制究竟是怎样的都不清楚,尚需要进一步的研究。对GCRV VP7和VP5的互作蛋白和作用机制进行研究,将有助于揭示GCRV的感染机理。CIK酵母双杂交cDNA文库的构建对于GCRV与宿主细胞间的互作机制研究、揭示病毒致病机理及其在宿主细胞内引起的信号传导和新型药物的开发和筛选具有重要的意义。
4 结论
本研究成功构建了草鱼细胞系CIK双杂交cDNA文库,并应用此文库筛选到GCRV VP7互作蛋白的阳性克隆,推测VP7的互作蛋白可能是草鱼肾组织细胞系40S核糖体亚基S20蛋白。
[1] 陈燕新, 江育林.草鱼出血病毒形态结构及其理化特性研究[J].科学通报, 1983, 18:1138-1140.
[2] Reinisch KM, Nibert ML, Harrison SC. Structure of the reovirus core at 3.6 A resolution[J]. Nature, 2000, 404(6781):960-967.
[3] Grimes JM, Burroughs JN, Gouet P, et al. The atomic structure of the bluetongue virus core[J]. Nature, 1998, 395(6701):470-478.
[4] 肖波.草鱼呼肠孤病毒及其免疫防治研究进展[J].鲁东大学学报, 2010, 26(1):48-53.
[5] 何福林, 肖克宇, 龚泽修.草鱼出血病病毒血凝作用及影响因素[J].湖南农业大学学报:自然科学版, 2002, 28(1):47-50.
[6] Seng EK, Fang Q, Lam TJ, et al. Development of a rapid, sensitive and specific diagnostic assay for fish Aquareovirus based on RT-
PCR[J]. Virological Methods, 2004, 118(2):111-122.
[7] 方勤, 肖调义, 李旅, 等.四株草鱼呼肠孤病毒毒株的细胞感染特性比较研究[J].中国病毒学, 2002, 17(2):182-184.
[8] Cheng LP, Fang Q, Shah S, et al. Subnanometer-resolution structures of the grass carp reovirus core and virion[J]. Molecular Biology, 2008, 382(1):213-222.
[9] Danthi P, Coffey CM, Parker JS, et al. Independent regulation of reovirus membrane penetration and apoptosis by the mu1 phi domain[J]. Plos Pathology, 2008, 4(12):e1000248:1-14.
[10] Cai L, Sun XY, Shao L, et al. Functional investigation of grass carp reovirus nonstructural protein NS80[J]. Virology Journal, 2011, 8:168.
[11] 左文功, 钱华鑫, 许映芳, 等.草鱼肾组织细胞系CIK的建立及其生物学特性[J].水产学报, 1986, 1:11-17.
[12] 方勤, 艾桃山, 邹桂平, 等.鳜鱼病毒病原及细胞感染特性的研究[J].中国病毒学, 2000, 15(3):297-301.
[13] 杨琳, 汪亚平, 廖兰杰, 等.发夹RNA在稀有鲫胚胎中的表达[J].水生生物学报, 2010, 34(5):1031-1036.
[14] Zhang LL, Luo Q, Fang Q, et al. An improved RT-PCR assay for rapid and sensitive detection of grass carp reovirus[J]. J Virol Methods, 2010, 169(1):28-33.
[15] Ye X, Tian YY, Deng GC, et al. Complete genomic sequence of a reovirus isolated from grass carp in China[J]. Virus Res, 2012, 163(1):275-283.
[16] 王土, 许丹, 吕立群.应用dsRNA测序技术检测草鱼呼肠孤病毒的混合感染[J].上海海洋大学学报, 2012, 21(5):756-762.
[17] Fields S, Song O. A noval genetic system to detect protein-protein interactions[J]. Nature, 1989, 340(6646):245-246.
[18] Reis AL, McCauley JW. The Influenza virus protein PB1-F2 interacts with IKKβ and modulates NF-κB signalling[J]. PLoS One, 2013, 8(5):e63852.
[19] Seo JK, Kwon SJ, Rao AL. Molecular dissection of Flock house virus protein B2 reveals that electrostatic interactions between N-terminal domains of B2 monomers are critical for dimerization[J].Virology, 2012, 432(2):296-305.
[20] Dietrich D, Seiler F, Essmann F, et al. Identification of the kinesin KifC3 as a new player for positioning of peroxisomes and other organelles in mammalian cells[J]. Biochim Biophys Acta, 2013, 1833(12):3013-3024.
[21] Wei JD, Kim JY, Kim AK, et al. RanBPM protein acts as a negative regulator of BLT2 receptor to attenuate BLT2-mediated cell motility[J]. J Biol Chem, 2013, 288(37):26753-26763.
[22] von Eichborn J, Dunkel M, Gohlke BO, et al. SynSysNet:integration of experimental data on synaptic protein-protein interactions with drug-target relations[J]. Nucleic Acids Res, 2013, 41(Database issue):D834-840.
[23] Chen J, Lu XJ, Yang HY, et al. An interaction between leukocyte cell-derived chemotaxin 2 and transferrin of ayu,Plecoglossus altivelis[J]. Fish Shellfish Immunol, 2009, 26(3):536-542.
[24] 徐坤, 王亮亮, 张婷婷, 等.大眼狮鲈鱼皮肤肿瘤病毒(WDSV)逆转录病毒周期蛋白(OrfA)相互作用蛋白的酵母双杂交筛选[J].中国生物化学与分子生物学报, 2012, 28(3):267-275.
[25] Otta SK. Host and virus protein interaction studies in understanding shrimp virus gene function[J]. Indian J Virol, 2012, 23(2):184-190.
[26] Sangsuriya P, Huang JY, Chu YF, et al. Construction and application of a protein interaction map for white spot syndrome virus(WSSV)[J]. Mol Cell Proteomics, 2014, 13:269-282.
[27] Yang CG, Wang XL, Zhang B, et al. Screening and analysis of PoAkirin1 and two related genes in response to immunological stimulants in the Japanese flounder(Paralichthys olivaceus)[J]. BMC Mol Biol, 2013, 14(1):10.
[28] 谢吉国, 闫秀英, 简纪常, 吴灶和.草鱼呼肠孤病毒vp5基因诱饵重组载体的构建、转化与自激活作用检测[J].广东海洋大学学报, 2012, 32(3):42-48.
[29] Zhu YY, Machleder EM, Chenchik A, et al. Reverse transcriptase template switching:A SMARTTMapproach for full-length cDNA library construction[J]. Biotechniques, 2001, 30(4):892-897.
[30] 荆晶, 刘铜, 陈捷.玉米弯孢叶斑病菌酵母双杂交cDNA文库的构建及评价[J].植物病理学报, 2012, 42(1):93-96.
[31]Parsons GD, Mackie GA. Expression of the gene for ribosomal protein S20:Effects of gene dosage[J]. Journal of Bacteriology, 1983, 154(1):152-160.
(责任编辑 马鑫)
Construction and Preliminary Application of the Yeast Two-hybrid cDNA Library from Grass Carp CIK Cells
Yan Xiuying1Xie Jiguo1Li Jie1Ding Yu1Wu Zaohe2Lu Yishan1Jian Jichang1
(1. Guangdong Key Laboratory of Pathogenic Biology and Epidemiology for Aquatic Economic Animals,Fisheries College of Guangdong Ocean University,Zhanjiang 524088;2. Zhongkai University of Agriculture and Engineering,Guangzhou 510225)
In order to explore the interactions between grass carp reovirus and the host cell proteins, the yeast two-hybrid cDNA library from grass carp CIK(Ctenopharyngodon idellus kidney) was constructed by SMART technology. Total RNA of the CIK cells was extracted and mRNA was purified. Then, mRNA as the template, the first strand of cDNA was synthesized by reverse transcription. The double-stranded cDNA was amplified through the long-distance PCR with the DNA polymerase. The cDNA library from grass carp CIK cells was constructed in the yeast strain 187, using SMART technology and the homologous recombination method. After testing, the conversion rate and the capacity of the original library was 1.6×105and 2.4×106, respectively. The length of the inserted double-stranded cDNA fragments were 250-2 000 bp. The titer of the library was 7×107CFU/mL and the recombination rate was 98%. This library had the well polymorphism and the integrity of the cDNA fragments. Using the constructed yeast two-hybrid from the CIK cells, the VP7 and VP5 protein in grass carp reovirus were as the baits for the screening experiment. The positive colonies of the interacting proteins of VP7 had been acquired, without the positive colony of the interacting protein of VP5. The construction of the yeast two-hybrid cDNA library from the grass carp CIK cells provided an important research tool for the study on the interaction mechanism of grass carp reovirus with the host cell.
Ctenopharyngodon idellus kidney(CIK) Yeast two-hybrid cDNA library Protein interaction
2013-11-26
国家“973”计划项目(2009CB118704)
闫秀英,女,博士,讲师,研究方向:水产动物病害防控;E-mail:yanxiuying1201@126.com
简纪常,男,博士,教授,研究方向:水产动物免疫学及病害控制;E-mail:jianjc@gmail.com
