菲律宾蛤仔不同品系斑马蛤的选择反应及现实遗传力*
2014-03-08闫喜武张跃环张国范
刘 振 闫喜武① 张跃环 杨 鹏 姚 托 杨 凤 张国范
(1.大连海洋大学水产与生命学院 辽宁省贝类良种繁育工程技术研究中心 大连 116023;2.中国科学院南海海洋研究所热带海洋生物资源与生态重点实验室 广州 510301;3.中国科学院海洋研究所 海洋生物技术研发中心 青岛 266071)
选择育种是动植物遗传改良中最经典、最有效的方法之一。对单性状的选择方法中,个体选择是最为常用的。个体选择也称大群选择或混群选择,只需根据个体本身表型值进行选择,简单易行,而且选择效果比较明显。它已经广泛应用于贝类的遗传改良,用于增加产量和提高存活率。近年来,国内外学者已经对多种贝类开展了选择育种研究,如长牡蛎(Liet al,2011)、海湾扇贝(Zhenget al,2004,2006)、虾夷扇贝(Lianget al,2010)、华贵栉孔扇贝(Liuet al,2012)、马氏珠母贝(邓岳文等,2008;Heet al,2008;Denget al,2009)、珠母贝(Wanget al,2011)、硬壳蛤(Hadleyet al,1991;Heffernanet al,1991)、菲律宾蛤仔(闫喜武等,2010,2011)等。
菲律宾蛤仔肉味鲜美,营养丰富,是我国的四大养殖贝类之一。目前,我国年产蛤仔在 200万吨以上,是我国单产最高的一种贝类(张国范等,2010)。蛤仔壳色存在复杂的多态现象,闫喜武等(2005,2010)以壳色为标记,通过定向选育获得了外观漂亮、抗性较强的斑马蛤品系。在此基础上,张跃环等(2008,2012)以斑马蛤为核心种质品系,通过与生长快的珍珠白品系杂交,获得了生长快、抗性强的白斑马品系;同时与抗性较强玛瑙黑品系杂交,获得了抗性极强的黑斑马品系。为了进一步提高斑马蛤各品系的生长性能,我们以三个品系的蛤仔作为基础群体,通过歧化选择,研究了它们的选择反应和现实遗传力。研究不仅具有重要的应用价值,而且具有重要的理论价值。一方面,通过选择所培育出的快速生长品系可直接应用于蛤仔的生产实践,另一方面,研究结果也可进一步充实贝类养殖遗传学理论。
1 材料与方法
1.1 亲贝来源及促熟
以斑马蛤、玛瑙黑、珍珠白品系为材料,通过群体自繁和群体杂交的方法获得了斑马蛤(Z,Zebra)、黑斑马(AbZ,Agate black Zebra)和白斑马(WZ,White Zebra)群体,每个群体经过4—5代选择,形成了不同斑马蛤品系。采用底播的方式放置在大连海量水产食品有限公司的虾池中进行生态促熟(闫喜武等,2008;霍忠明等,2010)。在相同的饲养条件下,于2011年9月下旬,3个斑马蛤品系性腺同步成熟。
1.2 实验设计与处理
从每个群体中随机选取 300个个体作为基础群体,用游标卡尺进行测量(精确至 0.02mm),绘制壳长频数分布直方图(图1)。按10%的留种率,选择最大的30个个体作为选择组(SG),在此之前从群体中随机抽取30个个体作为对照组(CG)。表1是三个品系的选择截点和选择强度。由于贝类的生长受环境的影响变化很大,故采用设置对照组的方法,消除环境因子对计算遗传力的影响(Zhenget al,2004,2006)。

表1 三个品系选择组和对照组亲本的大小、截头选择的截点和选择强度Tab.1 The shell length and its cut-off point,and selection intensity of three groups of zebra clam strains

图1 蛤仔斑马蛤三个群体的大小分布及被截头选择的亲本(黑色部分)Fig.1 Size distribution of three zebra strains and the parents selected for truncation selection(black parts)
1.3 受精和孵化
将处于临产状态的亲贝阴干 12h后,分别置于80L白桶中,加入新鲜海水待产,大约4h后亲贝开始排精产卵。产卵过程持续 1—2h,镜检受精情况后开始洗卵,除掉杂质和多余的精子。将受精卵分桶孵化,约24h后发育为D形幼虫。整个过程中,各实验组严格分离,防止混苗。期间水温 21.5—22.3°C,盐度22—24,pH 7.85—7.93。
1.4 幼虫培育、稚贝育成及越冬
幼虫培育在 80L的白色塑料桶中,密度为3—4ind./mL,各实验组分别设置3个重复。每2d换一次水,换水量100%。饵料每天投喂两次,前期为金藻,后期为小球藻,投饵量视幼虫摄食情况而定。为防止不同实验组幼虫混杂,换水网袋单独使用。幼虫培 育 阶 段 ,水 温 18.5—22.0°C,盐 度 22—24,pH7.88—8.52。在幼虫期,定期对密度进行调整,使各组之间密度保持一致。
稚贝培育阶段,随着稚贝的生长,投饵量视稚贝摄食情况而定,其它同幼虫期管理。由于进入深秋季节,水温逐渐降低,当稚贝壳长≥400μm 时,将其装入80目网袋,放到60m3的水泥池中进行室内越冬。期间水温为生态池中自然水温(-1.0—15.2°C),盐度26—30,pH7.80—8.28。期间定期更换网袋,调整密度,使其保持一致。
1.5 幼贝育成
从2012年 5月开始,将稚贝从室内转移至室外生态池中进行中间育成。期间水温 15.6—30.6°C,盐度22—26,pH7.64—8.56。期间定期更换不同目数的网袋(20—60目),消除网目对幼贝的影响,同时不断降低密度,但要保持各组密度一致。
1.6 测定指标
壳长小于 300μm的幼虫和稚贝在显微镜下用目微尺(100×)测量,壳长大于300μm小于3.0mm的稚贝测量在显微镜下用目微尺(25×)进行,壳长大于3.0mm的幼贝和亲贝用游标卡尺测量。每次测量设3个重复,每个重复随机测量30个个体。
1.7 数据处理
参照Zheng使用的方法(Zhenget al,2004,2006),将不同地理群体菲律宾蛤仔的选择反应(R)和现实遗传力()用以下两个公式计算:


公式(1)和(2)中:XSG和XCG分别是选择组(SG)和对照组(CG)个体的平均壳长,SCG是对照组(CG)的标准差,i是选择强度。
用 SPSS19.0统计软件对数据进行分析处理,不同实验组间数据的比较采用单因素方差分析方法,差异显著性设置为P<0.05。
2 结果
2.1 各品系壳长
对于斑马蛤品系而言,除 3日龄以外,上选组壳长均显著大于对照组(P<0.05);对于黑斑马而言,上选组壳长仅仅在6、240、300日龄显著大于对照组(P<0.05),其它时期与对照组无显著差异(P>0.05);对于白斑马而言,上选组壳长一直显著大于对照组(P<0.05)。说明各品系上选效果是不同的,存在着品系间遗传差异。因而对斑马蛤和白斑马蛤的选择是比较有效的(表2)。

表2 不同品系斑马蛤各实验组壳长Tab.2 Shell length of every experimental groups among different strains of zebra clam
2.2 选择反应和现实遗传力
三个品系蛤仔在不同时期的选择反应和现实遗传力估计值见表3。就选择反应而言,在幼虫期及稚贝期,白斑马>斑马蛤>黑斑马;养成期,白斑马(0.731)≈斑马蛤(0.727)>黑斑马(0.549)。就现实遗传力而言,在幼虫期及稚贝期,白斑马>斑马蛤>黑斑马;养成期,白斑马(0.417)≈斑马蛤(0.410)>黑斑马(0.313)。就总体而言,斑马蛤、黑斑马及白斑马在整个时期选择反应的平均值为 0.655、0.525、0.782,3个品系的平均值为 0.654;白斑马的现实遗传力最高(0.445),黑斑马的最低(0.300),斑马蛤居中(0.372),3个品系的平均值为0.373(表4)。
表3 各品系斑马蛤不同阶段的选择反应和现实遗传力Tab.3 Response to selection(R)and realized heritability()of each strains of zebra clam during different stage
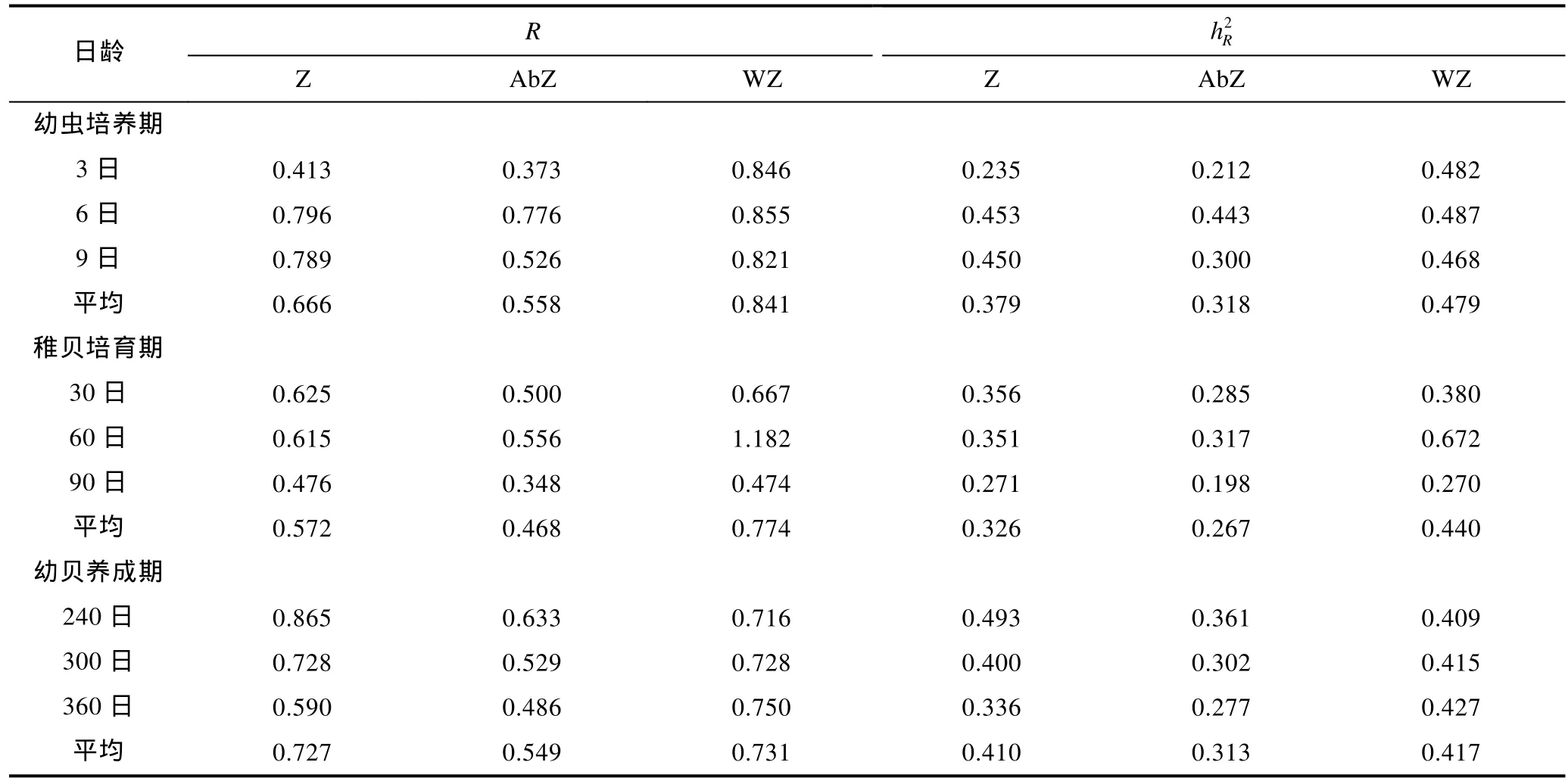
表3 各品系斑马蛤不同阶段的选择反应和现实遗传力Tab.3 Response to selection(R)and realized heritability()of each strains of zebra clam during different stage
R:选择反应;:现实遗传力;Z:斑马蛤;AbZ:黑斑马;WZ:白斑马
日龄R2 hR Z AbZ WZ Z AbZ WZ幼虫培养期3日 0.413 0.373 0.846 0.235 0.212 0.482 6日 0.796 0.776 0.855 0.453 0.443 0.487 9日 0.789 0.526 0.821 0.450 0.300 0.468平均 0.666 0.558 0.841 0.379 0.318 0.479稚贝培育期30日 0.625 0.500 0.667 0.356 0.285 0.380 60日 0.615 0.556 1.182 0.351 0.317 0.672 90日 0.476 0.348 0.474 0.271 0.198 0.270平均 0.572 0.468 0.774 0.326 0.267 0.440幼贝养成期240日 0.865 0.633 0.716 0.493 0.361 0.409 300日 0.728 0.529 0.728 0.400 0.302 0.415 360日 0.590 0.486 0.750 0.336 0.277 0.427平均 0.727 0.549 0.731 0.410 0.313 0.417
表4 各品系斑马蛤总体水平的选择反应和现实遗传力Tab.4 Thetotal response to selection(R)and realized heritability()of each strains of zebra clam

表4 各品系斑马蛤总体水平的选择反应和现实遗传力Tab.4 Thetotal response to selection(R)and realized heritability()of each strains of zebra clam
R:选择反应; :现实遗传力;Z:斑马蛤;AbZ:黑斑马;WZ:白斑马
类别 Z AbZ WZ Mean R 0.655±0.155 0.525±0.116 0.782±0.170 0.654±0.179 h 0.372±0.042 0.300±0.028 0.445±0.031 0.373±0.070 2 R
3 讨论
在选择育种过程中,采用理想的选择群体是非常重要的。群体存在较高的遗传变异水平,是群体选育的关键前提(马爱军等,2012)。邓岳文等通过建立遗传变异程度较高的马氏珠母贝群体,得到选系 F2早期的选择反应为0.63—0.89(邓岳文等,2008)。郑怀平等选用海湾扇贝北方亚种两个不同养殖群体的亲本构建了一个基础群体,这一群体可以被看作是一个基因库,结果表明选择不仅显著地提高了生长速度和存活率,而且缩短了附着变态时间,与对照组相比,选择组具有更大的潜在优势(郑怀平,2005)。在本研究中,三个品系在整个阶段选择反应的平均值与海湾扇贝 1998—1999年引种的野生群体大小比较接近,但略小于早期的马氏珠母贝及不同地理群体的菲律宾蛤仔(Zhenget al,2004;邓岳文等,2008;闫喜武等,2010)。三个斑马蛤品系的选择反应不同,且存在一定的差异,但三个群体的选择反应还是处于较高的水平,其中,以白斑马群体的选择反应最大,斑马群体次之,黑斑马群体的选择反应最小。本次实验所用斑马蛤源自于莆田群体,其比例仅为 0.25%,且经过了几代的定向纯化而成(闫喜武等,2005);而白斑马群体最初是由斑马蛤和白蛤通过群体杂交所得,后来经过连续几代的定向选育出来的,其基因型相对丰富,所以其选择反应较高也是合理的。然而用同样方法获得的黑斑马群体的选择反应却明显低于白斑马群体。一方面,斑马蛤、玛瑙黑的生长速度在蛤仔中均较慢,通过杂交得到的黑斑马蛤并未表现出杂种优势(苏家齐等,2013),无论是选择组还是对照组在生长速度上基本不存在显著差异,黑斑马亲本的选择组壳长也是最小的,说明黑斑马群体生长性状的遗传变异程度较低,导致其选择反应相对较小。另一方面,选择反应是加性遗传方差与选择强度共同作用的结果(Falconer,1996),黑斑马的选择强度在三个品系中最小,可能也是导致选择反应较小的缘故。此外,在幼贝养成期,无论是选择组还是对照组,斑马蛤和白斑马品系都是可以稳定遗传的,但是黑斑马出现了个别壳色分离现象,这可能对其选择反应有一定程度的影响。
一般而言,某一性状的遗传力越大,其加性遗传效应可能就越明显(Toroet al,1995;Falconer,1996;张国范等,2009),选择也越有效,而现实遗传力与选择反应是两个不同数量遗传学的概念,但在某种程度上二者具有一定的相关性(Falconer,1996;Crenshawet al,1991)。斑马蛤、黑斑马和白斑马品系的现实遗传力估计值在0.30—0.45之间。与其它双壳类关于现实遗传力的报道(Newkirket al,1983;Wada,1986;Hadleyet al,1991;Ibarraet al,1999)相比,斑马蛤和黑斑马品系现实遗传力的估测结果只是中等的,但白斑马品系的估测结果相对较高。总体来说,斑马蛤与白斑马品系具有一定的选择潜力,尤其是白斑马品系。由此可见,通过选择对其进行遗传改良,是提高产量的一种可行的方法。
邓岳文,符 韶,杜晓东,2008.马氏珠母贝选系F2早期选择反应和现实遗传力估计.广东海洋大学学报,28(4):26—29
霍忠明,闫喜武,张跃环等,2010.菲律宾蛤仔(Ruditapes philippinarum)大连群体家系建立及生长比较.海洋与湖沼,41(3):334—340
马爱军,王新安,黄智慧等,2012.大菱鲆(Scophthalmus maximus)家系选育 F2早期选择反应和现实遗传力估计.海洋与湖沼,43(1):57—61
苏家齐,闫喜武,张跃环等,2013.两种三元杂交菲律宾蛤仔壳色品系间的聚合杂交.海洋与湖沼,44(2):513—518
闫喜武,2005.菲律宾蛤仔养殖生物学、养殖技术和品种选育.青岛:中国科学院海洋研究所博士学位论文
闫喜武,张跃环,孙焕强等,2011.菲律宾蛤仔海洋橙品系两个世代的杂交与近交效应.海洋与湖沼,42(2):309—316
闫喜武,张跃环,霍忠明等,2008.常见滩涂贝类亲本生态促熟工艺.专利号:ZL200810013304.6
闫喜武,张国范,杨 凤等,2005.菲律宾蛤仔莆田群体两个壳色品系生长发育的比较.大连水产学院学报,20(4):266—269
闫喜武,张跃环,霍忠明等,2010.不同地理群体菲律宾蛤仔的选择反应及现实遗传力.水产学报,34(5):705—710
张跃环,2008.菲律宾蛤仔壳色、壳型的品系选育及遗传机制研究.大连:大连水产学院硕士论文
张跃环,闫喜武,李少文等,2012.菲律宾蛤仔(Ruditapes philippinarum)紫壳内面和白黄壳内面品系的三元杂交.海洋与湖沼,43(3):300—307
张国范,郑怀平,2009.海湾扇贝养殖遗传学.北京:科学出版社,52—73
张国范,闫喜武,2010.蛤仔养殖学.北京:科学出版社,11—13
郑怀平,2005.海湾扇贝两个养殖群体数量性状及壳色遗传研究.青岛:中国科学院海洋研究所博士论文
Crenshaw J J W,Heffernan P B,Walker R L,1991.Heritability of growth rate in the southern bay scallop,Argopecten irradians concentricus(Say,1822).Journal of Shellfish Research,10(1):55—63
Deng Y W,Fu S,Du X D,2009.Realized heritability and genetic gain estimates of larval shell length in the Chinese Pearl OysterPinctada martensiiat three different salinities.North American Journal of Aquaculture,71(4):302—306
Falconer D S,1996.Introduction to quantitative genetics.2nd edn.Longman Press,London,UK:247—306
Hadley N H,Dillon J R T,Manzi J J,1991.Realized heritability of growth rate in the hard clamMercenaria.mercenaria.Aquaculture,93(2):109—119
Heffernan P B,Walker R L,Crenshaw J J W,1991.Negative larval response to selection for increased growth rate in the northern quahog,Mercenaria mercenaria.Journal of Shellfish Research,1:199—202
He M X,Guan Y Y,Yuan T,2008.Realized heritability and response to selection for shell height in the pearl oysterPinctada fucata(Gould).Aquaculture Research,39(8):801—805
Ibarra A M,Ramirez J L,Ruiz C Aet al,1999.Realized heritabilities and genetic correlation after dual selection for total weight and shell width in catarina scallop(Argopecten circularis).Aquaculture,175(3):227—241
Liu J Y,Liu Z G,2012.Response to selection for shell length in the noble scallop,Chlamys nobilis(Reeve).African Journal of Biotechnology,11(36):8794—8799
Liang J,Zhang G F,Zheng H P,2010.Divergent selection and realized heritability for growth in the Japanese scallop,Patinopecten yessoensis Jay.Aquaculture Research,41(9):1315—1321
Li Q,Wang Q Z,Liu SKet al,2011.Selection response and realized heritability for growth in three stocks of the Pacific oysterCrassostrea gigas.Journal of Shellfish Research,77(4):643—648
Newkirk G F,Haley L E,1983.Progress in selection forgrowth rate in the European oysterOstre aedulis.Marine ecology progress series,10(1):77—79
Toro J E,Sanhueza M A,Winter JE,et al,1995.Selection response and heritability estimates for growth in the Chilean OysterOstrea chilensis(Philippi,1845).Journal of Shellfish Research,14:87—92
Wada K T,1986.Genetic selection for shell traits in the Japanese pearl oyster,Pinctada fucata martensii.Aquaculture,57(1—4):171—176
Wang Q H,Deng Y W,Du X D,2011.Realized heritability and genetic gains of three generation for superior growth in the pearl oysterPinctada martensii.Acta Ecologica Sinica,31(2):108—111
Zheng H P,Zhang G F,Liu Xet al,2004.Different responses to selection in two stocks of the bay scallop,Argopecten irradians irradiansLamarck(1819).Journal of Experimental Marine Biology and Ecology,313(2):213—223
Zheng H P,Zhang G F,Liu Xet al,2006.Sustained response to selection in an introduced population of the hermaphroditic bay scallopArgopecten irradians irradiansLamarck(1819).Aquaculture,255(1—4):579—585
