东海南部海域头足类群落结构特征及其与环境关系*
2014-03-08朱文斌薛利建卢占晖徐汉祥徐开达
朱文斌 薛利建 卢占晖 徐汉祥 徐开达①
(1.中国海洋大学水产学院 青岛 266003;2.浙江省海洋水产研究所 浙江省海洋渔业资源可持续利用技术研究重点实验室农业部重点渔场渔业资源科学观测实验站 舟山 316004)
头足类是一种生命周期短、生长迅速、资源恢复力强的海洋动物。东海区头足类资源丰富,长期以来一直是海洋捕捞的重要对象之一(覃涛等,2011)。历史上曼氏无针乌贼(Sepiella maindroni)曾是舟山渔场著名的“四大鱼产”之一,东海区的最高年产量曾达到7×104t,但由于过度捕捞,从20世纪80年代开始,资源量急剧下降,至今已不能形成鱼汛(宋海棠等,2009)。与此同时,随着20世纪90年代发展了单拖作业,新的头足类资源诸如太平洋褶柔鱼(Todarodes pacificus)、金乌贼(Sepia esculenta)等逐渐被开发和利用(郑元甲等,1999)。
对于中国沿海头足类生态学方面的研究,以往主要集中在种类组成和数量分布方面(董正之,1992;宋海棠等,1996,2008;徐开达等,2008;黄梓荣,2008;俞存根等,2009),也有少量关于其资源量评估、群聚特征以及近岸海域群落结构的研究(丁天明等,2001;李圣法等,2006;陈强等,2012)。对于头足类群落结构、动态变化以及与环境之间的关系等方面的研究,至今尚未见报道。本研究以2008—2009年进行的“东海区主要渔场重要渔业资源调查与评估”课题开展的周年 4个季度的桁杆拖虾网调查所获得的头足类资源资料,对东海南部海域头足类的种类组成、优势种、群落的物种多样性以及群落与环境因子的关系等方面进行了初步探讨,旨在为保护东海南部头足类群落,修复生态环境,更好地利用头足类资源以及渔业主管部门制定管理政策提供科学依据。
1 材料与方法
1.1 数据来源及处理
本研究所用数据来自2008年5月(春)、8月(夏)、11月(秋)和2009年2月(冬)进行的“东海区主要渔场重要渔业资源调查”课题所获得的头足类调查资料。调查范围为26°00′—28°00′N,126°00′E以西海域,共设置30个站位,站位分布见图1。调查船主机功率255kW,调查网具为桁杆拖虾网,桁杆长36m,囊网10支,网囊网目25mm。采用走航式24h不间断调查,平均拖速2kn,每站拖网时间均标准化为1h,采用温盐深测量仪同步调查各站位的水深、表层水温、表层盐度、底层水温和底层盐度等5项环境指标,调查采样及测定均按《海洋调查规范——海洋生物调查》(国家技术监督局,1991)的有关标准进行。各站点渔获物样品随机采样1箱(20kg),渔获物中头足类种类尽可能鉴定到最小分类单元,并记录每种的重量和尾数。
1.2 分析方法
1.2.1 优势种 群落优势种采用 Pinkas等(1971)的相对重要性指数(index of relative importance,IRI)进行划分。其公式为:

式中,N%为某一种类的尾数占总尾数的百分比;W%为某一种类的重量占总重量的分比;F为某一种类出现的站数占调查总站数的百分比。本文中将相对重要性指数(IRI)大于 1000的种类定为优势种(陈强等,2012)。
1.2.2 群落多样性指数 由于不同种类及同种类个体间差异很大,Wilhm等(1971)提出用生物量表示的多样性更接近种类间能量的分布,因此文中根据生物量(单位时间内所捕获的各头足类种类的重量)计算头足类群落物种多样性(Ludwiget al,1988)。
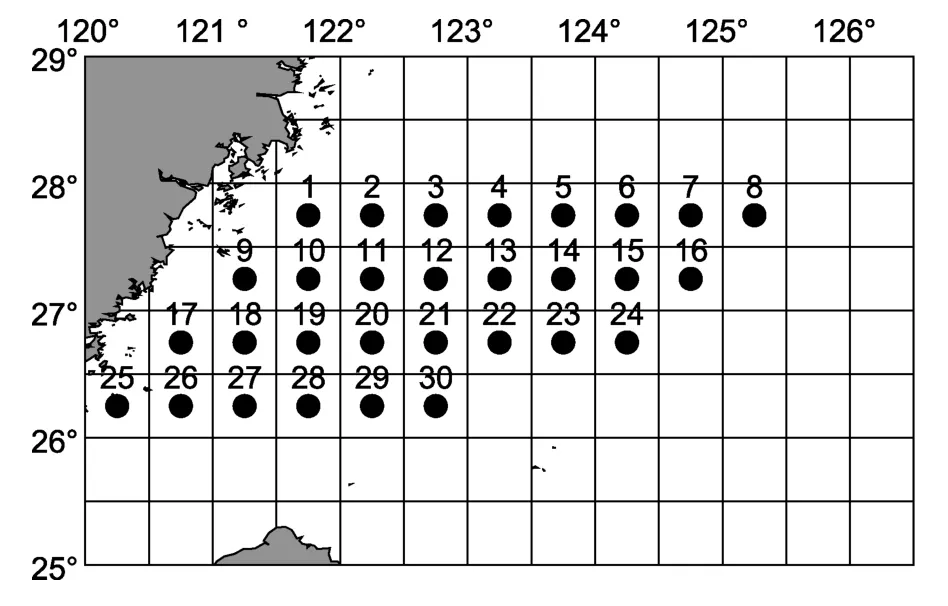
图1 东海南部调查站位分布图(●)Fig.1 Location of sampling stations(●)in the southern East China Sea
本文头足类群落多样性分析运用下列公式(Margalef,1957;Pielou,1975;马克平,1994):
种类丰富度指数D:
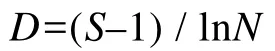
Shannon-Wiener多样度指数H′:

种类均匀度指数J′:

式中,S为种类数;N为总尾数;W为总重量;Pi为i种头足类占总重量的比例。
1.2.3 群落与环境因子的关系 采用典范对应分析(canonical correspondence analysis,CCA)对东海南部头足类 4个季节的调查数据与环境因子的关系进行分析,分析前先对头足类种类进行筛选,排除出现频率少于1个站位的的物种,以减少机会种对分析的干扰。在数据分析之前,物种数据和环境数据均进行lg(x+1)转换,以使它们的分布更接近正态分布(刘淑德等,2008)。
以上数据分析中,群落的物种多样性指数使用PRIMER5.0软件进行处理;典范对应分析(CCA)在CANOCO4.0软件进行运算,排序结果用物种-环境因子关系的双序图表示(ter Braak,1986;ter Braaket al;1988),站位图利用suffer作图软件进行绘制。
2 结果
2.1 种类组成
四个航次共采集头足类 26264尾,重 307.47kg,隶属于3目6科8属22种(表1)。其中,以乌贼目种类最多,为 11种,其次为枪形目(6种)和八腕目(5种)。各季节的头足类种类组成中,以夏季出现的种类最多,有18种。根据头足类的适温类型,暖水性种类最多,为15种,暖温性种类有6种,冷温性种类1种。
2.2 优势种组成
依据IRI的分析结果(表1),春季优势种有4种,分别为神户乌贼(Sepia kobiensis)、双喙耳乌贼(Sepiola birostrata)、多钩钩腕乌贼(Abralia multihamata)和短蛸(Octopus ocellutus),这4种优势种在调查海域出现的站位数分别为17、16、14和11个,4种优势种的尾数占头足类渔获总尾数和总重量的百分比分别为70.6%和69.6%;夏季优势种有3种,分别为双喙耳乌贼、杜氏枪乌贼和真蛸,这3种优势种在调查海域出现的站位数分别为18、17和14个,3种优势种的尾数占头足类渔获总尾数和总重量的百分比分别为49.6%和 30.3%;秋季优势种有 4种,分别为神户乌贼、真蛸、双喙耳乌贼和柏氏四盘耳乌贼,这4种优势种在调查海域出现的站位数分别为 15、14、13和13个,4种优势种的尾数占头足类渔获总尾数和总重量的百分比分别为 77.1%和 64.4%;冬季优势种有 3种,分别为杜氏枪乌贼、神户乌贼和柏氏四盘耳乌贼,这 3种优势种在调查海域出现的站位数分别为 24、17和18个,3种优势种的尾数占头足类渔获总尾数和总重量的百分比分别为78.6%和77.7%。
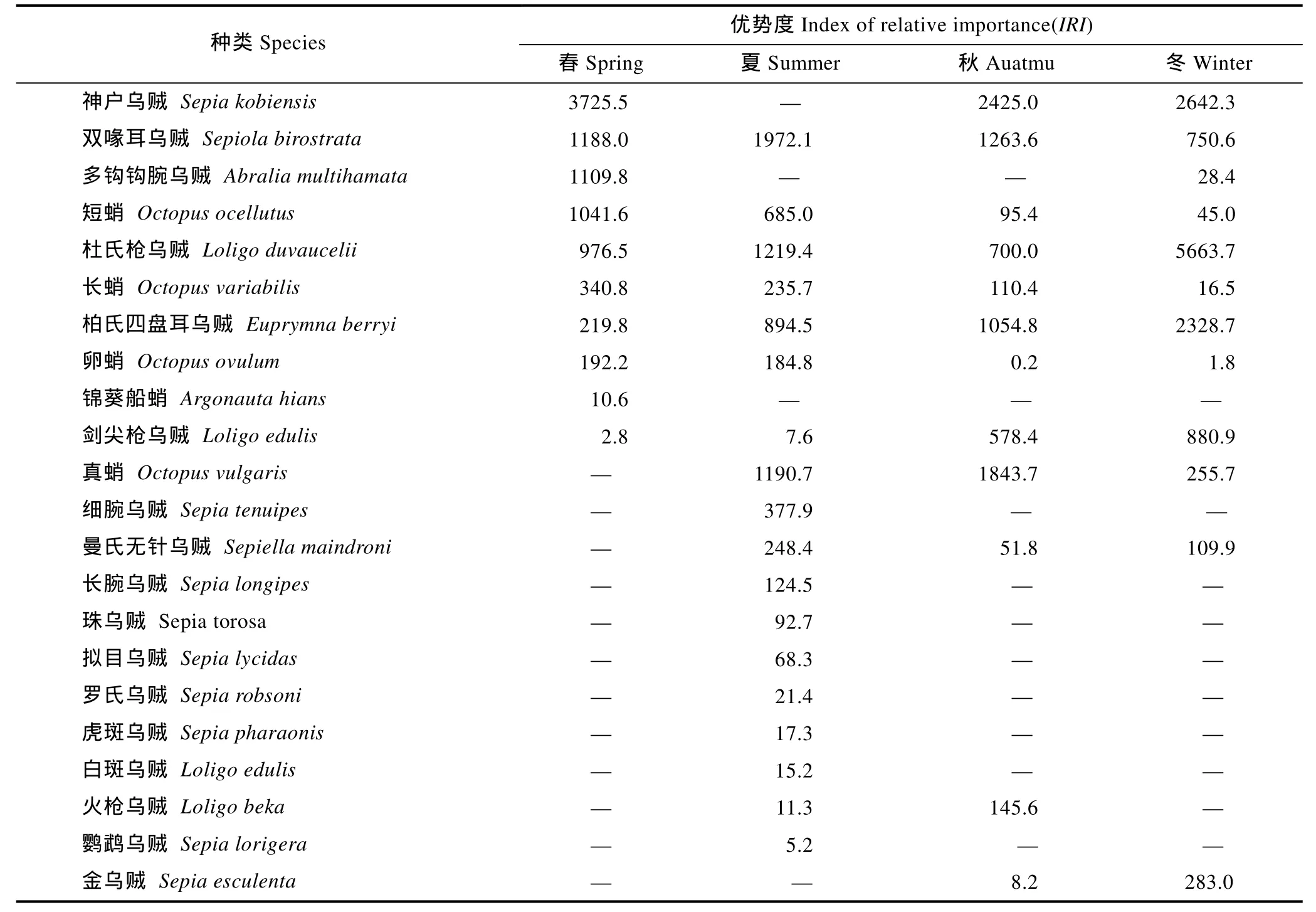
表1 东海南部海域头足类种类及其优势种组成Tab.1 The total species and dominant species component of cephalopod community in the southern East China Sea
2.3 群落多样性指数
东海南部各季节的头足类群落物种多样性指数变化如表2所示。种类丰富度指数D的变化范围为0—1.30,其中冬季最高,秋季最低;Shannon-Wiener多样性指数H′的变化范围为0—1.69,其中夏季最高,秋季最低;均匀度指数J′的变化范围为 0.02—1.00,其中秋季最高,冬季最低。调查海域的种类丰富度指数D在全年四个季节的分布情况均呈由南至北逐步下降的趋势;Shannon-Wiener多样度指数H′在春、夏、冬三季由南至北呈下降趋势,只有秋季沿纬度变化不大,且维持在一个较为稳定的水平上;种类均匀度指数J′在调查海域各个季节之间相差较小。t检验表明:种类丰富度指数D、Shannon-Wiener多样度指数H′和均匀度指数J′三个多样性指数在四个季节中的差异均不显著(R=0.7012,P>0.05)。
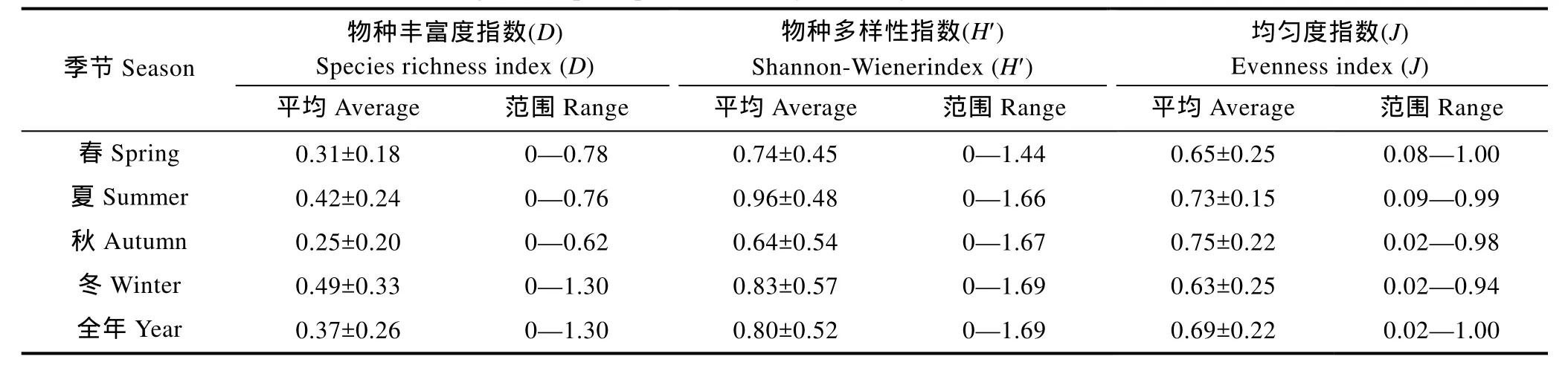
表2 东海南部头足类群落多样性指数的季节变化Tab.2 Seasonal changes of cephalopod community diversity indices in the southern East China Sea
2.4 群落与环境因子关系的CCA分析
春季,调查海域头足类群落与表层温度(ST)、表层盐度(SS)、底层温度(BT)、底层盐度(BS)和水深(D)等5个环境因子的CCA排序结果(图2)。根据计算结果,前两个排序轴的特征值(Eigenvalues)分别为0.202和0.147,前两个环境因子排序轴与种类排序轴之间的相关系数分别为0.5593和0.7546,前两个种类排序轴近似垂直(相关系数–0.0102),前两个环境排序轴的相关系数为 0,说明排序轴与环境因子间线性结合的程度较好地反映了调查站位与环境之间的关系,排序的结果是可靠的。
CCA第一排序轴与 5个环境因子的相关性均不明显,仅与水深(D)(相关系数–0.1933)有一定的负相关;第二排序轴与底层温度(BT)呈最大正相关,其值为 0.6559,而与底层盐度(BS)呈较强的负相关,其值为–0.4090;表层温度(ST)与物种第一、第二排序轴的相关性分别为–0.1353和–0.2203,表层盐度(SS)与物种第一、第二排序轴的相关性分别为–0.0121和0.0452。根据环境因子和排序轴的相关系数,影响调查海域头足类群落的主要环境因子为底层温度(BT),其次为底层盐度(BS),水深(D)、表层温度(ST)和表层盐度(SS)对群落结构的影响较小。

图2 春季头足类站位与环境因子的CCA排序Fig.2 CCA ordination biplot between sampling stations and environmental factors in spring
夏季,调查海域头足类群落与表层温度(ST)、表层盐度(SS)、底层温度(BT)、底层盐度(BS)和水深(D)等5个环境因子的CCA排序结果(图3)。根据计算结果,前两个排序轴的特征值(Eigenvalues)分别为0.209和0.122,前两个环境因子排序轴与种类排序轴之间的相关系数分别为0.7141和0.7272,前两个种类排序轴略有角度(相关系数–0.1171),前两个环境排序轴的相关系数为 0,说明排序轴与环境因子间线性结合的程度也能较好地反映了调查站位与环境之间的关系,排序的结果是较为可靠的。
CCA第一排序轴与 5个环境因子的相关性中,和底层温度(BT)成最大负相关,其值为-0.5243,与底层盐度(BS)呈较强的正相关,其值为 0.4391,与表层温度(ST)、表层盐度(SS)和水深(D)相关性不明显;第二排序轴与与5个环境因子的相关性均不明显,仅与表层盐度(SS)有一定的正相关(相关系数 0.4549)。根据环境因子和排序轴的相关系数,影响调查海域头足类群落的主要环境因子为底层温度(BT),其次为表层盐度(SS)和底层盐度(BS),水深(D)和表层温度(ST)对群落结构的影响较小。
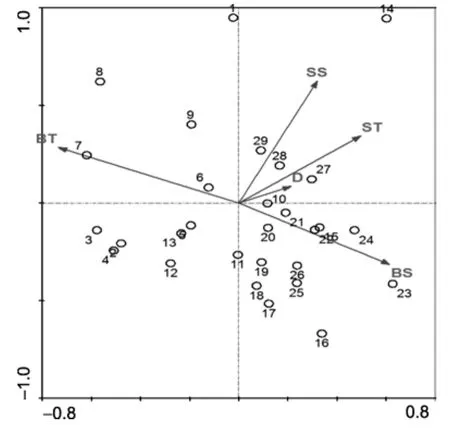
图3 夏季头足类站位与环境因子的CCA排序Fig.3 CCA ordination biplot between sampling stations and environmental factors in summer
秋季,调查海域头足类群落与表层温度(ST)、表层盐度(SS)、底层温度(BT)、底层盐度(BS)和水深(D)等5个环境因子的CCA排序结果(图4)。根据计算结果,前两个排序轴的特征值(Eigenvalues)分别为0.209和0.137,前两个环境因子排序轴与种类排序轴之间的相关系数分别为0.6540和0.7218,前两个种类排序轴略有角度(相关系数-0.1248),前两个环境排序轴的相关系数为 0,说明排序轴与环境因子间线性结合的程度也能较好地反映了调查站位与环境之间的关系,排序的结果是较为可靠的。

图4 秋季头足类站位与环境因子的CCA排序Fig.4 CCA ordination biplot between sampling stations and environmental factors in autumn
CCA第一排序轴和第二排序轴与 5个环境因子的相关性中,仅底层温度(BT)与第二排序轴呈较强的负相关,其值为–0.6335,其它环境因子与前两个排序轴之间的相关性均不大,由此可见:影响秋季头足类群落的环境因子主要是底层温度(BT)。
冬季,调查海域头足类群落与表层温度(ST)、表层盐度(SS)、底层温度(BT)、底层盐度(BS)和水深(D)等5个环境因子的CCA排序结果(图5)。根据计算结果,前两个排序轴的特征值(Eigenvalues)分别为0.127和0.078,前两个环境因子排序轴与种类排序轴之间的相关系数分别为0.6717和0.7104,前两个种类排序轴略有角度(相关系数 0.0963),前两个环境排序轴的相关系数为 0,说明排序轴与环境因子间线性结合的程度也能较好地反映了调查站位与环境之间的关系,排序的结果是较为可靠的。
CCA第一排序轴和第二排序轴与 5个环境因子的排序关系和秋季较为一致,同样是底层温度(BT)与第二排序轴呈较强的正相关,其值为 0.5324其它环境因子与前两个排序轴之间的相关性均不大,由此可见:影响冬季头足类群落的环境因子同样是是底层温度(BT)。
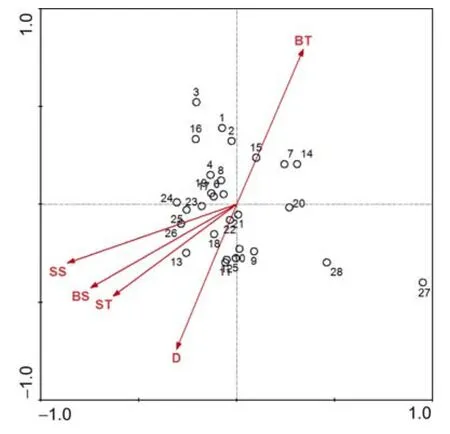
图5 冬季头足类站位与环境因子的CCA排序Fig.5 CCA ordination biplot between sampling stations and environmental factors in winter
3 讨论
3.1 头足类种类及优势种组成
根据本次调查结果,东海南部海域共有 22种,主要以乌贼目和枪形目为主,分别有11种和6种,占总种类数的77.3%。从种类的季节分布来看,以夏季出现种类最多,达到 18种,其它三个季节出现种类差别不大,分别为春季(10种),秋季和冬季(12种),这可能是由于夏季各种水系活动最为强烈,海洋环境变化幅度较大,各种海洋生物的活动范围最为广泛,调查捕获的几率相对较高有关。从种类的区系特征来看,东海南部海域主要受黑潮暖流及其西分支——台湾暖流的影响,因此本次调查的种类组成以热带和亚热带和暖水种和暖温种为主,如神户乌贼、杜氏枪乌贼、柏氏四盘耳乌贼和真蛸等。同以往调查相比,俞存根等(2009)在 2006—2007年在浙江南部海域进行的调查结果显示,共捕获头足类 36种,明显高于本次调查出现的种类,这主要是因为本次调查的网具为桁杆拖虾网,该种网具主要以捕捞底层与近底层渔业生物为主,而头足类大多有昼夜迁移和垂直分布的特点(董正之,1991),在一定程度上不能很好地反映出调查海域头足类的组成特点。与东海不同海域相比,舟山渔场及邻近海域头足类种类有25种(覃涛等,2011),而整个东海近海则有 35种(董正之,1988)。
本次调查中,头足类群落的优势种主要是神户乌贼、双喙耳乌贼、多钩钩腕乌贼、短蛸、杜氏枪乌贼、真蛸和柏氏四盘耳乌贼等。其中,神户乌贼、双喙耳乌贼和杜氏枪乌贼为主要优势种,这与俞存根等(2009)在相似海域以及李圣法等(2006)在整个东海的调查结果较为一致。与历史资料相比较,20世纪70年代以前,曼氏无针乌贼、金乌贼、白斑乌贼和虎斑乌贼等大型头足类是调查海域乃至整个东海区的主要优势种(李圣法等,2006)。而20世纪90年代以来,随着单拖作业的不断发展,捕捞力量不断加强,头足类中某些经济价值较高的种类逐渐被剑尖枪乌贼、神户乌贼以及蛸类等种类所替代(郑元甲等,1999),根据这种趋势可以看出,调查海域乃至整个东海的头足类的群落结构已经发生的很大的改变。
3.2 群落结构特征变化
根据东海南部群落生物多样性的分析表明,四个季节的物种丰富度指数(D)均维持在一个较低水平。从时间分布来看,以秋季平均数值最低,仅为0.25,冬季最高,为0.49;从空间分布来看,物种丰富度指数(D)沿纬度呈由北至南逐渐升高的趋势,沿经度方向则是近海高、外海低。这与董正之的研究结果一致,即中国沿海的头足类总体上由北向南,物种丰富度是逐渐增加的(董正之,1988)。Shannon-Wiener多样性指数H′全年的变化范围为0—1.69,其从时间分布情况看,夏季平均值最高,秋季最低;从空间分布来看,该指数与物种丰富度指数(D)的分布情况大致相当,只是在秋季略有变化,究其原因可能是与该季节是以神户乌贼、金乌贼等有针乌贼类的渔汛季节,大批生产渔船均在调查海域进行生产活动,对头足类群落的扰动及破坏较为剧烈有关。均匀度指数J在在调查海域各个季节之间相差较小,这种现象在其它游泳动物群落的研究中也同样存在(朱鑫华等,1994;李建生等,2004;刘勇等,2004)。
3.3 头足类群落与环境因子的关系
鱼类种类组成在时空上呈现一定的异质性,这与复杂的理化因子和海底地貌关系十分密切(任一平等,2006)。本文对东海南部海域的头足类与环境因子进行典范对应分析,结果表明:底层温度(BT)是影响头足类分布最为重要的环境因子,其次为底层盐度(BS),而水深(D)、表层温度(ST)和表层盐度(SS)与头足类的分布关系则不是十分明显。这主要是由于水深,尤其是底层水温是导引头足类生殖洄游和越冬洄游的一个主要外界因素,而洄游活动能够最为直接的影响某种海洋生物的分布情况。同时,以前的研究资料也印证过此种论断(李圣法等,2006;陈强等,2012)。但是,由于调查项目的局限性,诸如浮游生物的生物量、叶绿素a、营养盐、pH等生物与非生物因子对头足类分布与群落结构的影响未能涉及,这也有待于进一步的研究。
丁天明,宋海棠,2001.东海中北部海区头足类资源量的评估.水产学报,25(3):215—221
马克平,1994.生物多样性研究的原理与方法.北京:中国科技出版社,23—27
朱鑫华,吴鹤州,徐凤山等,1994.黄渤海沿岸水域游泳动物群落多样性及其相关因素的研究.海洋学报,16(3):102—112
任一平,徐宾铎,叶振江等,2005.青岛近海春、秋季渔业资源群落结构特征的初步研究.中国海洋大学学报,35(5):792—798
刘 勇,李圣法,程家骅,2006.东海、黄海鱼类群落结构的季节变化研究.海洋学报,28(4):108—114
刘淑德,线薇微,刘 栋,2008.春季长江口及其邻近海域鱼类浮游生物群落特征.应用生态学报,19(10):2284—2292
李圣法,严利平,李惠玉等,2006.东海区头足类群聚空间分布特征的初步研究.中国水产科学,13(6):936—944
李建生,李圣法,任一平等,2004.长江口渔场渔业生物群落结构的季节变化.中国水产科学,11(5):432—439
宋海棠,丁天明,余匡军等,1996.东海北部头足类的种类组成和数量分布.浙江海洋学院学报,18(2):99—106
宋海棠,丁天明,徐开达,2008.东海头足类的数量分布与可持续利用.中国海洋大学学报,38(6):911—915
宋海棠,丁天明,徐开达,2009.东海经济头足类资源.北京:海洋出版社,1—120
陈 强,王家樵,张雅芝等,2012.福建闽江口及附近海域和厦门海域头足类种类组成的季节变化.海洋学报,34(3):179—184
国家技术监督局,1991.海洋调查规范——海洋生物调查.北京:中国标准出版社,1—324
郑元甲,凌建忠,严利平等,1999.东海区头足类资源现状与合理利用的探讨.中国水产科学,13(6):936—944
俞存根,虞聪达,宁 平等,2009.浙江南部外海头足类种类组成和数量分布.海洋渔业,31(1):27—33
徐开达,朱文斌,李鹏飞等,2008.东海北部浅海渔场头足类的数量分布.浙江海洋学院学报,27(1):27—31
黄梓荣,2008.南海北部陆架区头足类的种类组成和资源密度分布.南方水产,4(5):1—7
董正之,1988.中国动物志:软体动物门:头足纲.北京:科学出版社,17—21
董正之,1991.世界大洋经济头足类生物学.济南:山东科学技术出版社,2—30
董正之,1992.头足类的分布特点及其区系特征.黄渤海海洋,10(4):37—43
覃 涛,俞存根,陈全震等,2011.舟山渔场及邻近海域头足类(Cephalopod)种类组成和数量分布.海洋与湖沼,42(1):124—130
Ludwig J A,Reynolds J F,1988.Statistical Ecology.New York:John Wiley &Sons,213—256
Margalef R,1957.Information theory in ecology.General Systematics Advancing Frontiers Plant Sci,3:37—71
Pielou E C,1975.Ecological diversity Diversity.New York:A Wiley-interscience Publication,286—287
Pinkas L,Oliphant M S,Iverson L K,1971.Food habits of albacore,bluefin tuna,and bonito in California waters.Clif Dep Fish Game.Fishery Bulletin,152:1—105
ter Braak C J F,1986.Canonical correspondence analysis:A new eigenvector technique formu ltivariate direct grad ient analysis.Ecology,67:1167— 1179
ter Braak C J F,Prentice I C,1988.A theory of grad ient analysis.Advances in Ecological Research,18:271—317
Wilhm J L,1968.Use of biomass units in Shannon formula.Ecology,49:153—156
