异质环境下入侵植物薇甘菊的适应性与繁殖特性
2014-02-09徐高峰申时才张付斗
徐高峰,申时才,张付斗
云南省农业科学院农业环境资源研究所,云南 昆明 650205
异质环境下入侵植物薇甘菊的适应性与繁殖特性
徐高峰,申时才,张付斗*
云南省农业科学院农业环境资源研究所,云南 昆明 650205
薇甘菊(Mikania micrantha H.B.Kunth)为世界上最具有入侵性和危害性的外来入侵物种之一,对其控制与管理已成为长期以来世界性难题。了解入侵植物薇甘菊在异质环境下的适应性与繁殖特性对安全有效预警、监测和防治该入侵物种具有重要意义。本文于薇甘菊盛花期,根据薇甘菊入侵生境的光照条件、群落结构和生长方式差异,在薇甘菊常见的4种入侵生境(林地、荒地、农田和和河边)选取10个样地,调查研究了不同生境条件下薇甘菊的开花结实和繁殖分配,并运用植物叶片功能性状的研究方法研究了不同生境条件下薇甘菊的比叶面积(SLA)、叶干物质比例(LMF)和茎干物质比例(SMF)的3种叶片功能性状。研究结果表明,生境条件对薇甘菊的开花结实和繁殖分配具有显著影响。其中抛荒农田(样地Ⅷ)和河流边缘(样地Ⅸ和Ⅹ)薇甘菊的总花数、种子量、生殖枝茎生物量分配和花生物量分配均显著大于其他生境,显示在有利于其生长的条件下,薇甘菊种群倾向于有性繁殖;而在不利于薇甘菊生长的低光照林地(样地Ⅰ)和种间竞争强的农田红薯地(样地Ⅵ)生境条件下,薇甘菊的总花数和种子量明显小于其他生境,但营养枝叶生物量分配显著大于其他样地,而茎生物量分配值则处于中间值,表明在不利于其生长的生境条件下,薇甘菊通过提高营养枝茎的生物量分配和叶的生物量分配来适应,其种群则更倾向于克隆繁殖。通过分析不同生境条件下薇甘菊的叶片功能性状,结果表明,在不利于薇甘菊生长的低光照林地(样地Ⅰ)和种间竞争强的农田红薯地(样地Ⅵ)生境条件下,薇甘菊的叶面积和茎干物质比例(SMF)显著小于其他样地,且彼此差异不显著;但比叶面积(SLA)和叶干物质比例(LMF)则显著大于其他样地,显示薇甘菊可通过提高比叶面积(SLA)和叶干物质比例(LMF),降低茎干物质比例(SMF)来适应低光照和种间强竞争等不利生境;而在抛荒农田(样地Ⅷ)和河流边缘(样地Ⅸ和Ⅹ)有利于其生长的生境条件下,薇甘菊则通过相反的策略来适应环境。以上结果表明薇甘菊通过调节各功能性状之间的权衡来适应环境,对异质环境条件具有不同的生长与繁殖适应对策,具有强的生态适应性。
薇甘菊;比叶面积;叶干物质比例;茎干物质比例;繁殖分配
薇甘菊(Mikania micrantha H.B.Kunth)为世界上最具有入侵性和危害性的外来入侵物种之一(Mini和Abraham, 2005; Maja等, 2008),原产于中美洲和南美洲,19 世纪传播至东南亚、中国香港和广东,20世纪50年代由缅甸边境传入云南省(Zhang等, 2004; 昝启杰等, 2000; 杜凡等, 2006)。在入侵地,薇甘菊通过快速生长、攀援、缠绕、覆盖或释放化感物质等方式,影响本地植物的生长和种群的自然更新,破坏土壤结构和土壤微生物,导致本地物种灭绝和生物多样性降低,严重危及入侵地生态系统的安全,对其控制与管理已成为长期以来世界性难题之一 (冯惠玲等, 2002; Li 等, 2006)。
入侵物种传入新的生境通常是少数个体和小种群,其如何突破瓶颈效应,在入侵地建立种群并扩张蔓延是国内外研究的热点与难点?目前大多数研究表明,入侵物种对入侵生境在环境选择压力下适应性响应能力直接决定其入侵性与扩散性,而群落的可侵入性则决定其能否成功入侵与扩散(Richards等, 2006; Schoeppner和Relyea, 2009; Matesanz等, 2010)。因此,了解入侵植物对环境的适应特征,预测入侵植物种群、群落乃至整个生态系统对竞争、气候变化以及土地利用变化的响应,对探索安全有效的防控措施具有重要意义。
植物功能性状是植物在长期进化过程中适应不同环境的结果,能有效地利用植物的生理、形态和生活史等特征,反映个体、种群、群落和生态系统水平上的生物之间、生物与环境之间的相互作用,揭示生物与生长环境间的相互关系(Violle等, 2007; Westoby和Wright, 2006)。目前,大多数植物生态学家认为,在众多的植物性状中,植物的一些叶片性状与植物的生长对策及植物利用资源的能力紧密联系,能够反应植物适应环境变化所形成的生存对策。如比叶面积(SLA)、叶干物质比例(LMF)和茎干物质比例(SMF)反映了植物叶片捕获光照资源的能力和生物量分配方式,是重要的植物叶片功能性状(Poorter, 1999; Markesteijn和Poorter)。而对比分析异质生境下薇甘菊的比叶面积(SLA)、叶干物质比例(LMF)和茎干物质比例(SMF)的差异,对探讨异质环境下薇甘菊的生态适应性和生态功能具有重要意义。
长期以来,国内外学者广泛研究了生境条件对入侵植物薇甘菊的繁殖能力和表型可塑性的影响。如杨期和等研究了土壤肥力、水分和生境郁闭度对薇甘菊开花结实的影响,表明土壤湿度大,薇甘菊的花数多,结实率高,花期长(杨期和等, 2003)。张炜银等研究了不同群落中薇甘菊种群的种子生产特征,显示群落的光照强度显著影响薇甘菊的种子量,并影响其繁殖分配,如在乔木中的薇甘菊以有性生殖为主,在草丛中的薇甘菊以无性繁殖为主(张炜银等, 2003;2005)。邓雄研究发现薇甘菊能够在很短的时间内对外界光照的变化进行形态适应性和生理可塑性响应(邓雄, 2010)。作者前期也研究了土壤水分对薇甘菊的克隆繁殖能力和形态特征的影响,结果表明在干旱(土壤含水量为12.5%)和淹水(水深1 cm)条件下,薇甘菊均可通过营养繁殖体克隆繁殖并生长(徐高峰等, 2013)。薇甘菊入侵生境广泛,现有报道多研究单一环境因子对薇甘菊开花结实与表型可塑性的影响,而对不同生境中薇甘菊的叶片功能性状和繁殖特征的研究非常有限。因此,本文以不同生境(林地、荒地、农田和和河边)条件下的薇甘菊种群的叶片功能性状、开花结实和生物量分配为研究对象,探讨不同生境条件下薇甘菊的生态适应性和繁殖特征,以期为科学防控薇甘菊入侵提供理论依据。
1 材料和方法
1.1 研究区域概况
研究区域位于云南省德宏州陇川县(N 24°08′~24°39′,E 97°39′~98°17′),海拔780~2618.8 m,属南亚热带季风气候,冬无严寒、夏无酷暑,历年平均气温18.9 ℃,降雨量1595 mm,降水季节分配不均,全年75%~85%的降水集中在5—10月份。研究区是世界生物多样性的核心区域之一,也是云南省薇甘菊入侵危害时间最长的区域之一,区域内薇甘菊分布广泛。
1.2 样地设置
样地的设立具备以下特点:具有代表性,薇甘菊发生严重,多数地区薇甘菊的覆盖度为100%,同一研究区薇甘菊具有相同的生长发育期,地势和土壤类型基本一致,植物的分布基本均匀。为避免放牧、经贸往来和人工防治等人为因素对研究造成影响,本研究样地选择在中缅国境线中国区一侧进行。该区域人烟稀少(多数区域为无人区),植被受人为干扰少。在薇甘菊常见的4种入侵生境(林地、荒地、农田和和河边),根据生境的光照水平、土壤水分和群落结构等差异选取10个不同的小生境进行调查研究,各生境薇甘菊种群盖度为70%~100%,具体情况见表1。
1.3 调查方法
2013年11月25—30日,在薇甘菊盛花期,选择薇甘菊密度较为均一的地段,以随机样方法在每样地随机选择20个0.25 m2(0.5 m×0.5 m)的样方,共计200个样方,调查样方内薇甘菊的花枝数、小花数等。由于薇甘菊为藤本植物,野外条件下枝条间相互缠绕交错,很难判断其主茎与分枝且将单株单独分开。因此本研究采用拔除法,既取样时将取样框内薇甘菊植株齐地面全部剪下,用保鲜袋低温保存至室内。将采集回来的薇甘菊的地上部分按照营养枝、生殖枝分开, 并除去植株表面灰尘。营养枝和生殖枝再按茎、叶、花序、种子等构件分开。2014年2月15—20日,薇甘菊种子成熟时, 对薇甘菊种群进行种子生产特征的研究:每一样方内随机剪取100个头状花序, 收集其所结的饱满种子, 统计种子数。
1.4 测量方法
根据每样方分开后的营养枝和生殖枝构件,随机抽取20个生殖枝统计薇甘菊的头状花序数目和小花数。并在每样方随机抽取100片位于薇甘菊植株中下部叶片,用Li-3000A型叶面积仪测定叶面积。将各构件分别装入纸袋于80 ℃烘至恒质量,冷却后用电子天平(精确度0.0001 g)立即称量,测定薇甘菊叶片、地上茎、花等质量。
1.5 数据分析
叶面积与叶干质量的比值为SLA(cm2·g-1),叶干质量与叶、茎总干质量的比值为LMF (g·g-1),茎干质量与叶、茎总干质量的比值为SMF(g·g-1) (Cornelissen, 2003)。计算公式如下:
比叶面积(SLA)=叶片面积(cm2)/叶片干质量(g)
叶干物质比例(LMF)=叶片干质量/(叶片干质量+茎干质量)
茎干物质比例(SMF)=茎干质量/(叶片干质量+茎干质量)
所有数据采用DPS v9.01版软件进行分析,采用单因素方差分析(One-Way ANOVA)检验,并用Duncan’s新复极差法检验进行多重比较,分析不同数据组间的差异。
2 结果与分析
2.1 不同生境条件下的薇甘菊叶片功能性状
2.1.1 叶面积
不同生境条件下薇甘菊的平均叶面积大小不一,具体表现为: 样地Ⅷ薇甘菊的平均叶面积最大,且与样地Ⅹ薇甘菊的叶面积差异不显著,但显著大于其它样地薇甘菊的叶面积。样地Ⅰ和Ⅵ薇甘菊叶面积显著小于其他样地,但彼此差异不显著。研究结果也表明同一生境不同的光照、群落结构和生长方式对薇甘菊的叶面积也有显著影响,表现为:在林地,薇甘菊的叶面积随着光照强度的增加表现为先增大后减小;而在路边荒地,当薇甘菊入侵地以外来种为主的群落结构时,其叶面积大于以本地种为主的群落结构;而在农田,抛荒地薇甘菊的叶面积显著大于红薯和甘蔗种植地;但在河边,攀援生长和匍匐生长的薇甘菊叶面积大小虽存在差异,但不显著(图1 A)。
2.1.2 比叶面积(SLA)
由图1 B可知,薇甘菊的比叶面积大小在各样地间也呈现出不同程度的差异,表现为:样地Ⅰ薇甘菊的比叶面积为891.02 cm2·g-1,显著大于其他样地;样地Ⅴ薇甘菊的比叶面积最小,但和样地Ⅲ、Ⅳ薇甘菊的比叶面积差异不显著。研究结果也表明同一生境不同的光照、群落结构和生长方式对薇甘菊的比叶面积也有显著影响,表现为:在林地,薇甘菊的比叶面积随着光照强度的增加逐渐变小,且差异显著;而在路边荒地,两种不同群落结构的薇甘菊比叶面积间无显著差异;在农田,栽种红薯使薇甘菊的比叶面积显著增加;但在河边,攀援生长和匍匐生长的薇甘菊比叶面积差异不显著。
2.1.3 叶干物质比例(LMF)
薇甘菊的叶干物质比例大小在各样地间也呈现出不同程度的差异,表现为:样地Ⅰ薇甘菊的叶干物质比例最大,且与样地Ⅵ差异不显著,但显著大于其他样地薇甘菊的叶干物质比例。样地Ⅴ和Ⅸ薇甘菊叶干物质比例显著小于其他样地,但彼此差异不显著。同一生境不同的光照、群落结构和生长方式对薇甘菊的叶干物质比例也有显著影响,表现为:在林地,薇甘菊的叶干物质比例随着光照强度的增加表现为先减小后增大;而在路边荒地,当薇甘菊入侵地以外来种为主的群落结构时,其叶干物质比例显著大于以本地种为主群落结构的薇甘菊叶干物质比例;而在农田,红薯地薇甘菊的叶干物质比例显著大于甘蔗种植地和抛荒地;在河边,攀援生长的薇甘菊叶干物质比例显著小于匍匐生长(图1C)。
2.1.4 茎干物质比例(SMF)
由图1D可知,样地Ⅴ和样地Ⅸ薇甘菊的茎干物质比例最大,均为0.63 g·g-1,样地Ⅰ的茎干物质比例最小,为0.51 g·g-1,与样地Ⅵ无显著差异,但显著小于其他样地。同一生境不同的光照、群落结构和生长方式对薇甘菊的茎干物质比例也有明显影响,表现为:在林地,薇甘菊的茎干物质比例随着光照强度的增加表现为先增大后减小;而在路边荒地,当薇甘菊入侵地以外来种为主的群落结构时,其茎干物质比例明显小于以本地种为主群落结构的薇甘菊茎干物质比例;而在农田,红薯地薇甘菊的茎干物质比例显著小于甘蔗种植地和抛荒地;在河边,攀援生长的薇甘菊茎干物质比例显著大于匍匐生长。
2.2 生境条件对薇甘菊开花结实的影响
不同生境条件对薇甘菊的开花结实存在显著影响,农田荒地(样地Ⅷ)薇甘菊的花枝数、花序数和总花数最大,但和样地Ⅴ、Ⅸ和Ⅹ差异不显著。样地Ⅵ薇甘菊的花枝数、花序数、总花数和种子量最小,但和样地Ⅰ和Ⅶ差异不显著。
同一生境不同的光照、群落结构和生长方式对薇甘菊的开花结实也有显著影响。如林地生态系统,随着光照强度的最强,薇甘菊的花枝数、花序数、总花数和种子量均显著增加。群落结构对薇甘菊的开花结实也有显著影响,如路边荒地,薇甘菊的花枝数、花序数、总花数和种子量在以入侵物种飞机草为主的生态系统中均显著小于本地物种群落。在农田生态系统中,种植红薯和甘蔗能显著降低薇甘菊的总花数和种子量。在对河边攀援生长和匍匐生长的薇甘菊花枝数、花序数、总花数和种子量调查发现,生长方式对薇甘菊开花和种子影响不显著(表2)。
2.3 不同生境条件下薇甘菊地上组织的生物量积累与分配
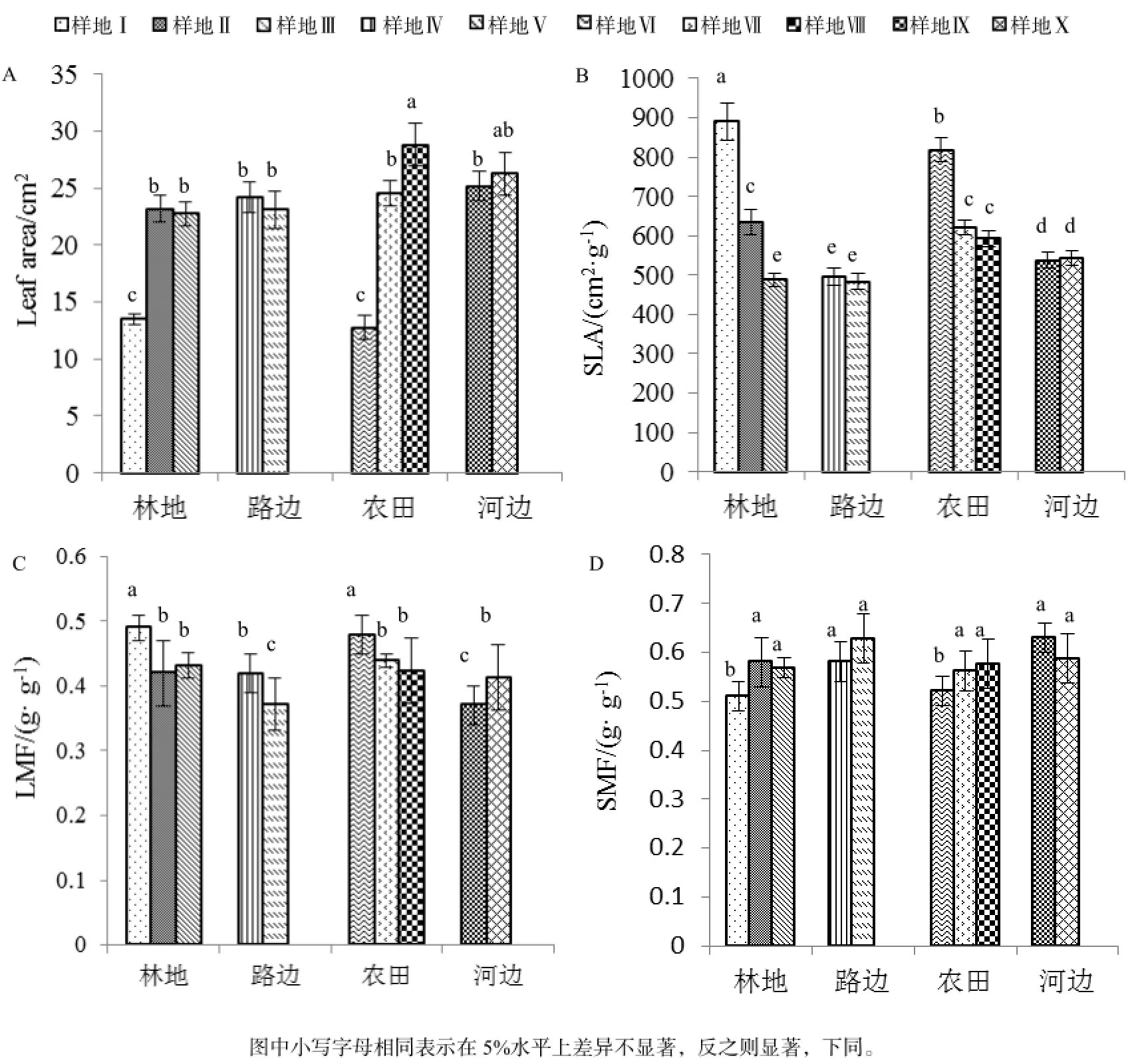
图1 不同生境条件下薇甘菊叶片功能性状比较Table 1 General variation in leaf structural traits of M. micrantha in different habitats

表2 生境条件对入侵植物薇甘菊开花结实的影响Table 2 Effects of different habitats on reproductive characteristics of Mikania micrantha
生境条件对薇甘菊的生物量积累存在显著影响。样地Ⅷ薇甘菊的地上生物量和花生物量最大,但和样地Ⅴ、Ⅸ和Ⅹ的薇甘菊地上生物量差异不显著,表明抛荒农田、水源地和路边空地均有利于薇甘菊生长。样地Ⅵ薇甘菊的地上生物量和花生物量最小,但和样地Ⅰ和Ⅶ差异不显著,表明郁闭度很高的森林和强的种间竞争均不利于薇甘菊生长。另外研究结果也表明样地Ⅴ营养枝茎生物量最大,而生殖枝茎的最大生物量为样地Ⅹ(表3)。
不同生境条件对薇甘菊种群各构件的生物量分配具有显著影响。样地Ⅳ薇甘菊的营养枝茎生物量分配最大,但和样地Ⅱ、Ⅴ、Ⅶ和Ⅸ无显著差异,而样地Ⅹ薇甘菊的生殖枝茎的生物量分配最大。样地Ⅵ的薇甘菊营养枝叶的生物量分配最大,但与样地Ⅰ无显著差异。而薇甘菊的花生物量分配的最大值为样地Ⅸ,但和样地Ⅱ、Ⅲ、Ⅴ、Ⅷ和Ⅹ无显著差异(表4)。
3 讨论
植物生长所必须的光、水、CO2和矿质元素等资源在自然界中呈异质性分布,迫使入侵植物做出形态、生理、生活史特征以及可塑性响应(Westoby和Wright, 2006)。目前,大多数植物生态学家认为,在众多的植物性状中,植物的一些叶片性状与植物的生长对策及以植物利用资源的能力紧密联系,能够反应植物适应环境变化所形成的生存对策。如比叶面积(SLA)可以反映植物获取光照资源的能力,生长在资源较为丰富的环境中的物种通常具有高的比叶面积, 而低比叶面积的植物能够更好地适应资源贫瘠和干旱的环境(Lambers和Poorter, 1992; Wilson等, 1999)。本研究结果表明,比叶面积的变化是入侵植物薇甘菊适应各种异质生境的重要手段之一,其中光照和种间竞争对薇甘菊的比叶面积具有显著影响,而群落结构和生长方式对薇甘菊的比叶面积则影响不显著。如在群落结构和土壤类型相似的林地(样地Ⅰ、样地Ⅱ和样地Ⅲ),随着光照强度的增加,样地Ⅰ薇甘菊的比叶面积显著大于样地Ⅱ和样地Ⅲ,表明在郁闭度较大的林地,薇甘菊通过提高叶片投入,扩大叶面积来适应林下阴暗的环境。而在面对较强的竞争压力,如在农田红薯地(样地Ⅵ),薇甘菊的比叶面积也显著增加,显示薇甘菊通过扩大叶面积来提高竞争能力。而在路边和河边,薇甘菊有相对较低的比叶面积,其叶寿命相对较长,长的叶寿命不断累积叶面积,提高光截获能力,在一定程度上使得植物获得高的能量积累。

表3 不同生境条件对薇甘菊地上部生物量积累的影响Table 3 Effects of different habitats on above ground biomass accumulation of Mikania micrantha

表4 不同生境条件对薇甘菊地上部生物量分配的影响Table 4 Effects of different habitats on above ground biomass allocation of Mikania micrantha
生物量分配方式的变化是植物应对环境胁迫的有效方式之一,植物的叶干物质比例(LMF)和茎干物质比例(SMF)可以揭示植物的生物量分配方式。在生长发育过程中,植物总是不断地调整其生长和生物量的分配策略来适应环境(Zhu等, 2009)。高的叶干物质比例有利于幼苗截获更多的光能, 获得更多的光能量。本研究中,如样地Ⅱ和样地Ⅲ,随着光照强度的增加,薇甘菊的叶干物质比例增加,茎干物质比例降低;而在有攀援物的条件下(样地Ⅸ),薇甘菊将更多的生物量分配给茎干(高的茎干物质比例),以促进其高生长。光照是限制藤本植物生长的主要因子(Letcher 和Chazdon, 2009),在郁闭度大的林下和覆盖度大的红薯地,为了截获光能而将更多的生物量分配给叶片(高的叶干物质比例)。上述结果表明,入侵植物薇甘菊具有强的生态适应性,在光环境较好的生境,薇甘菊将更多的生物量投入到茎干上,这有利于它们获取更多的资源,避免被周围的植物遮蔽,同时对茎干投入的增加加强了物理强度,也是植物应对风胁迫和重力胁迫的一种适应。
在植物整个生长发育阶段中,生长、繁殖和防御等各种功能对有限的资源始终存在着竞争,植物的能量分配格局反映了植物在生长发育过程中对环境的响应规律和能量分配对策(Cheplick, 1995; 赵磊等,2007)。植物繁殖格局不仅受到环境等因素的影响,而且在有性繁殖与未来生存、营养体生长或无性繁殖等功能间存在着权衡(Trade-offs)。植物在进行有性繁殖的过程中,需要在不同的组织水平上不断地在生殖器官和相应的营养支撑器官之间进行资源分配, 建立不同的生活史对策来适应环境的选择压力。植物产生的种子数量多少既能反映其生物学特性, 也能反映其对环境的适应方式以及环境对其有性生殖的影响。种子产量的多少对植物是否有足够的种子经散布后形成土壤种子库, 度过不良环境, 在适宜条件下补充幼苗库, 保证种群的稳定或进一步发展具有重要作用。本研究结果说明,在光照、土壤水分较为充足的条件下(样地Ⅷ、样地Ⅸ和样地Ⅹ),薇甘菊的生殖枝和花的生物量分配显著大于其他生境,而在光照弱(样地Ⅰ)和竞争强(样地Ⅵ)的生境,薇甘菊营养枝生物量分配比例则显著大于其他生境,表明入侵植物薇甘菊在有性和性繁殖之间的资源分配存在着权衡。当在境条件有利于薇甘菊生长发育时,薇甘菊将增加生殖枝的投资,倾向于有性繁殖方式;而当环境条件不利时,则将增加营养枝的投资,倾向于克隆繁殖繁殖。
4 结论
综上所述,在不同的生境下,入侵植物薇甘菊通过调整功能性状:比叶面积(SLA)、叶干物质比例(LMF)和茎干物质比例(SMF)以及营养枝、生殖枝和花的生物量分配来适应异质环境中资源的变化。当外界资源(包括光照、水分和种间竞争等)条件有利时, 薇甘菊种群倾向于有性繁殖,而在资源限制性(资源胁迫)条件下, 薇甘菊种群倾向于无性繁殖(克隆)提高自己及其后代的幸存几率。
CHEPLICK G P. 1995. Life history trade-offs in Aphibromus scabrivalvis (Poaceae): Allocation to clonal growth, storage, and cleistogamous reproduction[J]. American Journal of Botany, 82(5): 621-629.
CORNELISSEN J H C, LAVOREL S, GARNIER E, et al. 2003. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide[J]. Australian Journal of Botany, 51: 335-380.
LAMBERS H, POORTER H. 1992. Inherent variation in growth rate between higher plants: a search for physiological cause and ecological consequences[J]. Advances in ecological research, 23:188-242.
LETCHER S G, CHAZDON R L. 2009. Lianas and self-supporting plants during tropical forest succession[J]. Forest Ecology and Management, 257: 2150-2156.
LI W H, ZHANG C B, JIANG H B, et al. 2006. Changes in soil microbial community associated with invasion of the exotic weed, Mikania micrantha H. B. K[J]. Plant and Soil, 281(1/2): 309-324.
MAJA W, ZERBE S, KUO Y L. 2008. Distribution and ecological range of the alien plant species Mikania micrantha Kunth(Asteraceae) in Taiwan [J]. Journal of Ecology and Field Biology, 31(4): 227-290.
MARKESTEIJN L, POORTER L. 2009. Seedling root morphology and biomass allocation of 62 tropical tree species in relation to drought-and shade-tolerance[J]. Journal of Ecology, 97: 311-325.
MATESANZ S, GIANOLI E, VALLADARES F. 2010. Global change and the evolution of the phenotypic plasticity in plants[J]. Annals of the New York Academy of Sciences, 1206: 35-55.
MINI A, ABRAHAM C T. 2005. Biology of mile-a minute weed (Mikania micrantha H.B.K), an alien invasive weeds in kerala[J]. Indian Journal of Weeds Science, 37(2): 153-154.
POORTER L. 1999. Growth responses of 15 rain-forest tree species to a light gradient: the relative importance of morphological and physiological traits[J]. Functional Ecology, 13: 396-410.
RICHARDS C L, BOSSDORF O, MUTH N Z, et al. 2006. Jack of all trades, master of some? On the role of phenotypic plasticity in plant invasions[J]. Ecology Letter, 9: 981-993.
以中国知网(CNKI)数据库为来源,点击“高级搜索”,在搜索栏中分别输入作者“许钧”、主题词“翻译”进行文献计量,所获取和计算的数据截取1987—2018年份以表格呈现如下。
SCHOEPPNER N M, RELYEA R A. 2009. Phenotipic plasticity in response to fine-grained environmental variation in predation[J]. Functional Ecology, 23: 587-594.
VIOLLE C, NAVAS M L, VILE D, et al. 2007. Let the concept of trait be functional![J] Oikos, 116: 882-892.
WESTOBY M, WRIGHT I J. 2006. Land-plant ecology on the basis of functional traits[J]. Trends in Ecology & Evolution, 21: 261-268.
WILSON P, THOMPSON K, HODGSON J. 1999. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies[J]. New phytologist, 143:155-162. ZHANG L Y, YE W H, CAO H L et al. 2004. Mikania micrantha H.B.K in China: An overview[J]. Weed Research, 44 (1): 42-49.
ZHU Y J, DONG M, HUANG Z Y. 2009. Response of seed germination and seedling growth to sand burial of two dominant perennial grasses in Mu-Us sandy grassland, Semiarid China[J]. Rangeland Ecology & Management, 62: 337-344.
杜凡, 杨宇明, 李俊清, 等. 2006, 云南假泽兰属植物及薇甘菊的危害[J].云南植物研究, 28(5): 505-508.
冯惠玲, 曹洪麟, 粱晓东, 等. 2002. 薇甘菊在广东的分布与危害[J]. 热带亚热带植物学报, 10(3): 263-270.
杨期和, 冯惠玲, 叶万辉, 等. 2003. 环境因素对薇甘菊开花结实影响初探[J]. 热带亚热带植物学报, 11(2):123-126.
徐高峰, 申时才, 张付斗, 等. 2013. 土壤水分对薇甘菊不同繁殖体单位存活能力和植株表型可塑性影响[J]. 中国农业科学, 2013, 46(15): 3134-3141.
昝启杰, 王勇军, 王伯荪, 等. 2000. 外来杂草薇甘菊的分布及危害[J].生态学杂志, 19 (6): 58-61.
张炜银, 李鸣光, 臧润国, 等. 2005. 外来杂草薇甘菊种群土壤种子库动态[J]. 武汉植物学研究, 23(1): 49-52.
张炜银,李鸣光,王伯荪, 等. 2003. 外来杂草薇甘菊在不同群落中的种子生产特征[J]. 武汉植物学研究, 21(2): 143-147.
赵磊, 智颖飙, 李红丽, 等. 2007. 初始克隆分株数对大米草表型可塑性及生物量分配的影响[J]. 植物生态学报, 31(4): 607-612.
Adaptability and Reproductive Characteristics of Mikania micrantha H.B.K under Different Habitats
XU Gaofeng, SHEN Shicai, ZHANG Fudou*
Agricultural Environment & Resource Research Institute, Yunnan Academy of Agricultural Sciences, Kunming 650205, China
Mikania micrantha H.B.K is considered one of the world’s most invasive and destructive species, and its control and management has become a difficult problem in the world. Currently, many domestic and foreign research showed that invasive alien plants has stronger adaptation ability to the heterogeneous environment than native species. Research on adaptability and reproductive characteristics under different habitats has provided the scientific ways for implement effective monitoring, early-warning and control of M. micrantha. In this paper, we selected the 10 habitats which represented 4 population (woodland, wasteland, farmland and riverside) in full-bloom stage of M. micrantha, sexual reproduction, reproductive allocation and the major functional traits of specific leaf area (SLA), leaf mass fraction (LMF), specific stem density (SSD) and stem mass fraction (SMF) on M. micrantha under different habitats were measured. The results showed sexual reproduction and reproductive allocation were effected significantly of different habitats of M. micrantha. Total flower number, seed number, biomass allocation of stem of reproductive shoots and flower of barren field (plot Ⅷ) and riverside (plots Ⅸ and Ⅹ) were significantly higher than other plots, populations tend to sexual reproduction. However, the total flower number and seed number were decreased significantly, but the nutrition branches and leaf biomass allocation were significantly larger than the other plots, and the value of stem biomass allocation is in the middle of the range when the population of invasive alien species M. micrantha in unfavorable light condition (such as sample I) and community structure(such as sample VI). Which showed to adapt the adverse habitats (such as sample I, IV, VI), invasive alien species M. micrantha enhanced the biomass allocation of stem on vegetative shoot and leaves, the population was more inclined to clonal propagation. there were also significant differences among varied habitats in functional traits. In the low light (plotⅠ) and strongly interspecific competition (plotⅥ), leaf area and SMF were lowest than others, but showed no significant difference of each other, but SLA and LMF were significantly greater than the others. Invasive alien species M. micrantha adapt to the stress habitat (such as low light, drought and interspecific competition) by increasing SLA and LMF and decreasing leaf area and SMF, but using opposite strategy to adapt to the favorable habitat. Our study suggests that invasive alien species M. micrantha using different reproductive strategies, such as changes the value of different functional traits, to adapt to environmental.
Mikania micrantha H.B.Kunth; specific leaf area; leaf mass fraction; stem mass fraction; reproductive allocation
Q948
:A
:1674-5906(2014)08-1258-07
徐高峰,申时才,张付斗. 异质环境下入侵植物薇甘菊的适应性与繁殖特性[J]. 生态环境学报, 2014, 23(8): 1258-1264.
XU Gaofeng, SHEN Shicai, ZHANG Fudou. Adaptability and Reproductive Characteristics of Mikania micrantha H.B.K under Different Habitats [J]. Ecology and Environmental Sciences, 2014, 23(8): 1258-1264.
云南省应用基础研究重点项目(2010CC002)
徐高峰(1979年生),男,副研究员,硕士,从事入侵生态学研究。E-mail:xugaofeng1059@163.com
*通信作者:张付斗(1971年生),研究员,硕士,从事杂草学和生态学研究。
2014-03-28
