宁夏盐池人工封育区植物群落结构及多样性
2014-02-08郝智如张克斌
郝智如,张克斌
北京林业大学水土保持和荒漠化防治教育部重点实验室,北京 100083
宁夏盐池人工封育区植物群落结构及多样性
郝智如,张克斌*
北京林业大学水土保持和荒漠化防治教育部重点实验室,北京 100083
为探究宁夏盐池人工封育区植物群落的结构和变化规律,将研究区划分为核心区(E)、边缘区(E1)、外围区(E2)3个区域,采用小样方法对各区域内的植物种名、株数、盖度、高度、生物量等进行调查,并结合多样性指数(物种多样性指数、群落优势集中性指数、群落均匀度指数和物种丰富度指数)、主分量分析和极点排序的方法对其进行分析。结果表明:宁夏盐池人工封育区以菊科和豆科为优势种,区域E 2011年共有植物9科21种,菊科和豆科占52.4%;2012年共有植物7科20种,菊科和豆科占50.0%;2013年共有植物9科22种,菊科和豆科占50.0%。区域E12011年共有植物8科18种,菊科和豆科占55.6%;2012年共有植物7科16种,菊科和豆科占50.0%;2013年共有植物9科16种,菊科和豆科占50.0%。区域E22011年共有植物6科15种,菊科和豆科占60.0%;2012年共有植物8科19种,菊科和豆科占47.4%;2013年共有植物9科17种,菊科和豆科占35.3%。区域E2物种数量较少,各区域优势种(豆科和菊科)的优势地位随着封育年限的增加逐渐减弱,草原植被群落趋于稳定。区域E2植物群落多样性指数、均匀度指数均为3个区域中最低,而优势集中性指数以E2最高。各区域植物群落的多样性指数、均匀度指数和丰富度指数表现一致。主分量分析的结果显示影响核心区植物群落的主要环境因子为表层土壤速效P含量、0~10 cm和20~30 cm土壤含水量,影响边缘区植物群落的主要环境因子为表层土壤速效P、有机质含量以及0~10 cm和20~30 cm土壤含水量,而影响外围区植物群落的主要环境因子为表层土壤速效K含量、20~30 cm土壤含水量。极点排序图表明区域E各样方间相似性较高,植被群落单一。建议盐池县的封育年限以2~3 a为周期进行轮牧,以保持草地生态系统的平衡。
植物群落;多样性;主分量分析;极点排序;人工封育
人工封育是恢复退化草场的手段之一。封育作为退化草场恢复的一项重要措施,已被世界各国广泛采用(Meissner 和 Facelli,1999;Turner,1990)。目前国内外已经开展了许多有关封育措施的研究。Alice等在放牧区、围封禁牧区、围封放牧区3种区域条件下,对南美草原植物种类、组成群落的变化进行研究,结果表明在放牧条件下草地物种的丰富度和多样性高于围封禁牧区,地上净初级生产力比围封禁牧区高出50%以上;而围封放牧区的地上净初级生产力在3种区域中最高,比放牧区高出30%左右(Alice等,2005)。张克斌等以宁夏盐池为例,采用样线调查法进行结皮及植被调查,研究表明:北方半干旱区生物结皮对植物生长存在先促进后抑制的作用;在风蚀环境条件和不稳定的土壤条件下,生物结皮的出现可创造适合植物生长的微环境,为植物的生长提供有利条件,且有结皮现象的植被盖度明显高于无结皮现象的植被盖度(张克斌等,2008)。王燕等对在完全封育、人工栽培灌木的封育条件下生长的草地植物的优势度、科属组成及多样性的研究表明,在人工栽培灌木的封育区,植物种的丰富度及多年生草本植物种数都低于完全封育的天然植被区;天然植被区的植物组成结构复杂,抵御外界环境干扰能力较强;综合多样性指数表现为完全封育区的SW多样性指数、Patrick指数均显著高于人工栽培灌木的封育区(p<0.05),而其余各指数两者差异均不显著;与对照区相比,两种封育区生长的植物长势相对较好,且处于完全封育状态下的天然草地其自身的恢复能力相对较强(王燕等,2009)。郑敬刚等通过对经过10 a封育的草场的植物种类的组成及多样性、土壤含水量及有机质等的变换情况进行研究,结果表明围封增加了群落的植被盖度、多年生草本植物和旱生植物的种数,且物种的丰富度指数、多样性指数、均匀度指数均高于未封育区,同时封育区的土壤有机质、含氮总量也明显增加(郑敬刚等,2011)。本文通过对柳杨堡的植物群落特征及多样性进行分析,采用主分量分析法研究影响植物群落的环境因子,以及对各样地的植物群落进行极点排序,以期为宁夏盐池的草场管理提供合理的建议。
1 研究方法
1.1 样地设置
本研究依据当地主要土地利用类型和主要荒漠化治理工程种类,结合国家荒漠化定位监测项目,选择具有代表性的地段设置固定样地,采用GPS进行定位监测。研究区域位于盐池县的鄂尔多斯缓坡丘陵区毛乌素沙地西南缘的柳林堡乡人工封育区,距盐池县城20 km。采用如图1所示的处理方法对研究区样地进行划分:核心区(E)、边缘区(E1)、外围区(E2),其各区域理位置分别为:核心区,东经107°24′15″,北纬37°50′44″ ;边缘区,东经107°24′04″,北纬37°50′45″ ;外围区,东经107°23′47″,北纬37°50′47″。如表1所示,核心区始于1991年第1批全国防沙治沙试验示范区,区域周围有铁丝网围栏,可完全防止野生动物和家畜的采食,主要植物种有阿尔泰狗娃花(Heteropappus altaicus (Willd) Novopokr)、赖草(Leymus secalinus (Georgi) Tzvel.)、猪毛蒿(Artemisia scoparia Waldst. et Kit.)、刺沙蓬(Salsola ruthenica Iljin)等;边缘区从2002年以来采取围栏封育措施,主要植物种有黑沙蒿(Artemisia ordosica Krasch.)、刺沙蓬(Salsola ruthenica Iljin)、丝叶山苦荬(Ixeris chinensis)等;外围区从2002年以来虽也采取了封育,但还是受到了一定程度的人为干扰和放牧的影响,主要植物种有黑沙蒿(Artemisia ordosica Krasch.)、刺沙蓬(Salsola ruthenica Iljin)、苦豆子(Sophora alopecuroides L.)等(张克斌等,2004)。3个研究区域呈近距离直线分布,其自然条件基本一致。
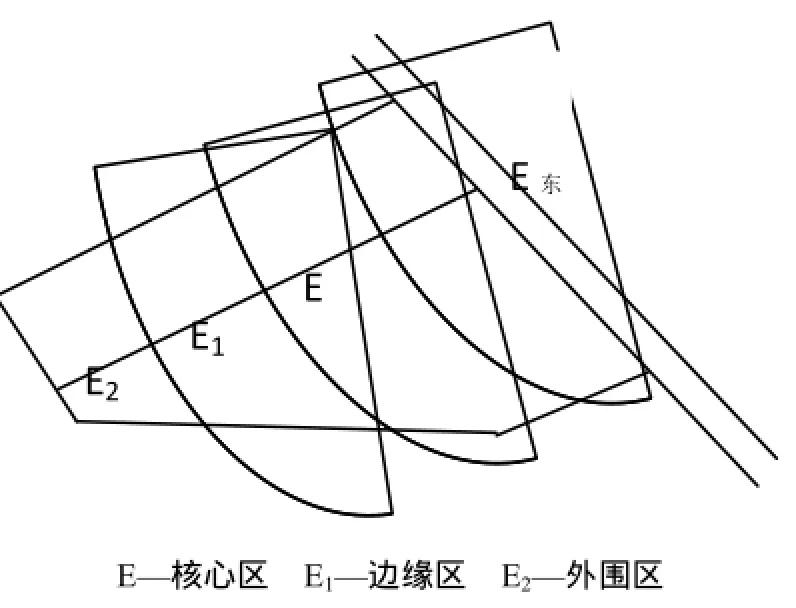
图1 研究区分布图Fig.1 The Distribution of Research Area
1.2 调查方法
研究调查时间为2006~2013年每年的7月份。具体调查方法为:从E开始沿着样带方向随机布设1 m×1 m的样方,E、E1、E2各布设10个样方,记为S1~S10,共30个样方。调查内容包括植物种名、株数、盖度、高度、生物量等。其中植物盖度采用针刺法,即用粗约为2 mm的细针从样方植物的上方随机垂直下插100次,记录触到植物体的次数,用百分数表示即为盖度。生物量测定是剪取植物的地上部分,用秤称得其鲜重重量。
1.3 数据处理
采用Excel2010和SPSS18.0统计分析软件进行处理。
1.3.1 多样性指数
植物群落的多样性选用物种多样性指数、群落优势集中性指数、群落均匀度指数和物种丰富度指数4种指标进行分析,其公式如下(韩争伟等,2013;王琳等,2004):
物种丰富度(S)和个体总数(N)在多样性分析中分别采用其自然对数(lnS和lnN)。
Shannon-Wiener多样性指数H′:
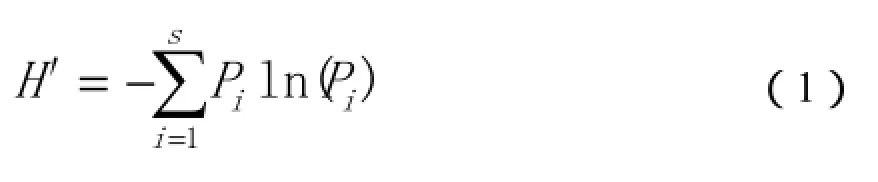
Simpson优势集中性指数C:

式(1)、(2)中,Pi为相对重要值,即Ni—第i种植物的重要值;N—样带植物重要值总和。
Berger-Parker优势度指数I:

式(3)中,Nmax—优势类群的数量;N—全部的种群数量。
Pielou均匀度指数J:

式(4)中,H′为Shannon—Wiener多样性指数;S为样带中物种数。
1.3.2 主分量分析与极点排序
以2013年的数据为样本,以优势种和非优势种的盖度、株数、重量、重要值为变量,进行主分量分析。极点排序采用Bray-Curtis相异系数法,群落间相异系数采用下式计算(Bray和Curtis,1957;淮虎银和侯扶江,1997;刘建军等,1996):
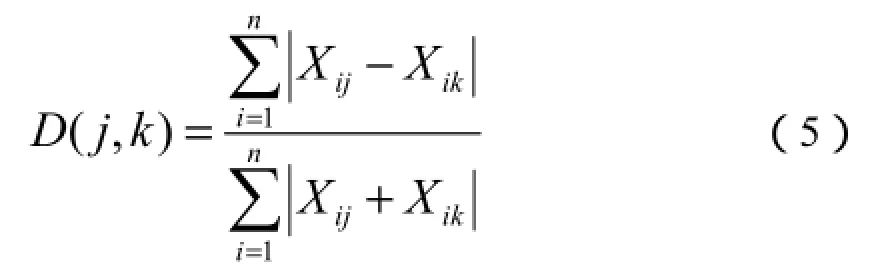
式(5)中:D(j,k)为群落j、k之间的相异系数;n为参加排序的群落类型数;Xij、Xik分别为植物种数i在群落j和k中的重要值(群落中某种植物出现的样方数占整个群落样方数的百分比)。
选择相异系数最大的2个群落作为x轴的2个端点,其他群落在x轴上的坐标根据下式计算:
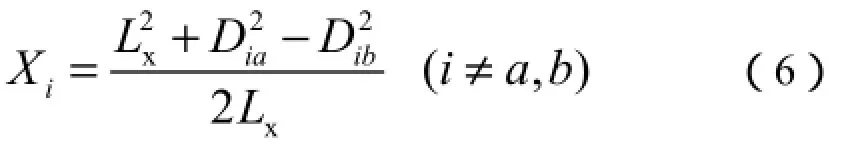
式(6)中:Lx为x轴两端点的距离;Dia和Dib分别为群落i与2个端点群落a、b的相异系数。
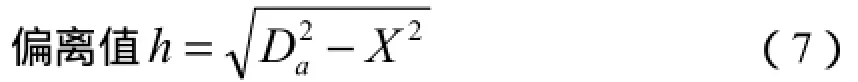
选取与x轴偏离值最大的群落为y轴的一个端点,另一个端点选取与前者(与x轴偏离值最大的群落)距离系数最大的群落,两端点确定后,其他点的坐标计算公式如下:
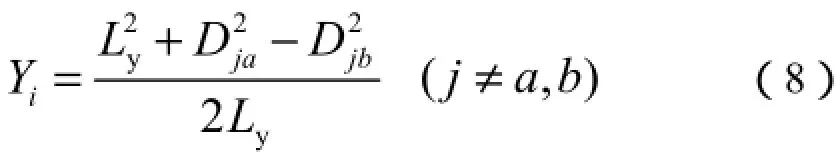
式中:Ly为y轴两端点的距离;Dja和Djb分别为群落j与2个端点群落a、b的相异系数。
2 结果与分析
2.1 柳杨堡植被群落特征
2011年样地E中共有植物9科21种,其中:菊科6种,占28.57%;豆科5种,占23.81%;禾本科3种,占14.29%;藜科2种,占9.52%;柽柳科、苋科、大戟科、萝藦科和紫葳科各1种,各占4.76%。2012年共有植物7科20种,其中:菊科和豆科各5种,各占25%;禾本科3种,占15%;藜科、苋科和大戟科各2种,各占10%;蓼科1种,占5%。2013年共有植物9科22种,其中:菊科7种,占31.82%;禾本科5种,占22.73%;豆科4种,占18.18%;藜科、苋科、大戟科、紫葳科、百合科和旋花科各1种,各占4.55%。
2011年样地E1种共有植物8科18种,其中:豆科6种,占33.33%;菊科4种,占22.22%;禾本科3种,占16.67%;芸香科、藜科、苋科、大戟科和萝藦科各1种,各占5.56%。2012年共有植物7科16种,其中:菊科5种,占31.25%;豆科3种,占18.75%;禾本科、藜科和苋科各2种,各占12.50%;大戟科和萝藦科各1种,各站6.25%。2013年共有植物9科16种,其中:菊科和豆科各4种,各占25%;禾本科2种,占12.25%;芸香科、藜科、苋科、大戟科和旋花科各1种,各占6.25%。
2011年样地E2中共有植物6科15种,其中:菊科5种,占33.33%;豆科4种,占26.67%;禾本科和藜科各2种,各占13.33%;苋科和大戟科各1种,各占6.67%。2012年共有植物8科19种,其中:菊科6种,占31.58%;禾本科4种,占21.05%;豆科3种,占15.79%;苋科2种,占10.53%;藜科、大戟科、萝藦科和紫葳科各1种,各占5.26%。2013年共有植物9科17种,其中:菊科和禾本科各4种,各占23.53%;豆科和藜科各2种,各占11.76%;芸香科、苋科、大戟科、萝藦科和紫葳科各1种,各占5.88%。
由上文可知,样地E2的平均物种数低于样地E和样地E1,这是由于样地E2存在一定程度上的动物采食和践踏,动物行为导致了群落中某些植物的消失,从而使群落物种数量下降。从2011年至2013年,3个区域的群落伴生物种逐渐增多,豆科和菊科优势种的优势地位逐渐减弱,这表明随着植物群落演替的进行,草原逐渐趋于稳定,环境适应能力趋于更强。由表2中可知,2011—2013年,区域E2中黑沙蒿均具有明显的优势,其重要值分别占整块样地的35.54%,45.72%和32.50%;而区域E与区域E1无特定的优势种。2011年,区域E中植物物种以阿尔泰狗娃花和狗尾草为主,2012以阿尔泰狗娃花和黑沙蒿为主,2013年以角蒿和刺沙蓬为主。2011年,区域E1中植物物种以黑沙蒿和刺沙蓬为主,2012年以猪毛蒿和赖草为主,2013年以阿尔泰狗娃花和黑沙蒿为主。3年来,区域E和区域E1优势种的变化无明显规律。
2.2 植物群落多样性特征指数时间动态
从图2中可以看出,各区域多样性指数随封育年限的增加呈上升的总趋势,这说明随着封育年限的延长,植物群落越来越复杂,物种多样性越来越丰富。其中,区域E2的多样性指数低于其他2个区
域,而区域E与E1多样性指数接近。3个区域群落优势集中性指数变化趋势与多样性指数变化趋势相反。区域E1的优势集中性指数曲线低于其他两个区域,并且变化平缓,而其他两个区域的优势集中性指数变化幅度较大。均匀度指数反应的是群落中物种分布的均匀程度。3个区域中均匀度最低的是以黑沙蒿为单优势种的E2区域。区域E1均匀度曲线波动小,而E与E2变化幅度较大。此外,从图2可知,均匀度指数与多样性指数变化趋势基本一致。物种丰富度表示群落中物种数量的多少,3个区域物种丰富度指数均随封育年限的增加呈现上升的总趋势,其中区域E2的平均丰富度指数低于其他2个区域。3个区域丰富度指数波动各异,可能原因是由于各区域的封育年限、人为干扰程度等不尽相同。
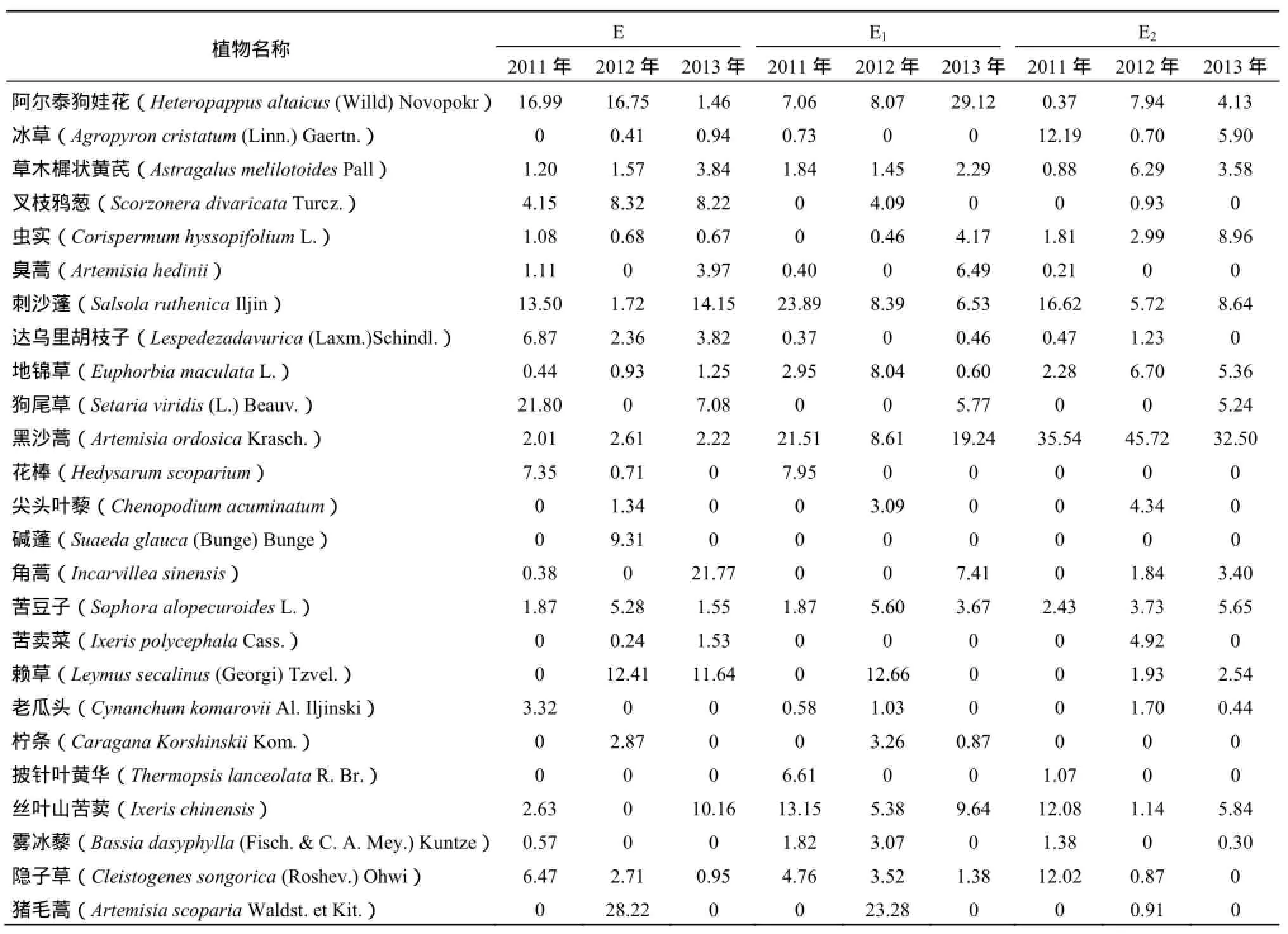
表2 各样地不同年份主要植物的重要值Table 2 The important value of main plant for different year of each sample
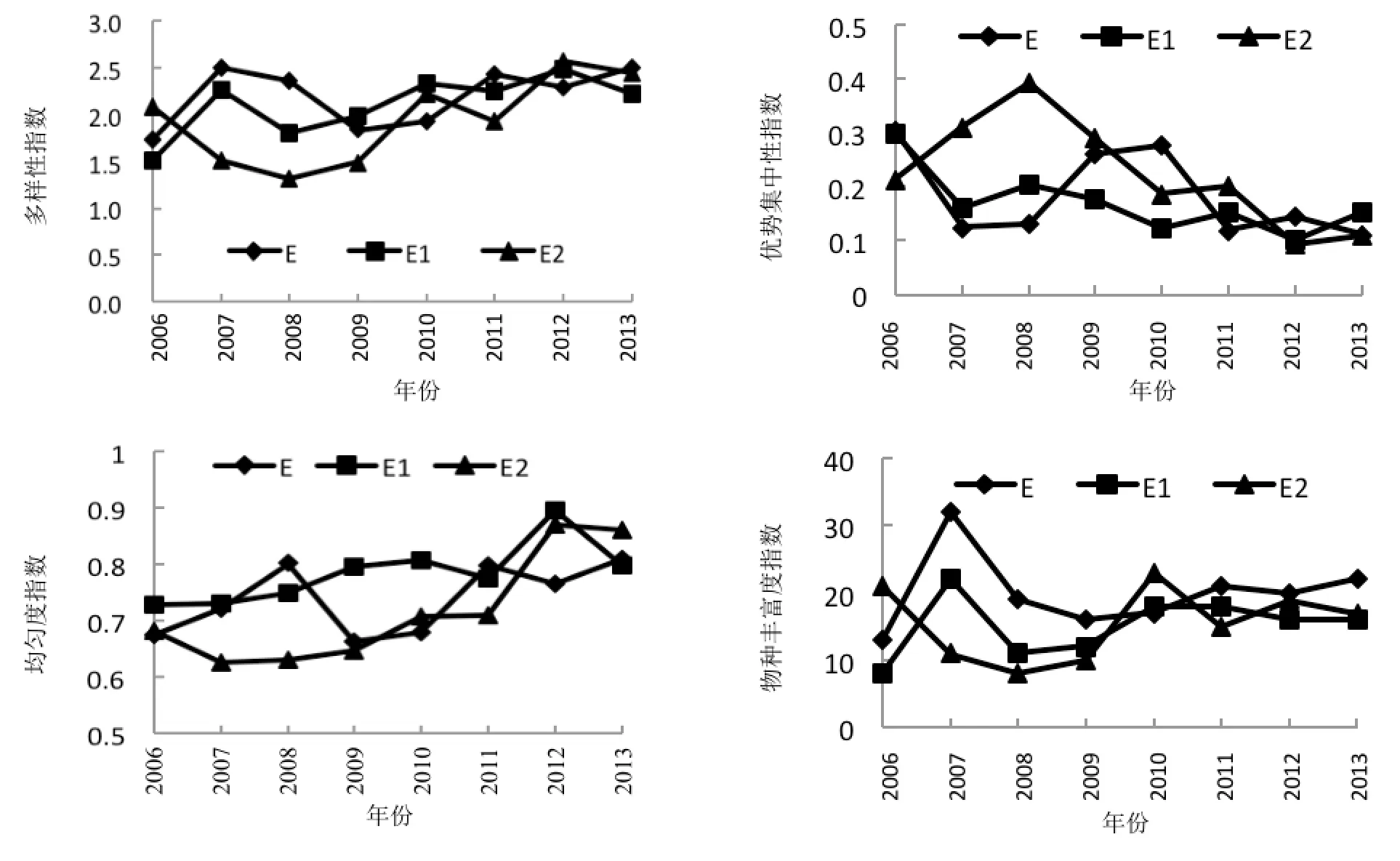
图2 各样地植物群落多样性指数(H′)、均匀度指数(J)、物种丰富度指数(R)和优势集中性指数(C)的时间动态Fig.2 The time-pattern tendency of diversity index (H′), evenness index (J), specific richness (R) and dominant concentration index (C) of plant communities in different samples
植物群落的丰富度指数、均匀度指数与多样性指数关系密切。如表3所示,本文各研究区域植物群落的丰富度指数与多样性指数均呈显著正相关关系(p<0.05),其中区域E的丰富度指数与多样性指数呈显著正相关关系(p<0.01)。区域E和区域E2植物群落的均匀度指数与多样性指数分别呈显著正相关关系(p<0.05)和极显著正相关关系(p<0.01)。
2.3 植物群落的主分量分析
为了获知影响柳杨堡植物群落的主导因素,本文分别对各研究区域的植物群落进行主分量分析,各因子的特征向量见表4。
从表4可知,3个区域前3个主分量的累计贡献率均超过85%。核心区(E)的第一主分量代表了表层土壤速效P含量和0~10 cm土壤含水量,累计贡献率为44.67%,第二主分量代表了20~30 cm土壤含水量,两个主分量累计贡献率达86.91%,表明核心区表层土壤速效P含量、0~10 cm与20~30 cm土壤含水量是影响植物群落的主要因子。
边缘区(E1)植物群落的第一主分量代表了表层土壤有机质含量,第二主分量代表了0~10 cm土壤含水量,第三主分量代表了表层土壤速效P含量和20~30 cm土壤含水量,3个主分量的累计贡献率达91.67%。其中,第一主分量贡献率为33.29%,第二主分量贡献率为32.03%,第三主分量的贡献率为26.35%,三个主分量贡献率近似,表明边缘区植物群落主要受表层土壤速效P和有机质含量、0~10 cm和20~30 cm土壤含水量的影响。
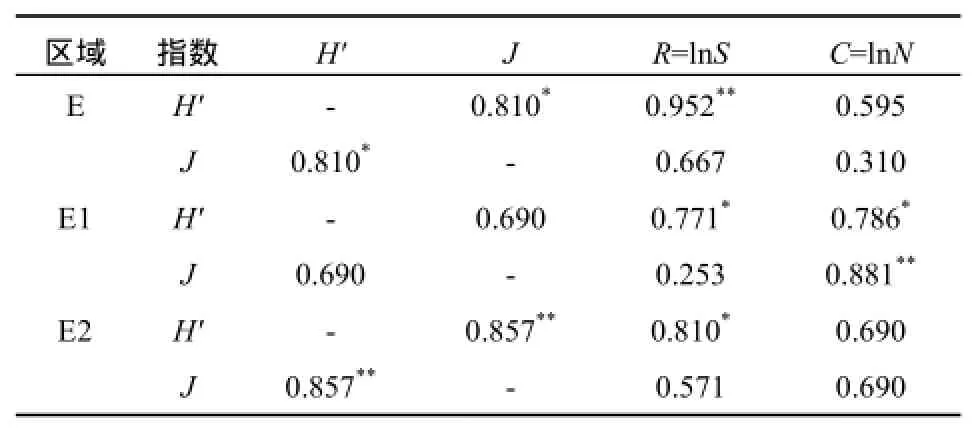
表3 多样性特种指数间的相关系数Table 3 The correlation coefficients of indices of diversity
外围区(E2)植物群落的第一主分量代表了表层土壤速效K含量和20~30 cm土壤含水量,第二主分量代表了表层土壤有机质含量,累计贡献率达86.05%,说明表层土壤速效K和有机质含量,以及20~30 cm土壤含水量主导了外围区植物群落的变化。
根据以上主分量分析的结果可知,柳杨堡人工封育核心区、边缘区和外围区植物群落变化的影响因子主要集中在20~30 cm土壤含水量,究其原因,可能是由于该土层的水分不易蒸发流失,且植物根系吸收的水分主要来自土壤20~30 cm深度。再者,已有研究表明在干旱-半干旱区水分是影响植物群落的主要因子(张克斌等,2004)。
2.4 植物群落极点排序
本研究采用极点排序法,利用植物群落的相异性系数(表5),计算出各植物群落的相关数值,并对其进行排序。以区域E为例,各样方植物群落的相异性系数如表所示,排序指标如表6所示。
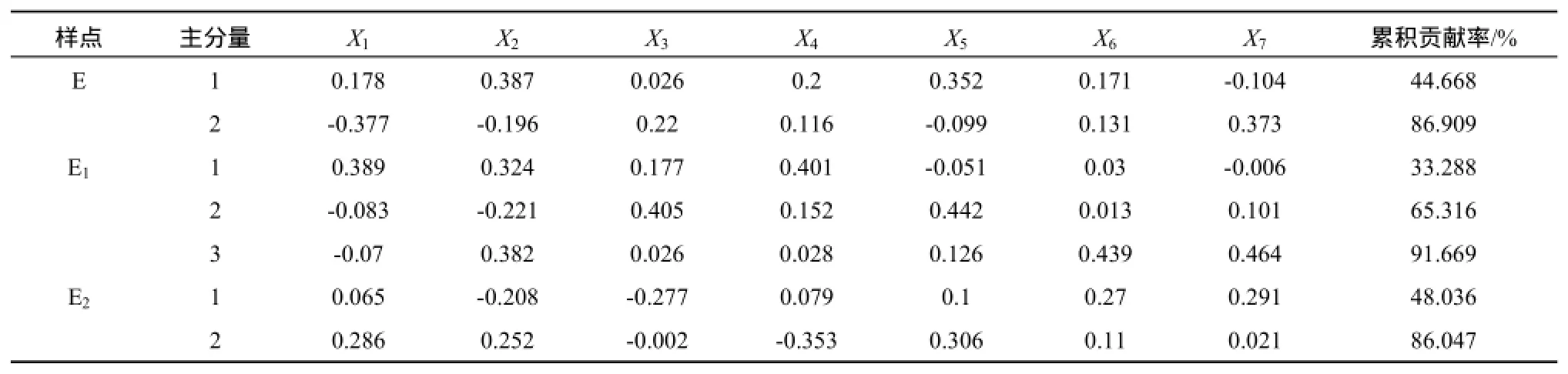
表4 各样点植物群落的主分量分析Table 4 Principal Component Analysis of the Plant Communities
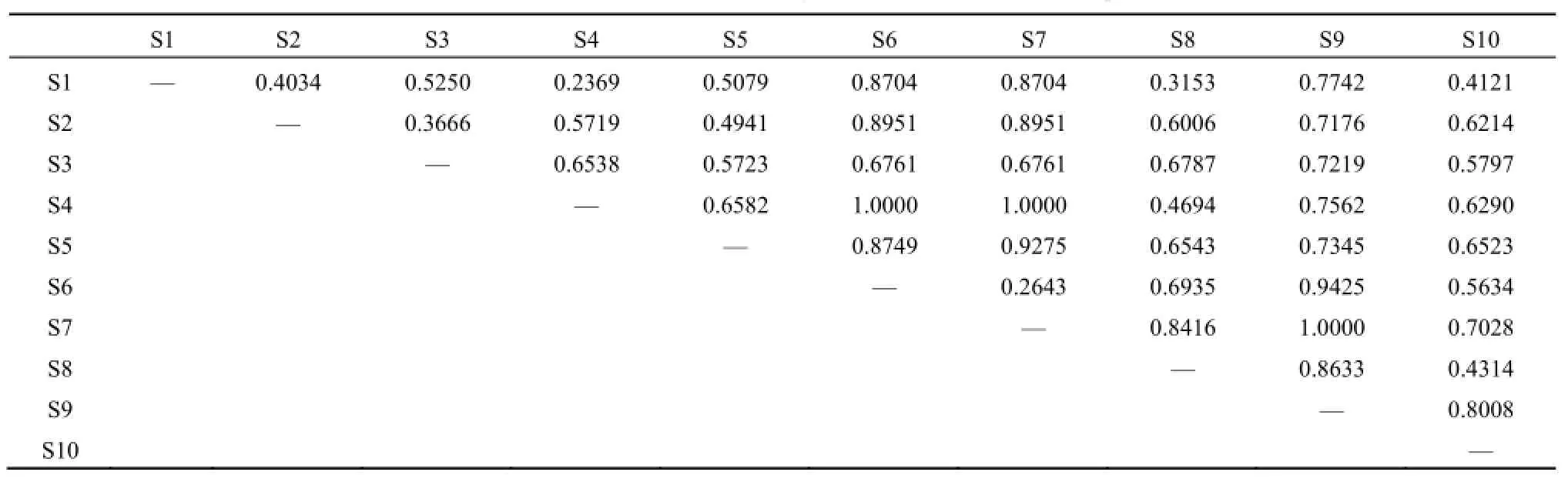
表5 样地E各样方群落的相异性系数Table 5 The Coefficient of Dissimilarity of Plant Communities in Sample E

表6 样地E各样方植物群落排序指标值Table 6 The Indices of Ordination of Plant Communities in Sample E

图3 各样点植物群落极点排序Fig.3 Polar Ordination of Plant Communities
从图3可知,区域E各样方在极点排序图中分布相对集中,而E1、E2各样方在极点排序图中分布相对分散,这说明区域E各样方间植物群落的相似性较区域E1与E2的高。这是由于样地E处于完全封育状态,未受到人为和放牧的干扰,且封育时间较长,植物群落模式趋向单一。由此说明,长期的封育会使植物群落多样性弱化,不利于草场植被的恢复与管理。
3 讨论与结论
3.1 讨论
柳杨堡人工封育轻度干扰区(区域E2)的平均物种数低于无干扰区(区域E与区域E1),这是由于人为活动及放牧致使区域内某些物种消失。在时间尺度上,3个区域优势种的地位逐渐减弱,此现象有利于草场的恢复,使草场的环境适应能力更强。
随着封育年限的增加,各区域植物群落多样性指数呈上升的总趋势;区域E2由于形成了以黑沙蒿为优势种的群落,其优势集中性指数高于其余2个区域;同时,区域E2植物群落的多样性指数、均匀度指数均低于区域E与区域E1。柳杨堡各样地植物群落的多样性指数与丰富度指数、均匀度指数均呈显著正相关,说明这3个指标均可有效反映柳杨堡地区植物群落的特征。
柳杨堡人工封育核心区(E)植物群落主要受表层土壤速效P含量以及0~10 cm和20~30 cm土壤含水量3个环境因子的影响,边缘区(E1)植物群落主要受表层土壤速效P和有机质含量以及0~10 cm和20~30 cm土壤含水量4个环境因子的影响,而外围区(E2)植物群落主要受表层土壤速效K含量以及20~30 cm土壤含水量2个环境因子的影响。柳杨堡属于干旱—半干旱的气候环境,因此土壤含水量成为影响植物群落的主要环境因子。本研究认为20~30 cm处的土壤含水量,是影响封育区植物群落的主要环境因子。
在极点排序图上,区域E各样方分布较E1与E2集中,因为区域E处于完全封育状态,不受人为干扰和放牧影响,且封育时间长,植物群落单一相似。由此说明,长期的完全封育不利于植物群落的演替。根据草地生态系统的可持续性原则,草地围封不应是无限期的,封育期过长,不但不利于牧草的正常生长和发育,而且枯草会抑制植物的再生和幼苗的形成,不利于草地的繁殖更新。因此,草地围封一段时间后,应进行适当利用,使草地生态系统的能量流动和物质循环与外界保持平衡,进而保持草地生态系统平衡。相关研究表明,适当的放牧有利于保持草地群落较高的生物多样性(祁彪等,2005;Collins,1987)。然而,封育时间的长短,应根据草地退化程度和草地恢复状况而定(郑翠玲等,2005;程积民等,1995;程积民和邹厚远,1998)。就盐池而言,建议以2~3 a为周期进行轮牧,更有利于草场的恢复。
3.2 结论
宁夏盐池人工封育区植物群落组成以豆科和菊科为主,其中外围区(样地E2)在封育过程中形成了以黑沙蒿为优势种的群落结构。柳杨堡人工封育区的植物群落主要受20~30 cm土壤含水量的影响。随着封育年限的增加,各区域优势种的地位逐渐减弱,物种多样性逐渐提高,但长期完全封育区(样地E)使得植物群落单一,不利于草场恢复。因此,建议杨柳堡以2~3 a为周期进行适当放牧,以保持草地生态系统的平衡。
ALICE A, MARTIN O, ELSA L, et al. 2005. Effect of grazing on community structure and productivity of a Uruguayan grassland[J]. Plant Ecology, 179:83-91.
BRAY J R, CURTIS J T. 1957. An ordination of the upland forest communities of Southern Wisconsin. Ecological Monographs, 27:325-349.
COLLINS S L. 1987. Interaction of disturbance in tall grass prairie: a field experiment[J].Ecology, 68(5):1243-1250.
MEISSNER R A, FACELLI J M. 1999. Effects of sheep exclusion on the soil seed bank and annual vegetation in chenopods shrub lands of south Australia[J]. Journal of Arid Environments, 42: 117-128.
TURNER R M. 1990. Long-term vegetation change at a fully protected Sonoran desert site[J]. Ecology, 7:464-477.
程积民, 邹厚远, 本江昭夫. 1995. 黄土高原草地合理利用与草地植被演替过程的试验研[J]. 草业学报, 4(4):17-22.
程积民, 邹厚远. 1998. 封育刈割放牧对草地植被的影响[J]. 水土保持研究, 5(1):36-54.
韩争伟,马玲,曹传旺,等. 2013. 太湖湿地昆虫群落结构及多样性[J]. 生态学报, 33(14):4387-4397.
淮虎银, 侯扶江. 1997. 青海湖湖盆南部植物群落二维极点排序[J]. 草业学报, 6(2):19-21.
刘建军, 雷瑞德, 尚廉斌, 等. 1996. 火地塘林区原生森林植被极点排序及分类[J]. 西北林学院学报, 11(增):37-42
祁彪, 张德罡, 丁玲玲, 等. 2005. 退化高寒干旱草地植物群落多样性特征[J]. 甘肃农业大学学报, 40(5):626-631.
王琳, 张金屯, 上官铁梁, 等. 2004. 历山山地草甸的物种多样性及其与土壤理化性质的关系[J]. 应用与环境生物学报, 10(1):18-22.
王燕, 王辉, 李永兵, 等. 2009. 干旱荒漠区封育沙地植被群落特征变化研究[J]. 水土保持通报, 29(5):40-44.
张克斌, 李瑞, 侯瑞萍, 等. 2004. 宁夏盐池县不同荒漠化治理措施植物多样性研究[J].中国水土保持科学, 2(4):66-72.
张克斌, 卢晓杰, 李瑞. 2008. 北方农牧交错带沙地生物结皮研究[J]. 干旱区资源与环境, 22(4):147-151.
郑翠玲, 曹子龙, 王贤, 等. 2005. 围栏封育在呼伦贝尔沙化草地植被恢复中的作用[J]. 中国水土保持科学, 3(3):78-81.
郑敬刚, 何明珠, 苏云, 等. 2011. 放牧和围封对干旱区草地生态系统的影响[J]. 河南农业科学, (12):80-83.
Structure and diversity of plant communities after cultivation and recovery in Yanchi county, Ningxia
HAO Zhiru,ZHANG Kebin
Key Laboratory of Soil and Water Conservation and Desertification Combating of the the Ministry of Education, Beijing Forestry University, Beijing 100083, China
In order to analyze the structure and variation of plant communities in Yanchi county, Ningxia, a population of plants emerging after cultivation was surveyed and the studied area was divided into three areas:core area (E), edge area (E1) and peripheral area (E2). Samples of the plant population were recorded according to species, abundance, coverage, height, and biomass. They were analyzed in respect to diversity indices (species diversity index, community dominant concentration index, community evenness index and species richness index), principal component analysis and polar ordination. The results showed that the dominant species after cultivation and recovery in Yanchi County, Ningxia are from the genera compositae and leguminosae. Four random samples each of 1m2were examined. Sample area E contained 9 families and 21 species in 2011, 7 families and 20 species in 2012, and 9 families and 22 species in 2013. The genera compositae and leguminosae accounted for 52.4% of the total population in 2011, and for 50.0% in 2012 and in 2013. Sample area E1 had 8 families and 18 species in 2011, 7 families and 16 species in 2012, and 9 families and 16 species in 2013. Compositae and leguminosae accounted for 55.6% in 2011, and for 50.0% in 2012 and in 2013. Sample area E2 had 6 families and 15 species in 2011, 8 families and 19 species in 2012, and 9 families and 17 species in 2013. Compositae and leguminosae accounted for 60.0% in 2011, for 47.4% in 2012 and for 35.3% in 2013. The number of species of sample area E2 was the lowest, and the dominant position of species in the genera (leguminosae and compositae) in all kinds of sample areas were gradually weakened, and the grassland vegetation community tended to be stable. The diversity index, and evenness index of E2was the lowest, and the advantage of concentration index was the highest. In all sample areas, the diversity index was significantly correlated with the evenness index and the richness index. The results of principal component analysis showed that the main environmental factors affecting the sample E were content of rapidly available P in soil surface, and soil moisture content at the depths of 0~10 cm and 20~30 cm. For the sample E1the factors were content of rapidly available P and organic matter in the soil surface, and soil moisture content at depths of 0~10 cm and 20~30 cm. For the sample E2the main environmental factors affecting the population were content of rapidly available K in the soil surface, and soil moisture content from 20~30 cm. The figures of polar ordination showed that the quadrats forming sample E were the most similar and the population of plants was relatively single. Therefore, it was concluded that the best time to cultivate in Yanchi is 2~3 years. And with 2~3 years as a cycle of rotational grazing can better promote the grassland restoration so that it can be kept the balance of the grassland ecosystem.
plant community; diversity; principal component analysis; polar ordination; artificial cultivating
S718.54
A
1674-5906(2014)03-0377-08
郝智如,张克斌. 宁夏盐池人工封育区植物群落结构及多样性[J]. 生态环境学报, 2014, 23(3): 377-384.
HAO Zhiru,ZHANG Kebin. Structure and diversity of plant communities after cultivation and recovery in Yanchi county, Ningxia [J]. Ecology and Environmental Sciences, 2014, 23(3): 377-384.
国家自然科学基金项目(30771764);国家林业局荒漠化定位监测项目(660550)
郝智如(1989年生),女,硕士研究生,研究方向为水土保持与荒漠化防治。E-mail: haozhiru891121@126.com
*通讯作者:张克斌(1957年生),男,教授,博士生导师,研究方向为荒漠化监测与防治。E-mail:ctccd@126.com
2014-02-09
