厚荚相思和尾叶桉人工林土壤线虫群落研究
2014-02-07赵灿灿
赵灿灿,王 伟
河南大学生命科学学院,河南 开封 475004
ZHAO Cancan*, WANG Wei
School of Life Science, Henan University, Kaifeng 475004, China
厚荚相思和尾叶桉人工林土壤线虫群落研究
赵灿灿*,王 伟
河南大学生命科学学院,河南 开封 475004
相思树和桉树等树种因其生长迅速、耐性好以及适合制作纸浆等特点,近年来在华南地区大面积种植,带来了巨大的经济利益,但是相思树和桉树如何影响林下土壤生物多样性和土壤肥力还不清楚,科学评价其林下土壤健康水平十分必要。土壤线虫的种类和数量十分丰富,在土壤食物网各营养级中占据重要位置,对土壤环境变化反应敏感,作为土壤健康的指示生物被广泛应用。本文以幼龄厚荚相思Acacia crassicapa和尾叶桉Eucalyptus urophylla人工林为研究对象,于干季和湿季分别对2个林分根区和非根区土壤线虫进行研究,探讨土壤线虫这种指示生物的数量、各营养类群比例和多样性的变化规律,以及它们和环境因子的关系。主要结果表明:(1)厚荚相思土壤线虫总数为每公斤干土6 741条,显著高于尾叶桉线虫总数的28.3%。厚荚相思人工林土壤食细菌线虫的百分比相比尾叶桉人工林高6.3%,且差异显著。(2)土壤线虫的群落组成变化具有明显的季节波动,湿季土壤食细菌线虫比例上升,而植物寄生线虫的比例下降,多样性指数由干季的0.87减小到0.75,统计差异均达到了显著水平。(3)根区比非根区显著提高了线虫数量的89.1%,这是由根区积累更多的有机碳和总氮引起的。总之,厚荚相思人工林为土壤自由生活线虫提供了优良环境,且在幼龄期没有表现出土壤酸化,生态效应优于尾叶桉人工林,在华南地区人工林营林过程中可适当增加厚荚相思林分面积。
指示生物;营养类群;外来物种;根际;生态效应
相思树为豆科植物,具有根瘤菌,有固氮作用,原产于澳大利亚。1985年中国林业科学研究院最早引入相思树,并开始在华南地区大面积种植(潘志刚和林鸿盛,1996)。厚荚相思Acacia crassicapa的高、径生长最快,年均高生长量达1.9~2.2 m,年均径粗生长达2.2~2.5 cm;并以其特有的耐寒性和耐盐渍,可以在水土严重流失的地区和滨海地区立地,也能迅速成林成材,干形较直且木材硬度适中,在广东作为红木家具开发,也可制较高强度的书写纸和印刷纸(肖石海等,1996)。桉树为桃金娘科植物,原产于澳大利亚,全世界有近百个国家将桉树引种栽培。我国于1890年将桉树引入,目前桉树人工林主要分布在广东、广西、海南、福建等省区,栽培面积已达150多万hm2,居世界第3位(林婷,2012)。桉树是世界上生长速度最快、经济价值最大的三大树种(松、桉、杨)之一,生长旺季时1天可长高3~4 cm,尤其适合生产短纤维纸浆,被认为是纸浆工业的“绿色黄金”。尾叶桉Eucalyptus urophylla是我国实施速生丰产用材林基地建设的首选树种之一,目前在华南地区大面积推广。相思树和桉树给人类带来了巨大的经济效益,但也引发了一系列生态上的争议,例如相思树存在着致酸效应,氢离子的释放来自于豆科植物依靠根固氮,通过固氮从土壤溶液中吸收的阳离子比阴离子多,植物总的过量阳离子含量增加导致土壤酸化,近一步影响土壤动物和微生物(Tang和Yu,1999;刘菊秀等,2000);桉树过度消耗水分,存在较强的化感作用,通过分泌特殊的化学物质,对其他植物、林下植被及土壤中的动物和微生物产生抑制作用,降低生物多样性(Shiva和Bandyopadhyay,1983;Poore和Fries,1985;廖崇惠,1990)。因此在我国华南地区厚荚相思和尾叶桉人工林种植面积逐年增加的背景下,评价其林下土壤健康水平十分必要。
土壤动物如线虫等是土壤生物的重要组成部分,在土壤有机质分解、养分循环和改善土壤结构中具有重要作用。土壤线虫因其种类和数量丰富、分布广泛、对生境变化反应灵敏等特点在土壤食物网中扮演重要角色,作为土壤健康的指示生物已成为生态学研究的重要内容(李玉娟等,2005;邵元虎等,2007)。本文通过比较不同季节厚荚相思和尾叶桉根区和非根区土壤线虫群落结构和多样性,研究厚荚相思和尾叶桉纯林在种植初期对土壤线虫的影响,从而为评价这2种林型的生态效应、指导人工林树种的选择以及促进土壤生态系统健康发展提供科学依据。
1 材料与方法
1.1 试验地概况
研究地点位于广东省鹤山市中国科学院鹤山森林生态系统国家野外科学观测研究站共和试验地(22°41′N, 112°54′E),地处南亚热带丘陵地区,平均海拔80 m。该地区年平均温度22.6 ℃,7月平均温度28.7 ℃,1月平均温度14.5 ℃,极端最高温度37.5 ℃,极端最低温度0 ℃,年平均≥10 ℃的有效积温为7597.2 ℃;年平均降水量为1700 mm,年平均蒸发量为1600 mm;地带性土壤为赤红壤,成土母质是砂页岩。树种种植前的本底调查结果显示各样方之间没有显著差异。厚荚相思和尾叶桉纯林于2005年种植,树间距为3 m× 2 m,每种纯林有3个重复,立地条件相对一致,林下优势植被为芒箕Dicranopteris dichotoma。
1.2 取样方法
于2006年11月(干季)和2007年5月(湿季)分别在厚荚相思和尾叶桉人工纯林进行取样,2种林型分别有3个重复林分。取样时考虑到地形坡度的影响,每一林型样方分别取上坡、中坡和下坡,每一取样坡度分根区和非根区采集土壤样品。采用内径为5 cm的土钻取样,每个土壤样品为3个钻混合,拣出可见的植物根系和石块并过2 mm筛。土壤样品标记好后,带回实验室4 ℃保存,用于土壤线虫的提取和土壤性质的测定。
1.3 测定方法
土壤含水量的测定采用烘干称重法,土壤总有机碳(TOC)的测定采用重铬酸钾氧化滴定法,土壤总氮(TN)的测定采用半微量凯氏定氮法,土壤pH值的测定采用去离子水浸提(体积比为1:2.5)pH计测定法。
土壤线虫的提取采用贝尔曼湿漏斗法(McSorley,1987)。贝尔曼湿漏斗法的原理是线虫没有主动的游泳能力,在浸水的条件下,土壤中的线虫会由于重力作用而沉在漏斗底部。将大约50 g新鲜土壤置于漏斗中浸水,48 h后打开下方的止水夹,收集到线虫溶液。土壤线虫标本用热福尔马林溶液固定,终浓度为4%,并用倒置显微镜(TS100, Nikon, Japan)进行计数和营养类群鉴定。如果线虫总数超过100条,则仅鉴定100条来代表各营养类群比例。线虫的营养类群包括4类,分别为食细菌线虫、食真菌线虫、植物寄生性线虫和捕食、杂食性线虫(Yeates等,1993)。香农威尔多样性指数H'的计算公式为H'= -∑ Pi(lnPi),式中Pi为第i个分类单元中个体所占的比例。
1.4 统计分析
不同林型、不同根区、不同季节和不同坡度之间土壤线虫和环境因子的差异采用多因素方差分析(SPSS 13.0 for Windows, SPSS Inc., USA),平均值之间显著性水平用LSD法检测,土壤线虫和环境因子的相关性检测采用回归分析方法。
2 实验结果
2.1 土壤性质
多因素方差分析结果表明林型对土壤含水量有显著影响(表1),厚荚相思林型土壤含水量为0.173,远高于尾叶桉林型土壤含水量0.141。采样季节对土壤含水量有极其显著的影响,湿季的含水量为0.180,远远高于干季含水量0.134。不同的采样坡度也显著影响着土壤含水量。根区土壤总有机碳显著高于非根区,厚荚相思土壤总有机碳为18.01 g·kg-1,高于尾叶桉(16.36 g·kg-1),但差异不显著。土壤总氮含量结果与有机碳相似,根区土壤总氮显著高于非根区,厚荚相思土壤总氮为1.04 g·kg-1,高于尾叶桉(0.98 g·kg-1),但差异不显著。林型、根区、季节和坡度的主效应均对土壤碳氮比(C/N)没有显著影响。季节对土壤pH值影响显著,湿季pH值小于干季,酸化更明显。坡度的主效应对总有机碳、总氮、碳氮比和pH均无显著影响。林型和根区的交互作用、林型和季节的交互作用以及根区和坡度的交互作用改变了碳氮比,效果显著。
2.2 土壤线虫群落
平均土壤线虫总数为每公斤干土5750 条。不同采样季节对线虫总数没有显著影响。厚荚相思土壤线虫总数为每公斤干土6741 条,高于尾叶桉线虫总数28.3%,差异显著(P<0.05)(表2,图1)。根区土壤线虫总数为7568条,高于非根区线虫总数53.9%,差异极显著(P<0.001,表2)。坡度对线虫总数没有显著影响。季节和林型的交互作用极显著地影响土壤线虫总数(P<0.01,表2)。在干季厚荚相思林下线虫总数为每公斤干土5201 条,尾叶桉林下线虫总数为每公斤干土5963条,差异不显著。在湿季厚荚相思林下线虫总数为每公斤干土8281条,尾叶桉林下线虫总数为每公斤干土3698条,极显著地低于厚荚相思(P<0.001)。

表1 土壤性质各处理的平均值和多因素方差分析Table1 Means and ANOVA of soil properties
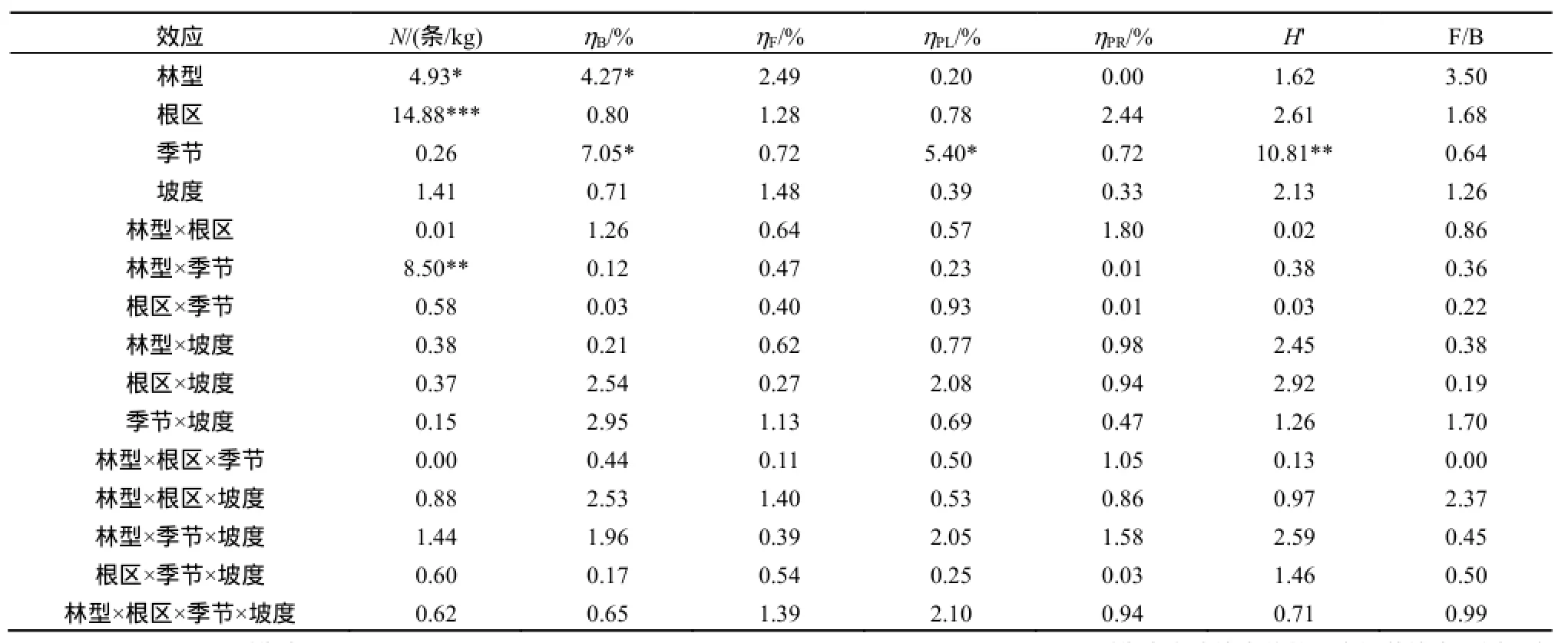
表2 土壤线虫各指标方差分析Table2 ANOVA of soil nematode index
食细菌线虫和食真菌线虫在各个处理中均为优势种群(图2)。不同采样季节对食细菌线虫和植物寄生性线虫百分比有显著的影响(P<0.05),不同林型对食细菌线虫百分比有显著的影响(P<0.05),根区和坡度对各营养类群百分比均没有显著的影响(表2)。干季:厚荚相思土壤中,食细菌线虫、食真菌线虫、植物寄生性线虫和捕食、杂食性线虫所占的百分比分别为64.6%、22.3%、10.5%和2.6%;尾叶桉土壤中,食细菌线虫、食真菌线虫、植物寄生性线虫和捕食、杂食性线虫所占的百分比分别为59.4%、25.0%、13.1%和2.6%。湿季:厚荚相思土壤中,食细菌线虫、食真菌线虫、植物寄生性线虫和捕食、杂食性线虫所占的百分比分别为73.5%、17.5%、5.8%和3.2%;尾叶桉土壤中,食细菌线虫、食真菌线虫、植物寄生性线虫和捕食、杂食性线虫所占的百分比分别为66.1%、23.9%、6.7%和3.4%。厚荚相思林型土壤食真菌线虫与食细菌线虫的比值为0.33(图3),低于尾叶桉食真菌线虫与食细菌线虫的比值0.44,非根际食真菌线虫与食细菌线虫的比值为0.34,低于根际食真菌线虫与食细菌线虫的比值0.43,但差异都不显著(表2)。
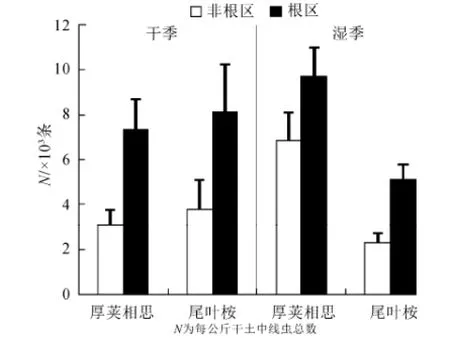
图1 不同季节厚荚相思和尾叶桉林型土壤线虫总数Fig.1 Total soil nematode individuals in Acacia crassicapa and Eucalyptus urophylla in different seasons
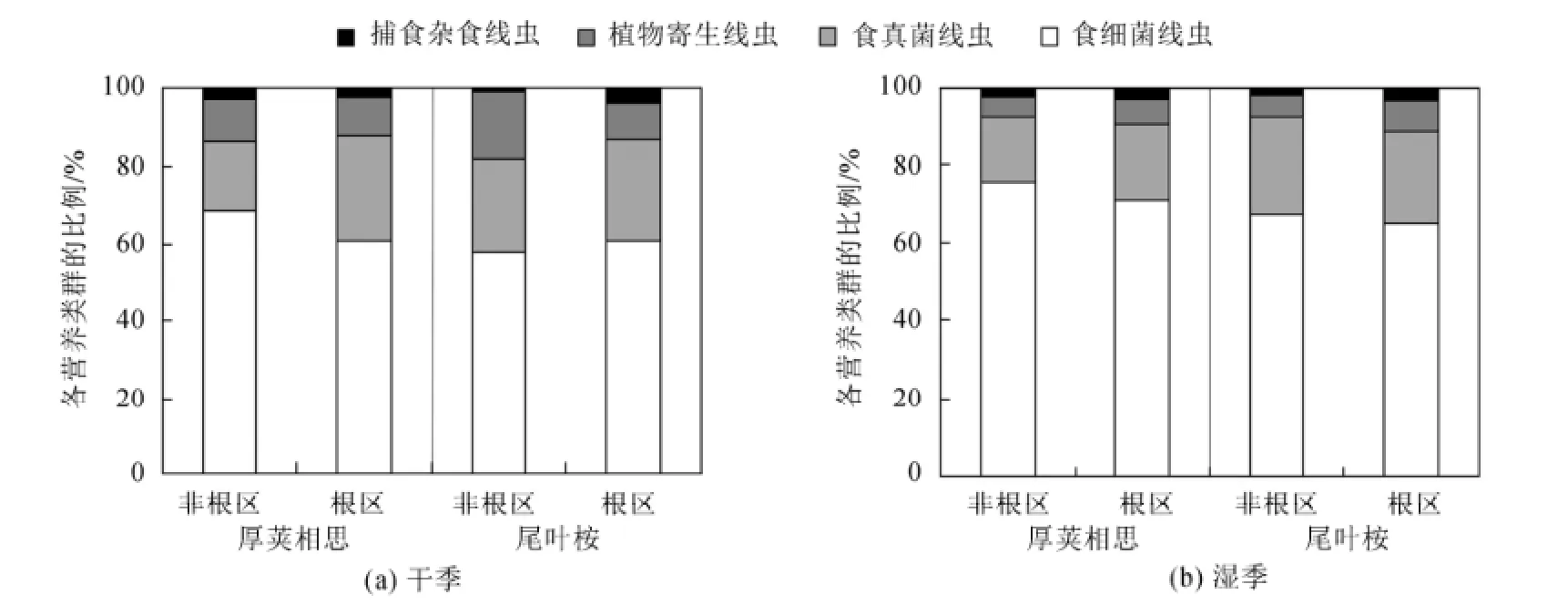
图2 不同季节厚荚相思和尾叶桉林型土壤线虫各营养类群的百分比Fig.2 Percentage of nematode trophic groups in Acacia crassicapa and Eucalyptus urophylla in different seasons

图3 不同季节厚荚相思和尾叶桉林型食真菌线虫与食细菌线虫的比值Fig.3 Ratio of fungivores to bacterivores in Acacia crassicapa and Eucalyptus urophylla in different seasons
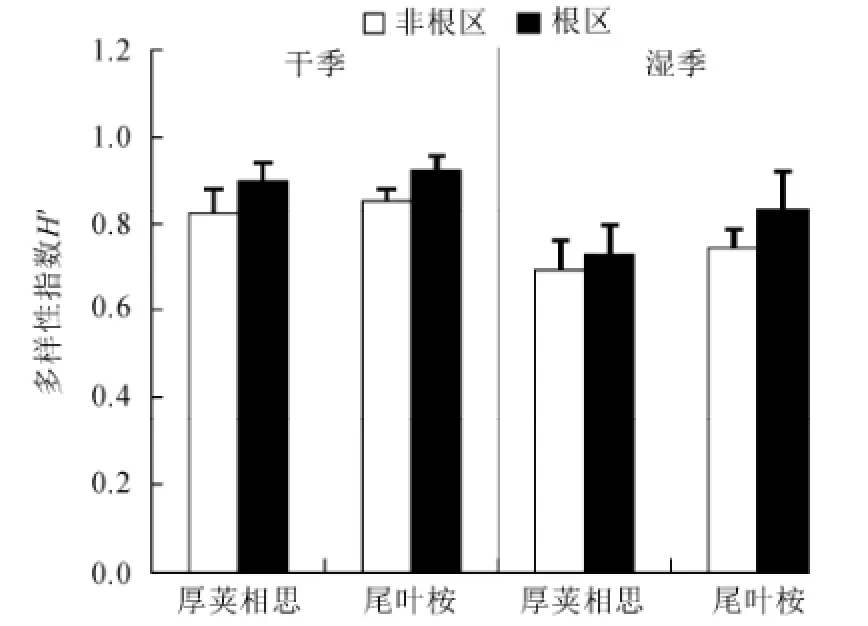
图4 不同季节厚荚相思和尾叶桉林型土壤线虫香农威尔多样性指数Fig.4 Shannon-Wiener diversity index of soil nematode in Acacia crassicapa and Eucalyptus urophylla in different seasons
干季土壤线虫的多样性指数为0.87,显著高于湿季0.75(图4)。不同林型、不同根区、不同坡度间多样性指数没有显著差异,但是根区土壤线虫的多样性指数有高于非根区的趋势。
2.3 土壤线虫和土壤性质的相关性
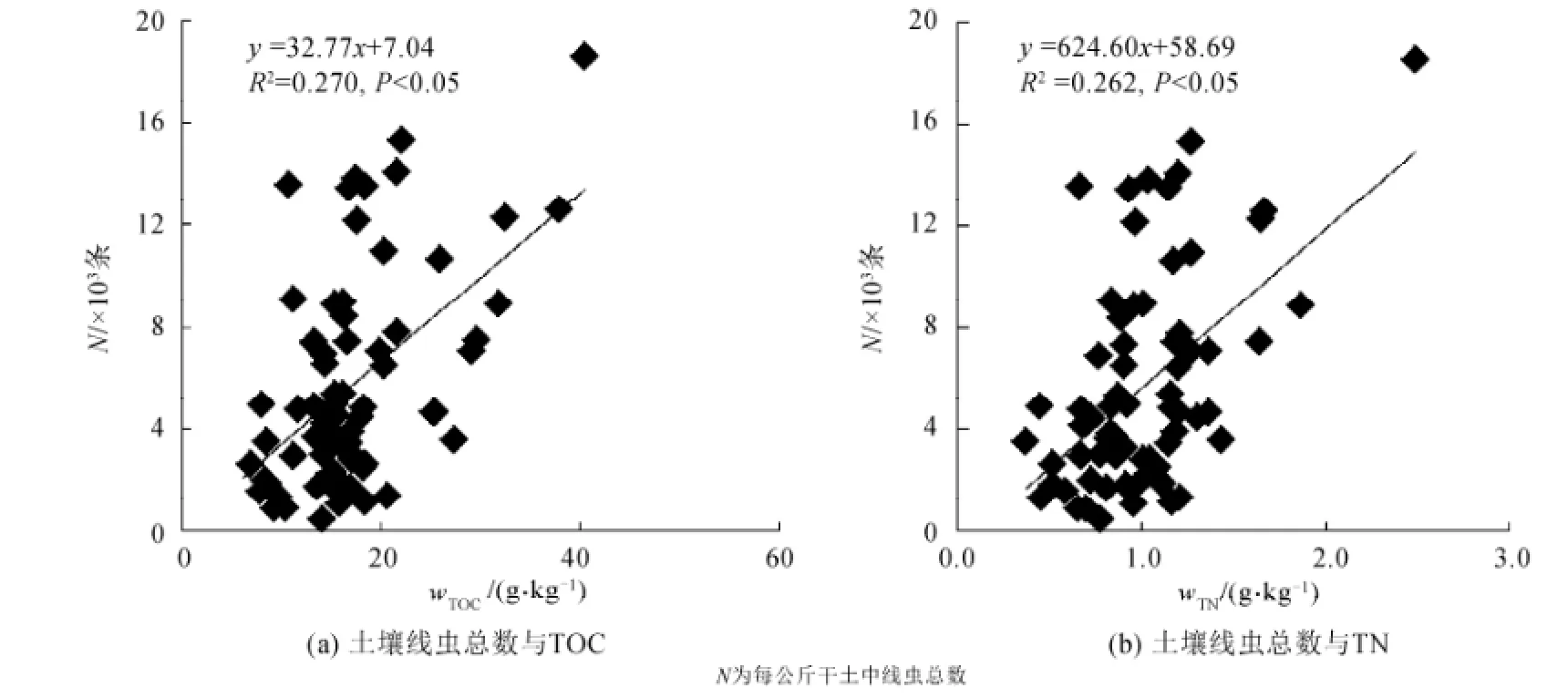
图5 土壤线虫总数与土壤总有机碳(TOC)和土壤总氮(TN)的关系Fig. 5 Correlations between soil nematode individuals with soil total organic carbon and total nitrogen
土壤线虫总数随总有机碳的升高而增加(图5),两者呈显著的正相关关系(R2=0.270, P<0.05)。土壤线虫总数随总氮的升高而增加,两者呈显著的正相关关系(R2=0.262, P<0.05)。线虫各营养类群的百分比、食真菌线虫与食细菌线虫的比值和线虫多样性指数与土壤性质没有相关性。
3 讨论
3.1 林型对土壤线虫的影响
平均土壤线虫总数为每公斤干土5750条,数量较大。在陆地生态系统中,森林生态系统土壤线虫最为丰富,尤其是开展本实验的亚热带森林生态系统,主要原因是森林具有丰富的植物多样性、丰富的凋落物和有机质积累,以及庞大复杂的地下根系分布,这为土壤线虫提供了一个资源丰富又相对稳定的生境(Brown和Gange,1990)。厚荚相思林型土壤线虫总数高于尾叶桉28.3%,差异显著(图1),且厚荚相思林型食细菌线虫的比例也显著高于尾叶桉(图2)。可能原因有以下几方面:第一,桉树存在强烈的化感作用,通过分泌特殊化学物质而对其他植物、林下植被和土壤动物、微生物产生抑制作用。化感物质的释放途径主要有地上部分的挥发、降水的淋溶、根系分泌和凋落物分解释放(Rice,1974;刘小香和谢龙莲,2004),其中后3种方式均可以将化感物质释放到土壤中影响土壤线虫和细菌数量。桉树对土壤线虫的抑制作用表现在直接和间接2个方面,化感物质可以直接影响线虫密度和细菌的生物量(Meyr和Huettel,1996;Hoagland等,2001),也可以通过减少地上植物群落的多样性和改变根区无机离子状态等,间接地影响土壤线虫和细菌的数量(Nilsson,1994)。第二,尾叶桉土壤含水量显著低于厚荚相思(表1),一方面是由于桉树生长迅速,需要大量的水来维持生长速率;另一方面是厚荚相思林下具有很厚的枯枝落叶层,不仅可以吸收降水和阻挡地表水径流,还可以减少土壤水分蒸发,使土壤疏松,通透保水性能更好(阳永泉等,2010)。这样,厚荚相思林型较高的土壤含水量很利于土壤线虫尤其是食细菌线虫的繁殖和生长。第三,厚荚相思根系十分发达,地下生物量远高于林分年龄相当的桉树(钟继洪等,2004),利于有机碳的积累,且厚荚相思为浅根树种,这为土壤表层微生物和线虫的发育和生殖提供了充足的食物资源;厚荚相思林下凋落物较多,也利于有机碳和总氮的积累。辽宁杨树林土壤线虫的研究结果表明,线虫总数量与土壤有机碳含量的相关性为R2=0.56,在水田和旱田则高达R2=0.86和R2=0.71(Ou等,2005),这与本研究结果吻合(图5)。因此在幼龄人工林,厚荚相思更易积累有机碳,利于食细菌线虫的增殖。
3.2 季节对土壤线虫的影响
干季厚荚相思和尾叶桉线虫总数差异不显著,而湿季厚荚相思林下线虫总数极显著地高于尾叶桉,表现出季节差异(图1)。华南地区地处亚热带季风气候带,雨热同期,适宜的温度和水分都利于土壤线虫和微生物的繁殖和发育(Verschoor等,2001)。湿季为植物的快速生长季,细根生物量也会达到最大,有研究表明线虫的密度与细根生物量呈显著的正相关关系(王邵军等,2008)。湿季厚荚相思凋落物的大量增加,导致土壤可溶性物质含量提高,促进了土壤微生物生物量的积累,这也是导致线虫总数增加的原因(Maxwell和Coleman,1995)。湿季食细菌线虫比例显著高于干季,而植物寄生线虫的比例显著低于干季,食真菌线虫和捕食杂食线虫无变化(图2),这是由于食细菌线虫数量的上升会与植物寄生线虫在空间上产生竞争,限制植物寄生线虫数量的增加(李琪等,2004),这是导致湿季食真菌线虫与食细菌线虫的比例低于干季的原因(图3)。在湿季土壤营养状况良好的情况下,本来占优势地位的食细菌线虫,由于较高的繁殖力(r-对策为主)比重近一步增加,造成了土壤线虫的多样性降低(图4),线虫群落对环境变化的敏感程度降低(李琪等,2004)。湿季土壤pH值显著低于干季(表1),可能原因是在植物的旺盛生长期,根系会分泌较多的有机酸造成土壤酸化,凋落物分解过程也会产生致酸物质(张希然和罗旋,1991)。湿季土壤pH值的降低引起土壤线虫总数的升高(图1),说明土壤线虫可耐微酸环境(Räty和Huhta,2003)。
3.3 根区对土壤线虫的影响
根区土壤线虫总数显著高于非根区(图1)。根区是受植物根系活动影响巨大,在物理、化学和生物学特性上与周围土壤有明显差别的土壤微区域,是植物根系和土壤生物群落互相作用的重要系统(王邵军等,2008)。由根系活动引起的根区土壤总有机碳和总氮的增加(表1),会提高土壤线虫的数量和多样性(图4);相反地,根区土壤线虫可以改变土壤养分的矿化速率和微生物的群落结构,间接改变植物之间的竞争平衡,从而对地上部分植物群落形成反馈作用(Kreuzer等,2004)。坡度对土壤线虫的总数、营养类群和多样性均无影响,这是因为试验地为丘陵地区,海拔差距较小,坡度缓慢,地表径流未对土壤线虫群落造成显著影响。
4 结论
尾叶桉土壤保水性差,会抑制土壤线虫数量和食细菌线虫比例;厚荚相思相对根系发达,保水能力强,为土壤自由生活线虫提供了优良环境,且在幼龄期没有表现出土壤酸化,生态效应优于尾叶桉。因此在中国华南地区人工林营林过程中可适当增加厚荚相思面积,特别是在退化较严重的地区,厚荚相思可作为恢复的先锋树种恢复土壤肥力。此外,厚荚相思和尾叶桉林型长期的生态环境效应还需要近一步研究探索。
致谢:感谢中国科学院华南植物园傅声雷研究员提供试验场地和实验指导。
BROWN V K, GANGE A C. 1990.Insect herbivory below ground[J]. Advances in Ecological Research, 20: 1-58.
HOAGLAND R E, ZABLOTOWICZ R M, OLESZEK W A.2001. Effects of alfalfa saponins on In Votro physiological activity of soil and rhizosphere bacteria[J]. Journal of Crop Production, 4: 349-361.
KREUZER K, BONKOWSKI M, LANGEL R, et al. 2004.Decomposer animals (Lumbricidae, Collembola) and organic matter distribution affect the performance of Lolium perenne (Poaceae) and Trifolium repens (Fabaceae)[J]. Soil Biology and Biochemistry, 36: 2005-2011.
MAXWELL R A, COLEMAN D C. 1995.Seasonal dynamics of nematodes and microbial biomass in soils of riparian-zone forests of the Southern Appalachians[J]. Soil Biology and Biochemistry, 27: 79-84.
MCSORLEY R. 1987.Extraction of nematodes and sampling methods[M]// In: Principles and practice of nematode control in crops.Sydney: Academic Press:13-41.
MEYR S L F, HUETTEL R N.1996. Application of a sex pheromone, pheromone analogs, and vertillium lecanii for management of Heterodera glycines[J]. Journal of Nematology, 28(1): 36-42.
NILSSON M C.1994. Separation of allelopathy and resource competition by the boreal dwarf shrub Empetrum hermaphroditum Hagerup[J]. Oecologia, 98: 1-7.
OU W, LIANG W, JIANG Y, et al.2005. Vertical distribution of soil nematodes under different land use types in an aquic brown soil[J]. Pedobiologia, 49: 139-148.
POORE M E D, FRIES C. 1985.The ecological effects of Eucalyptus[C]// Rome: Food and Agricultural 0rganization of the United Nation Forestry Papers, 59: 46-50.
RÄTY M, HUHTA V. 2003.Earthworms and pH affect communities of nematodes and enchytraeids in forest soil[J]. Biology and Fertility of Soils, 38: 52-58.
RICE E L. 1974.Allelopathy[M]. New York: Academic Press:166-179. SHIVA V, BANDYOPADHYAY J.1983. Eucalyptus- a disastrous tree for India[J]. The Ecologist,13: 184-187.
TANG C, YU Q. 1999.Impact of chemical composition of legume residues and initial soil pH on pH change of a soil after residue incorporation[J]. Plant and Soil, 215: 29-38.
VERSCHOOR B C, DE GOEDE R G M, DE HOOP J, et al.2001. Seasonal dynamics and vertical distribution of plant-feeding nematode communities in grasslands[J]. Pedobiologia, 45: 213-233.
YEATES G W, BONGERS T, DE GOEDE R G M, et al.1993. Feeding habits in soil nematode families and genera- an outline for soil ecologists[J]. Journal of Nematology, 25: 315-331.
李琪, 姜勇, 梁文举, 等. 2004.大气CO2浓度升高对稻田土壤线虫群落的影响[J]. 生态学杂志,23(3): 34-38.
李玉娟, 吴纪华, 陈慧丽, 等. 2005.线虫作为土壤健康指示生物的方法及应用[J]. 应用生态学报,16(8): 1541-1546.
廖崇惠. 1990.热带人工林生态系统的土壤动物[J]. 热带亚热带森林生态系统研究, 7: 141-147.
林婷. 2012.桉树人工林的生态效应研究[J]. 林业勘察设计, (1): 16-20.
刘菊秀, 周国逸, 温达志. 2000.广东鹤山陆地生态系统豆科树种林抗酸能力初探[J]. 资源生态环境网络研究动态, 11(3):1-5.
刘小香, 谢龙莲.2004. 桉树化感作用研究进展[J]. 热带农业科学,24(2): 54-61.
潘志刚, 林鸿盛. 1996.厚荚相思的引种、生长、良种繁育及利用[J].热带林业, 24(2): 52-58.
邵元虎, 夏汉平, 周丽霞, 等. 2007.香根草属与狼尾草属植物对土壤线虫营养类群的影响[J]. 热带亚热带植物学报,15(1): 1-8.
王邵军, 蔡秋锦, 阮宏华. 2008.不同生境柳杉根际线虫群落的生态特征[J]. 生态学杂志,27(4): 583-590.
肖石海, 方碧湘, 施纯淦, 等. 1996.厚荚相思等树种引种试验初报[J].福建林业科技, 23(增刊): 70-72.
阳永泉, 温远光, 何斌, 等. 2010.尾巨桉和厚荚相思人工林土壤肥力及其差异[J]. 林业科技开发, 24(2): 31-33.
张希然, 罗旋.1991. 红树林和酸性潮滩土[J]. 自然资源学报, 6(1): 55-62.
钟继洪, 李淑仪, 蓝佩玲, 等. 2004.刚果桉人工林营养元素生物循环研究[J]. 水土保持学报,18(6): 45-50.
Soil nematode community in Acacia crassicapa and Eucalyptus urophylla plantations
Wide areas of Acacia crassicapa and Eucalyptus urophylla were planted in South China recently due to their rapid growth, strong tolerance and suitability for paper pulp. However, it is still not clear how these two tree species impact soil biodiversity and fertility. It is necessary to evaluate their soil health condition scientifically. The number and variety of soil nematodes are abundant. Soil nematodes occupy the key positions in trophic levels of soil food web and are sensitive to environmental changes. As indicator of soil health, nematodes are used in ecology widely. In this study, soil nematodes were investigated in young Acacia crassicapa and Eucalyptus urophylla plantations with different seasons and sampling positions, to analyze the patterns of soil nematode individuals, community composition and diversity index, as well as relationships with environmental factors. The main results showed that: (1) Total nematode individuals in Acacia crassicapa plantation were 674.1 /100 g dry soil, more 28.3% than Eucalyptus urophylla plantation significantly. Percentages of bacterivores in Acacia crassicapa plantation were more 6.3% than Eucalyptus urophylla significantly. (2) Community composition of soil nematodes varied with the season. The percentage of bacterivores increased, whereas the percentage of plant parasites decreased in wet season. Diversity index decreased from 0.87 in dry season to 0.75 in wet season. Statistic differences were all significant. (3) Nematode density in rhizosphere was more 89.1% than non-rhizosphere which attributed to accumulated organic carbon and nitrogen in rhizosphere. In conclusion, Acacia crassicapa plantation provide optimal environment for soil free living nematodes, whereas soil acidification do not happen in young forest period. For ecological effects of Acacia crassicapa plantation are superior to Eucalyptus urophylla, planting areas of Acacia crassicapa may be expanded in South China in future.
biological indicator; trophic level; exotic species; rhizosphere; ecological effects
ZHAO Cancan*, WANG Wei
School of Life Science, Henan University, Kaifeng 475004, China
S154.1
A
1674-5906(2014)01-0073-07
赵灿灿,王伟. 厚荚相思和尾叶桉人工林土壤线虫群落研究[J]. 生态环境学报, 2014, 23(1): 73-79.
ZHAO Cancan, WANG Wei. Soil nematode community in Acacia crassicapa and Eucalyptus urophylla plantations [J]. Ecology and Environmental Sciences, 2014, 23(1): 73-79.
国家自然科学基金项目(31200408)
赵灿灿(1982年生),女,讲师,博士,从事土壤生态学研究,E-mail: cczhao2008@163.com;*通讯作者。王伟(1962年生),女,实验师,从事动物转基因育种和动物生理学实验研究。E-mail: wangwei8917@163.com
2013-08-25
