三种菊科入侵植物的生长与化学防御的关系研究
2014-02-07倪广艳朱丽薇牛俊峰赵秀华张振振赵培强
倪广艳,朱丽薇,牛俊峰,赵秀华,张振振,赵培强
中国科学院华南植物园,中国科学院退化生态系统植被恢复与管理重点实验室,广东 广州 510650
三种菊科入侵植物的生长与化学防御的关系研究
倪广艳*,朱丽薇,牛俊峰,赵秀华,张振振,赵培强
中国科学院华南植物园,中国科学院退化生态系统植被恢复与管理重点实验室,广东 广州 510650
“生活史理论”认为,植物可利用的资源总量是有限的,在植物的不同功能之间存在着此消彼长的权衡关系。入侵植物的生长和化学防御一般优于本地植物,那么其生长与化学防御之间是否存在权衡及其权衡关系怎样,目前尚不清楚。以广东省3种菊科入侵植物[三裂叶蟛蜞菊(Wedelia trilobata (L.) Hitchc.)、飞机草(Eupatorium odoratum)和薇甘菊(Mikania micrantha)]为研究对象,并分别以近缘或伴生的本地植物[蟛蜞菊(Wedelia chinenses)、华泽兰(Eupatorium chinense)和鸡矢藤(Paederia scandens)]为对照,研究入侵植物的生长特性(相对生长率和比叶面积)与化学防御物质(缩合单宁和总酚)含量,并基于这2种光合碳分配的主要形式,探讨入侵植物生长与化学防御之间的权衡关系。结果表明:3种入侵植物的相对生长率均高于本地对照种;薇甘菊的比叶面积大于对照种,而其他2种无明显优势。薇甘菊和三裂叶蟛蜞菊的缩合单宁显著高于对照种,飞机草的总酚含量高于对照种。我们的结果显示,入侵植物的生长和化学防御均优于本地植物,但它们的碳同化能力相近;因此,入侵植物特殊的内在资源分配与利用机制可能是其成功入侵的关键。
菊科入侵植物、相对生长率、比叶面积、总酚、缩合单宁
外来生物入侵(biological invasion)影响和改变陆地生态系统的生物多样性和功能过程,破坏生态系统的生产力与稳定性,已成为全球性的环境问题(Dukes和Mooney,1999)。从根本上控制外来种入侵,认识和掌握“什么样的物种更易成为入侵种”是入侵生态学、森林管理学研究和园林引种的关注热点。已有研究表明,外来植物成功入侵与其自身的生物学特性有关,表现为强竞争力、强繁殖力、强化感作用及宽生态幅等,而这些均涉及植物生长最初的过程-碳元素的获取与分配。Leishman等(2007,2010)比较研究了全球范围内近百种入侵植物与本地植物叶片的碳获取特征发现,两者在碳获取方面差异并不显著。然而,目前关于外来植物光合碳分配策略的研究则较少见。
“生活史理论”认为,植物体内可利用的资源总量是有限的,投入到某一功能(性状)资源量的增加必然会导致其投入到其它功能(性状)资源量的减少,即在植物的不同功能之间存在着“此消彼长”的权衡关系(trade-off)(Smith和Fretwell, 1974; Arendt, 1997;Paul-Victor等,2010)。然而,众所周知,在入侵地多数入侵植物具有明显的生长优势(Blossey和Nötzold, 1995; Vilà和Weiner, 2004; Bossdorf等, 2004; Ni等, 2010)。同时,入侵植物还通过多种途径如降低适口性、分泌化感物质、挥发油等,降低或避免其他生物(植物、微生物、昆虫等)的侵害,甚至排斥其他生物的生长(Callaway和Aschehoug, 2000; Prati和Bossdorf, 2004; Cappuccino和Arnason, 2006)。但是,这些研究结果多来自不同入侵物种,即便是同一入侵物种也是来自不同时间、不同地点。因此,简单地认为入侵植物具有高生长和强防御并不严谨,有必要在同一环境下研究同一无性系繁衍的入侵植物的生长特性与防御能力。
菊科植物是入侵植物的重要来源(朱世新等,2005;Ni等, 2012),它在繁殖上具有诸多优势,如有效的扩散机制、良好的集群能力,以及不需要专化传粉者等,因此它能够快速蔓延,成为入侵植物最多的类群之一(郝建华等,2009)。目前,对于菊科入侵植物的生长和防御国内外已有较多文献报道,但对于其生长与化学防御之间是否存在权衡及其权衡关系怎样,还未见相关报导。为此,本研究以华南地区3种菊科入侵植物为研究对象,并以其近缘或伴生植物为对照,研究同一生境下,入侵植物的生长特性和化学防御物质含量,并探讨生长与化学防御之间的权衡关系。
1 研究材料与实验方法
1.1 植物材料
本研究所用植物材料为三裂叶蟛蜞菊(Wedelia trilobata (L.) Hitchc.)与其近缘种本地蟛蜞菊(Wedelia chinenses)、飞机草(Eupatorium odoratum)与华泽兰(Eupatorium chinense)、薇甘菊(Mikania micrantha)与其生态型接近的鸡矢藤(Paederia scandens)。实验材料均采自中国科学院华南植物园游览区(23°10′22.65″N, 113°21′29.75″E)。取样时遵循的原则为,配对比较的两组植物伴生在同一样地。
1.2 实验方法
盆栽试验在华南植物园科研区温室内进行。根据不同植物的茎节特点,分别剪取5~10 cm长带芽茎段沙培扦插繁殖。经14 d预培养后,选取生长健壮且长势一致的再生小苗,移栽至小花盆(直径20 cm,高16 cm)内。每种植物种植10盆,每盆定植两株。培养基质采用河沙和塘泥混合物(体积比为4∶1)。同时,选取10株大小近似的扦插苗带回实验室洗净、烘干(65°,4 d)、称质量,获取初始生物量。盆栽苗每2 d浇水1次,每2周施肥1次(Miracle-Gro,1 g·L-1,100 mL),105 d后,收获植物并烘干(45°,5 d)、称质量,研磨后过80目筛,用于测定化学防御物质。
1.2.1 生长指标
一盆内的2株植物视为一个样本。叶面积使用叶面积仪测定(Li-3100A,Licor,美国)。相对生长率(relative growth rate,RGR)根据Imaji和Seiwa(2010)的方法计算,即RGR=(ln W2-ln W1)/t,W1为扦插苗移植时的平均生物量,W2为收获时每个样本的生物量,t为移植和收获之间的时间间隔(d)。
比叶面积(specific leaf area,SLA)为叶面积与生物量之间的比值。
1.2.2 化学防御指标
1.2.2.1 总酚提取与测定
取250 mg干燥样品于干净试管,加入80%甲醇10 mL,微波振荡5 min,1600 r·min-1离心10 min,取上清液置于干净试管,使用氮吹仪吹至1 mL。残渣重复提取1次。
采用Folin-Ciocalteu法测定总酚含量(Singleton和Rossi, 1965)。取100 μL样品置于干净试管,加入0.5 mL Folin-Ciocalteu试剂(Sigma),0.4 mL Na2CO3溶液(7.5%),静置30 min。取溶液于765 nm测吸光值(Perkin-Elmer λ15 UV-vis spectrophotometer,美国)。以没食子酸(gallic acid equivalent,GAE)为标准品作标准曲线,得到回归方程y=1.8917 x+0.0025(r2=0.9920)。样品中的总酚以没食子酸的质量分数表示(mg·g-1)。
1.2.2.2 缩合单宁提取与测定
缩合单宁的提取与测定,主要依据袁红娥等(2009)的方法,略作改动。准确称取50 mg样品,放入具塞试管中,加入5 mL 70%甲醇,封口摇匀,室温下放置24 h,5000 r·min-1离心10 min,取上清液备测。把4 g香草醛溶于100 mL纯甲醇中,配制成4%的香草醛甲醇溶液。取4%的香草醛甲醇溶液3 mL、浓盐酸1.5 mL和待测提取液0.5 mL加入以铝箔遮光的具塞试管中,摇匀,水浴20 min(20 ℃)。在510 nm下比色(Lambda 650,美国),重复3次,记录吸光值。以4%的香草醛甲醇溶液3 mL、浓盐酸1.5 mL和70%的甲醇0.5 mL作空白对照。以儿茶素为标准样品(Sigma公司),绘制标准曲线,根据其标准曲线计算出提取液缩合单宁含量。儿茶素标准曲线的绘制:取1.0 g·L-1儿茶素溶液配制质量浓度梯度标准溶液,分别在510 nm处测定吸光值,以儿茶素浓度为横坐标,吸光值为纵坐标绘制标准曲线。测定的标准曲线方程为:y=1.4719x+0.0941(r2=0.9941),y为吸光值,x为质量分数(mg·g-1)。
1.3 数据统计
数据统计使用SPSS 13.0软件(SPSS, 美国),差异显著性检测采用独立样本T检验进行,使用Pearson correlation分析入侵植物、本地植物的化学防御与植物生长之间的权衡关系。
2 结果与分析
2.1 入侵植物与本地植物的生长
经过105 d温室内栽培,入侵植物飞机草和薇甘菊的相对生长速率均显著高于本地对照植物(p<0.05,图1A)。三裂叶蟛蜞菊的相对生长率极显著高于本地蟛蜞菊(p<0.01)。说明在生长环境、基质等一致的条件下,入侵植物更容易快速生长,造成入侵。
比叶面积是指单位生物量上的叶片面积,体现了植物进行光合固碳的能力。我们的研究结果显示,入侵植物薇甘菊的比叶面积明显大于其伴生植物鸡矢藤(p<0.05),而另两组入侵植物与对照植物之间无显著差异(p>0.05)(图1B),说明这两组入侵植物与对照植物之间的碳同化能力相近。
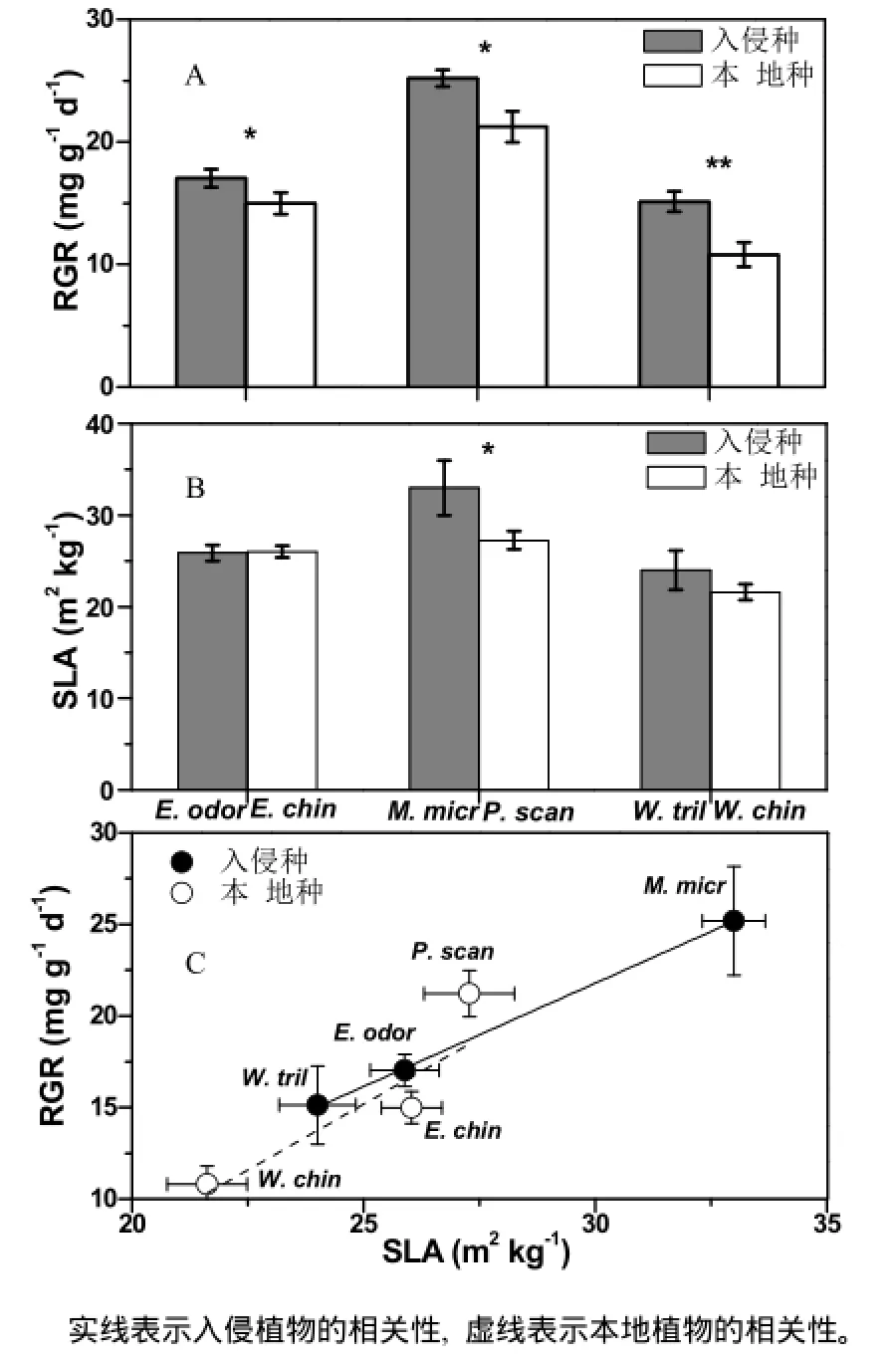
图1 入侵植物与本地植物的相对生长率、比叶面积及二者的关系比较Fig. 1 Comparisons of relative growth rate (A, RGR, mg·g-1·d-1) and specific leaf area (B, SLA, m2·kg-1) between invasive and native plant species; and C) the relationship of RGR and SLA of both the species. Bars represent means ± SEs, and asterisks indicate the significant differences
通过分析入侵种与本地种相对生长速率与比叶面积的关系(图1C)发现,无论是入侵种还是本地种其比叶面积和相对生长速率均呈显著正相关(p<0.05);而且还发现,本地植物RGR/SLA比值(斜率)较入侵植物高,揭示了本地植物光合产生的有机碳可能较入侵植物被更有效地用于生长。
2.2 三裂叶蟛蜞菊与本地蟛蜞菊的化学防御
总酚和缩合单宁是植物体内基于碳的、与抗虫性相关的重要次生代谢物质,它能够抑制昆虫对蛋白的消化(Bryant等, 1987; Matsuki等, 2004; Waterman和Mole, 1994; Nomura和Itioka, 2002)。我们的结果显示,入侵植物薇甘菊和三裂叶蟛蜞菊总酚的含量与本地植物差异并不明显(p>0.05),但是缩合单宁含量均显著高于其对照的本地植物(p<0.001)(图2A);入侵植物飞机草的总酚含量显著高于本地植物华泽兰(p<0.05),但二者缩合单宁含量差异不显著(p>0.05)(图2B)。
通过分析入侵植物与对照植物相对生长率的差值(生长能力)与其叶片缩合单宁含量的差值(化学防御能力)之间的关系,发现二者相对生长速率差值越大、缩合单宁含量的差值越大,呈显著正相关关系(图3),说明入侵植物的生长优势越强,其依赖缩合单宁的化学防御能力越强。
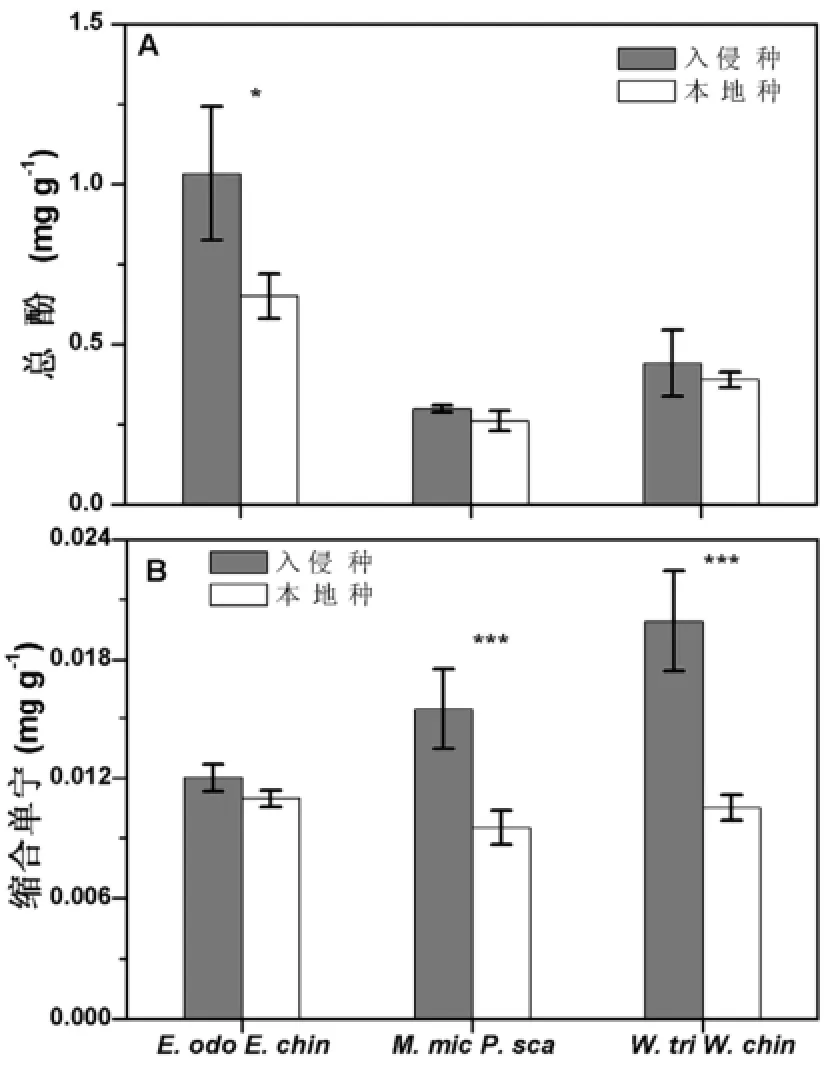
图2 入侵植物与本地植物缩合单宁含量的比较Fig. 2 Comparisons of total phenolics (A, mg·g-1) and condensed tannin (B, mg·g-1) between invasive and native plants. Bars represent means±SEs,and asterisks indicate the significant difference
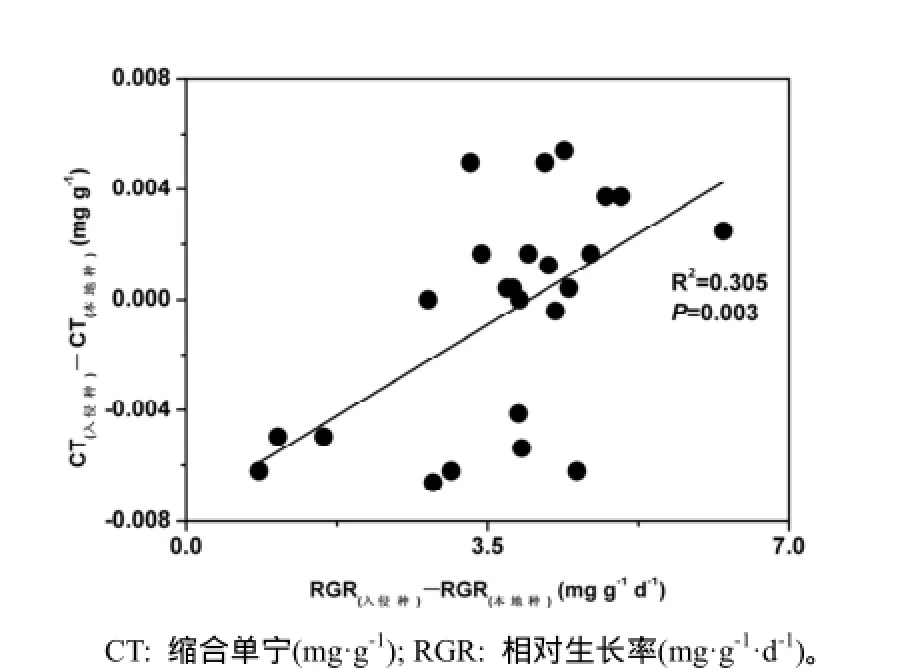
图3 入侵植物与本地植物之间相对生长率差与缩合单宁含量差的关系Fig. 3 Relationship of relative growth rate difference and condensed tannin difference between invasive and native plants
3 讨论
生长与防御是植物繁衍过程中光合碳进行分配的两个重要方面(Mooney,1972)。本研究结果显示,三种菊科入侵植物均具有明显的生长优势,同时,其化学防御能力也高于本地植物,而且生长优势越强、依赖缩合单宁的化学防御能力越强。我们的研究结果与前人得出的入侵植物生长强,其化学防御能力也强的结论一致(Ridenour等, 2008)。Almeida-Cortez等(1999)研究了31种草本菊科植物的生长与化学防御之间的权衡关系,结果也发现,菊科草本植物具有较高的相对生长率,其叶片的酚类物质含量也高。Imaji和Seiwa(2010)研究表明,蒙古栎(Quercus mongolica var. grosseserrate)幼苗的缩合单宁浓度与生长速率呈明显正相关。
相对生长率是指示植物生长的重要指标,已有研究表明,较高相对生长率有利于外来植物的成功入侵(Grime和Hunt, 1975; James和Drenovsky, 2007)。与前人的研究结果一致,本研究中涉及的3种菊科入侵植物均具有较高相对生长率(图1A)。比叶面积(SLA)反映了植物碳同化能力,是碳分配的重要前提(Reich等, 2003)。高SLA表示在相同的单位质量下,植物具有更大的叶片面积进行光合作用,进一步影响植物生长,因此也可用来判断植物的入侵性。然而,前人对入侵植物的SLA研究得出了不一致的结论。多数研究认为,入侵植物的SLA高于本地植物(Leishman等, 2007; Baruch和Goldstein, 1991; Baruch和Goldstein, 1999; Daehler, 2003; Osunkoya等, 2010);Feng等(2007)报道,在欧洲入侵的大叶醉鱼草(Buddleja davidii)与当地对照植物欧洲红瑞木(Cornus sanguinea)、西洋接骨木(Sambucus nigra)、垂枝桦(Betula pendula)等的SLA无明显差异。本研究中入侵植物三裂叶蟛蜞菊和飞机草和本地对照种的SLA近似,与Feng等(2007)的研究结果一致。薇甘菊的SLA明显大于对照种鸡矢藤,与Song等(2007)对相同植物研究得到的结果一致。我们的研究结果说明,三裂叶蟛蜞菊和飞机草的碳同化能力与本地对照种近似,薇甘菊的碳同化能力强于对照种。另外,前人的研究表明,SLA与RGR具有正相关关系(Lambers和Pooter, 1992; Reich等, 1997),本研究得出了一致的结果。
植物通过分泌次生代谢物进行化学防御,而这些物质绝大部分以碳元素为构建骨架(Rice,1984),因此,分泌化学物质是植物进行碳投资的重要方面(Orians等, 2010)。通常,入侵植物具有较强的化学防御能力,如散发挥发油(凌冰等,2003),释放化感物质等(Ni等, 2010; 倪广艳和彭少麟,2007;Chen等, 2009)。Cappuccino和Arnason(2006)研究认为,从入侵植物得到的一些化学物质更集中于某些种、属或科,而本地植物并无这种化学物质集中分布的现象。Kim和Lee(2011)发现,入侵植物的总酚含量约是本地植物的2.6倍。本研究结果显示,入侵植物薇甘菊和三裂叶蟛蜞菊均具有较高缩合单宁含量,飞机草具有较高总酚含量,显示这些菊科入侵植物具有较好的化学防御能力,可以抵御天敌昆虫啃食。
实际上,入侵植物具有较强的强化学防御能力不仅体现在其与入侵地的本地种比较上,也存在于其与原产地自身或其他本地种的比较上。如入侵我国华南地区的薇甘菊挥发油内普遍存在β-cadinene、allo-aromadendrene、β-caryophyllene及5-(1,1-dimethylethyl)-2,3-1H-Inden-1-one等多碳化合物,而生长在原产地秘鲁的薇甘菊检测不到这些物质,说明在入侵地入侵植物次生代谢物种类发生了改变,而且含量也明显增加(Ni等, 2007)。Pollock等(2009)的研究也表明,入侵植物斑点矢车菊(Centaurea diffusa)在入侵地分泌的化感物质(±)-catechin较原产地多。
尽管本研究未发现入侵菊科植物生长与化学防御之间存在明显的权衡关系,然而,这并不表示它们之间不存在权衡关系。影响入侵植物的防御与生长之间碳权衡分配的可能原因:1)入侵植物的建成成本低(Constructive Cost),不同植物构建相同生物量所需的碳量不同,建成成本较低的植物通常具有较高的生长速率(Lambers和Pooter, 1992)。Baruch和Gómez(1996)提出了入侵植物的低建成成本理论,认为构建相同生物量,入侵植物所需成本较低。如夏威夷群岛入侵植物的平均叶片建成成本较本地种低3%(Baruch和Goldstein, 1999)。入侵植物薇甘菊、三裂叶蟛蜞菊等的生物量建成成本低于本地植物肖梵天花(Urena lobata)和马缨丹(Lantana camara)等(Song等, 2007)。2)形成化学防御物质的成本,以本研究的缩合单宁和总酚为例,它们的形成亦需要成本,其高低与各自的分子量有关。一般而言,缩合单宁的分子量大,因此需要花费更多成本进行生产和保存(Briggs和Schultz, 1990);总酚是一类物质的总称,化学分子量各异,因此其生产和保存所花费的成本各异。3)有些酚类物质,如木质素(lignin),其作为生理结构的作用要远大于其作为化学防御物质的作用(Almeida-Cortez等, 1999),因此,出现防御物质与生长的正相关关系是可以预见的。4)化学防御物质的更有效性,在入侵地,由于本地生物(如植物、土壤微生物、昆虫等)与入侵植物缺乏长期协同进化的进程,因此本地生物对后者的化学防御物质反应敏感。如入侵地本地植物对入侵植物俄罗斯矢车菊(Acroptilon repens)的化学防御较原产地本地植物敏感(倪广艳和彭少麟,2007)。虽然本研究结果显示入侵植物具有生长和化学防御均优于本地植物,但是其内在生理学机制如何,是否是上述的某种或多种原因,还需要更多的研究深入探索。
参考文献:
ALMEIDA-CORTEZ J S, SHIPLEY B, ARNASON J T. 1999. Do plant species with high relative growth rates have poorer chemical defences?[J]. Functional Ecology, 13: 819-827.
ARENDT J D. 1997. Adaptive intrinsic growth rates: an integration across taxa[J]. Quarterly Review of Biology, 72: 149-177.
BARUCH Z, GOLDSTEIN G. 1999. Leaf construction cost, nutrient concentration and net CO2assimilation of native and invasive species in Hawaii[J]. Oecologia, 121: 183-192.
BARUCH Z, GóMEZ J A. 1996. Dynamics of energy and nutrient concentration and construction cost in a native and two alien C4grasses from two neotropical savannas[J]. Plant and Soil, 181: 175-184.
BLOSSEY B, NöTZOLD R. 1995. Evolutionary of increased competitive ability in invasive non-indigenous plants: a hypothesis[J]. Journal of Ecology, 83: 887-889.
BOSSDORF O, PRATI D, AUGE H, et al. 2004. Reduced competitive ability in an invasive plant[J]. Ecology Letters, 7: 346-353.
BRIGGS M A, SCHULTZ J C. 1990. Chemical defense production in Lotus corniculatus L. II. Trade-offs among growth, production and defense[J]. Oecologia, 83: 32-37.
BRYANT J P, CHAPIN III F S, REICHARDT P B, et al. 1987. Response of winter chemical defense in Alaska paper birch and green alder to manipulation of plant carbon/nutrient balance[J]. Oecologia, 72: 510-514.
CALLAWAY R M, ASCHEHOUG E T. 2000. Invasive plants versus their new and old neighbors: a mechanism fro exotic invasion[J]. Science, 290: 521-523.
CAPPUCCINO N, ARNASON J T. 2006. Novel chemistry of invasive exotic plants[J]. Biology Letters, 2: 189-193.
CHEN B M, PENG S L, NI G Y. 2009. Effects of the invasive plant Mikania micrantha H.B.K. on soil nitrogen availability through allelopathy in South China[J]. Biology Invasions, 11: 1291-1299.
DAEHLER C C. 2003. Performance comparisons of co-occurring native and alien invasive plants: implications for conservation and restoration[J]. Annual Review of Ecology, Evolution, and Systematics, 34: 183-211.
DUKES J S, MOONEY H A. 1999. Does global change increase the success of biological invaders?[J]. Trends in Ecology and Evolution, 14: 135-139.
FENG Y L, AUGE H, EBELING S K. 2007. Invasive Buddleja davidii allocates more nitrogen to its photosynthetic machinery than five native woody species[J]. Oecologia, 153: 501-510.
GRIME J P, HUNT R. 1975. Relative growth-rate: its range and adaptive significance in a local flora[J]. Journal of Ecology, 63: 393-422.
IMAJI A, SEIWA K. 2010. Carbon allocation to defense, storage, and growth in seedlings of two temperate broad-leaves tree species[J]. Oecologia, 162: 273-281.
JAMES J J, DRENOVSKY R E. 2007. A basis for relative growth rate differences between native and invasive forb seedlings[J]. Rangeland Ecology and Management, 60: 395-400.
KIM Y O, LEE E J. 2011. Comparison of phenolics compounds and the effects of invasive and native species in East Asia: support for the Novel weapons hypothesis[J]. Ecological Research, 26: 87-94.
LAMBERS H, POOTER H. 1992. Inherent variation in growth rate between higher plants: a search for physiological causes and ecological consequences[J]. Advances in Ecological Research, 23: 188-261.
LEISHMAN M R, HASLEHURST T, ARES A, et al. 2007. Leaf trait relationships of native and invasive plants: community-and global-scale comparisons[J]. New Phytologist, 176: 635-643.
LEISHMAN M R, THOMSON V P, COOKE J. 2010. Native and exotic invasive plants have fundamentally similar carbon capture strategies[J]. Journal of Ecology, 98: 28-42.
MATSUKI S, SANO Y, KOIKE T. 2004. Chemical and physical defense in the early and late leaves in three heterophyllous birch species native to northern Japan [J]. Ann. Bot.-Lon, 93:141-147.
MOONEY H A. 1972. The carbon balance of plants[J]. Annual Review of Ecology and Systematics, 3: 315-346.
NI G Y, LI F L, SONG L Y, et al. 2007. Allelopathic plants Mikania micrantha H.B.K[J]. Allelopathy Journal, 19: 287-295.
NI G Y, Schaffner U, Peng S L, et al. 2010. Acroptilon repens, an Asian invader, has stronger competitive effects on species from America than species from its native range[J]. Biological Invasions, 12: 3653-3663.
NI G Y, ZHAO P, HUANG Q Q, et al. 2012. Exploring the novel weapons hypothesis with invasive plant species in China[J]. Allelopathy Journal, 29: 199-214.
NOMURA M, ITIOKA T. 2002. Effects of synthesized tannin on the growth and survival of a generalist herbivorous insect, the common cutworm, Spodoptera litura Fabricius (Lepidoptera: Noctuidae) [J]. Applied Entomology and Zoology, 37: 285-289.
ORIANS C M, HOCHWENDER C G, FRITZ R S. 2010. Growth and chemical defense in willow seedlings: trade-offs are transient[J]. Oecologia, 163: 283-290.
OSUNKOYA O O, BAYLISS D, PANETTA F D, et al. 2010. Leaf trait co-ordination in relation to construction cost, carbon gain and resource-use efficiency in exotic invasive and native woody vine species[J]. Annuals of Botany, 106: 371-380.
PAUL-VICTOR C, Züst T, REES M, et al. 2010. A new methods for measuring relative growth rate can uncover the costs of defensive compounds in Arabidopsis thaliana[J]. New Phytologist, 187: 1102-1111.
POLLOCK J L, CALLAWAY R M, THELEN G C, et al. 2009. Catechin-metal interactions as a mechanism for conditional allelopathy by the invasive plant Centaurea maculosa[J]. Journal of Ecology, 97: 1234-1242.
PRATI D, BOSSDORF O. 2004. Allelopathic inhibition of germination by Alliaria petiolata (Brassicaceae) [J]. American Journal of Botany, 91: 285-288.
REICH P B, WALTERS M B, ELLSWORTH D S. From tropics to tundra: global convergence in plant functioning[J]. Proceedings of the national academy of sciences of the United States of America, 1997, 94: 13730-13734.
REICH P B, WRIGHT I J, CAVENDER-BARES J, et al. 2003. The evolution of plant functional variation: traits, spectra and strategies[J]. International Journal of Plant Science, 164: S143-164.
RICE E L. 1984. Allelopathy[M]. 2nd edn. Academic Press, Orlando.
RIDENOUR W M, VIVANCO J M, FENG Y, et al. 2008. No-evidence for trade-offs: Centaurea plants from America are better competitors and defends[J]. Ecological Monographs, 78: 369-386.
SINGLETON, V L, ROSSI JR J A. 1965. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents[J]. American Journal of Enology and Viticulture, 16: 144-158.
SMITH C C, FRETWELL S D. 1974. The optimal balance between size and number of offspring[J]. American Naturalist, 108: 499-506.
SONG L Y, NI G Y, CHEN B M, et al. 2007. Energetic cost of leaf construction in the invasive weed Mikania micrantha H. B. K. and its co-occurring species: implications for invasiveness[J]. Botanical Studies, 48: 331-338.
VILà M, WEINER J. 2004. Are invasive plant species better competitorsthan native plant species? Evidence from pair-wise experiments[J]. Oikos, 105: 229-238.
WATERMAN P G, MOLE S. 1994. Why are phenolic compounds so important[C]//Waterman P G, MOLE S. Analysis of phenolic plant metabolites. Blackwell Scientific Publications, Boston, USA: 44-65.
郝建华, 刘倩倩, 强胜. 2009. 菊科入侵植物三叶鬼针草的繁殖特征及其与入侵性的关系[J]. 植物学报, 44: 656-665.
凌冰, 张茂新, 孔垂华, 等. 2003. 飞机草挥发油的化学组成及其对植物、真菌和昆虫生长的影响[J]. 应用生态学报, 14: 744-746.
倪广艳, 彭少麟. 2007. 外来入侵植物化感作用与土壤相互关系研究进展[J]. 生态环境, 16: 644-648.
袁红娥, 袁善春, 佟丽丽, 等. 2009. 剪叶损伤与昆虫取食对兴安落叶松(Larix gmelinii) 针叶中缩合单宁诱导作用的差异[J]. 生态学报, 29: 1415-1420.
朱世新, 覃海宁, 陈艺林. 2005. 中国菊科植物外来种概述[J]. 广西植物, 25: 69-76.
Growth and chemical defense in three Asteraceae invasive weeds in lower subtropical China
NI Guangyan*, ZHU Liwei, NIU Junfeng, ZHAO Xiuhua, ZHANG Zhenzhen, ZHAO Peiqiang
South China Botanical Garden, Chinese Academy of Sciences, Key Laboratory of Vegetation Restoration and Management of Degraded Ecosystems, Chinese Academy of Sciences, Guangzhou 510650, China
A universal theory in ecology is that the allocations of limited resources to growth and defense are trade-off, while this theory appeared not to hold for invasive plants according to the previous studies. In this study, three invasive Asteraceae plant species in the Guangdong province i.e., Eupatorium odoratum, Mikania micrantha, Wedelia trilobata, and their congeneric or co-occurring native plant species i.e., Eupatorium chinensis, Paederia scandens, Wedelia chinensis were used to detect if there is a trade-off between growth and chemical defenses on the basis of carbon allocation, and investigate the trade-off relations between them. All the three invasive Asteraceae had higher relative growth rate (RGR) as compared to their related native counterparts, and M. micrantha had higher specific leaf area (SLA) as well. Two invasive Asteraceae (i.e., M. micrantha and W. trilobata) had higher condensed tannin content, while the other invasive species E. odoratum had higher total phenolics content, as compared to their native species. Our results indicate that the invasive plants have similar carbon assimilation but different carbon allocations between the growth and chemical defense, thus implying that the special underlying carbon utilization mechanism may be a key for their successful invasions.
Asteraceae invasive plants; Relative growth rate; Specific leaf area; total phenolics; condensed tannin
Q948
A
1674-5906(2014)01-0001-06
倪广艳,朱丽薇,牛俊峰,赵秀华,张振振,赵培强. 三种菊科入侵植物的生长与化学防御的关系研究[J]. 生态环境学报, 2014, 23(1): 1-6.
NI Guangyan, ZHU Liwei, NIU Junfeng, ZHAO Xiuhua, ZHANG Zhenzhen, ZHAO Peiqiang. Growth and chemical defense in three Asteraceae invasive weeds in lower subtropical China [J]. Ecology and Environmental Sciences, 2014, 23(1): 1-6.
国家自然科学基金项目(31200380);广东省自然科学基金(S2011040000331;S2013010012054);中国科学院知识创新工程项目(KSCX2-EW-J-28)
倪广艳(1978年生),女,助理研究员,博士,从事植物生理生态学研究。E-mail: guangyan.ni@scbg.ac.cn *通信作者
2013-10-11
