谷氨酰胺促泥鳅生长机理的初步研究
2014-01-21温安祥骆美琳
■李 源 温安祥 骆美琳
(1.四川农业大学,四川雅安625014;2.新乡市林业局,河南新乡 453000)
泥鳅(Misgurnus anguillicaudatus)隶属于鲤形目,鳅科,泥鳅属[1],全国各地都有分布。泥鳅作为一种小型经济鱼类,由于生活环境的特殊性,使之成为重要的科学研究对象。
谷氨酰胺(Glutamine,Gln)是构成蛋白质的基本氨基酸之一,对机体免疫及营养作用的研究主要集中在人畜禽方面,关于水生动物方面的研究只有零星报道,尤其是在泥鳅方面还未见报道。泥鳅是我国淡水特种经济动物之一,具有抗病力强、营养价值高、肉质鲜美,并含有多种维生素是人们喜爱的水产佳品,远销日本、韩国等地,是我国重要的出口水产品。泥鳅食性杂,各地均有分布,对水质环境的好坏具有很好的标志作用。目前由于泥鳅养殖的规模化、集约化,养殖环境逐渐恶化,疾病发病率逐年增高。因此,泥鳅配合饲料中免疫增强剂的研发显得极其重要。本试验探讨了饲料中添加不同水平的谷氨酰胺对泥鳅生长、非特异性免疫的影响,以期为进一步开发泥鳅配合饲料提供参考依据。
1 材料与方法
1.1 试验动物与日粮组成
试验鱼为雅安鱼种站2012年孵化的同一批健壮泥鳅(Misgurnus anguillicaudatus)鱼种,用3%的NaCl消毒10 min,饲养于四川农业大学生命科学与理学院动物学实验室。用基础饲料驯养10 d,备用。基础日粮组成及营养成分见表1[2]。
1.2 试验设计
选取360尾平均体重为(3.31±0.11)g的泥鳅随机分为6组(A~F组),每组设3个重复,每个重复20尾。饲养于规格为60 cm×45 cm×45 cm的鱼缸中。在基础日粮中添加浓度为0(A组)、0.2%(B组)、0.4%(C组)、0.8%(D组)、1.6%(E组)和3.2%(F组)的谷氨酰胺进行饲喂。其中对照组为A组,添加组为B~F组。
1.3 饲养管理
每天按鱼体重的3%投喂饵料,早8点、晚6点定时定量投喂;试验期间水温为23~27℃,pH值为7.2~7.4。每次投喂前观察鱼的活动,投喂1 h后观察残饵情况,试验期间每天采用虹吸法换水一次并根据残饵情况调节饲喂量,并记录水温、投喂量等;每周用2~3 g∕m3的五倍子[3]进行疾病预防。饲养试验80 d。
1.4 试验样品的采集
饲养试验结束后,在最后一次采食12 h后,从每个重复中随机捞取10尾作为测定样本。称取体重和测定体长后,在体表用酒精消毒后于无菌操作台中解剖并迅速取出内脏,分离脾脏、肝胰脏、肠道,称重并测量肠长;肠道一部分保存于10%的甲醛溶液中,用于组织切片观测;另一部分进行组织匀浆的制备:滤纸吸干肝胰脏与肠道表面多余水分,并剪开泥鳅肠道去除内容物,与预冷的0.85%NaCl按1∶9体积比于4℃条件下匀浆成10%组织匀浆液,3 000 r∕min冷冻离心10 min[4],上清液即为酶液,用于消化酶的测定。体表黏液的制备采用自然分泌法:每个重复随机取2尾鱼,用蒸馏水冲洗泥鳅体表后,并用滤纸轻轻吸干残余水分,用1 ml的注射器吸取1 ml的生理盐水,缓慢均匀地滴在放置于培养皿内蠕动的泥鳅体表,泥鳅受生理盐水刺激,蠕动激烈并分泌黏液,计时1 min后收集培养皿内黏液与生理盐水的混合液。
1.5 测定指标
1.5.1 生产性能和器官指标测定

式中:Wt——终末体重(g);
W0——初始体重(g);
t——试验持续天数(d)。
1.5.2 肠道酶活性
取肠道组织匀浆液分别测定蛋白质含量,蛋白酶、脂肪酶和淀粉酶活力。蛋白酶活力测定采用福林-酚法,淀粉酶活力测定采用碘-淀粉比色法,脂肪酶活力测定采用聚乙烯醇橄榄油法。
1.5.3 肠道组织切片
将泥鳅肠道分为前、中、后肠,采用常规组织学切片方法分别对其切片观测,计算绒毛高度及密度。绒毛密度为单位肠壁横切面内,绒毛中间部位宽度所占的横截面的比例。
1.5.4 非特异性免疫指标的测定
1.5.4.1 SOD活力
通过黄嘌呤及黄嘌呤氧化酶反应,系统产生超氧阴离子自由基(),后者氧化羟胺形成亚硝酸盐,在显色剂的作用下呈现为紫红色,用可见光分光光度计测其吸光度。当被测样品中含SOD时,则对超氧阴离子自由基有专一性的抑制作用,从而使形成的亚硝酸盐减少,比色时测定管的吸光度值低于对照管的吸光度值,通过公式计算可求出被测样品中的SOD活力。具体参照南京建成SOD测试盒说明书。
1.5.4.2 溶菌酶活力
溶菌酶活力采用南京建成生物研究所溶菌酶测试盒进行测定:在一定浓度的浑浊菌液中,溶菌酶能水解细菌胞壁上的肽聚糖,使细菌裂解造成菌液浓度降低,透光性增强,根据透光度的变化来推测溶菌酶的活力。具体方法参照测试盒说明进行。
1.6 数据处理
试验结果用“平均数±标准差”表示,用SPSS20先对数据作方差齐性检验(ANOVA),若处理间差异显著,再进行Duncan's法多重比较,P<0.05表示差异显著,P<0.01表示差异极显著,P>0.05表示无显著性差异。
2 试验结果
2.1 谷氨酰胺对泥鳅生长性能的影响(见表2)
由表2可知,饲料中添加不同水平的谷氨酰胺,对于泥鳅的增重率、特定生长率和饵料系数均有影响;添加组(B~F组)增重率较对照组均有所升高,C~E组显著高于对照组(P<0.05),其中,E组最高,而C~E组之间差异不显著(P>0.05);特定生长率C~E组较对照组显著升高(P<0.05),其中C~E组之间差异不显著(P>0.05),其他各组较对照组虽有所升高,但差异不显著(P>0.05);添加组(B~F组)饵料系数较对照组均有所降低,C~E组显著低于对照组(P<0.05)。

表2 谷氨酰胺对泥鳅生长性能的影响
谷氨酰胺对泥鳅器官发育指数的影响见表3。谷氨酰胺对泥鳅肠体指数有影响,随着添加量的增加,E、F组显著低于D组(P<0.05),其余各组之间差异不显著(P>0.05),但B、C组较对照组有所增高,而E、F组较对照组略低;谷氨酰胺对肝胰脏有影响,C组显著高于D组(P<0.05),D组极显著高于对照组(P<0.01);谷氨酰胺对肠长指数及脾脏指数影响不显著(P>0.05)。

表3 谷氨酰胺对泥鳅器官发育的影响(%)
2.2 谷氨酰胺对泥鳅消化道结构的影响(见表4、表5及图1)
由表4可知,谷氨酰胺对泥鳅前肠绒毛高度和绒毛密度均有影响。其中C~F组的前肠绒毛高度极显著高于对照组(P<0.01),B组显著高于对照组(P<0.05);前肠绒毛密度D~F组极显著高于对照组(P<0.01),C组显著高于对照组(P<0.05)。

表4 谷氨酰胺对泥鳅前肠绒毛高度及密度的影响

表5 谷氨酰胺对泥鳅中肠绒毛高度及密度的影响
据表5,谷氨酰胺对泥鳅中肠绒毛高度也有影响,其中D~F组的中肠绒毛高度极显著高于对照组(P<0.01),C组显著高于对照组(P<0.05),B组与对照组之间差异不显著(P>0.05);对中肠绒毛密度影响不显著(P>0.05)。
泥鳅后肠如图1所示,由图可见,泥鳅后肠肠壁较前肠与中肠薄,后肠绒毛极短且稀疏,紧贴肠壁,绒毛及肠壁周围分布有大量红细胞。
2.3 谷氨酰胺对泥鳅消化酶的影响(见表6)
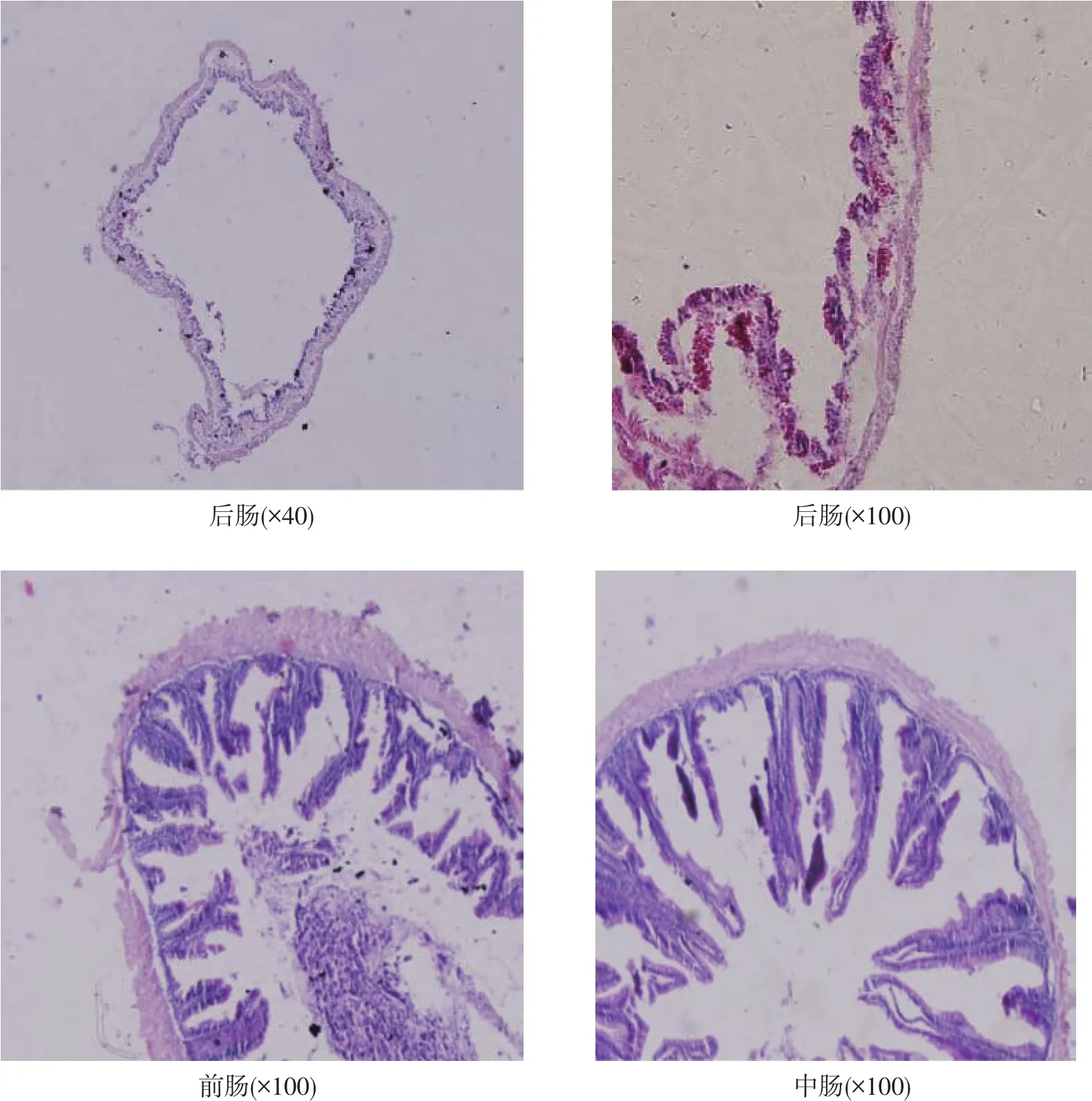
图1 泥鳅肠道切片图

表6 谷氨酰胺对泥鳅肠道消化酶活力的影响
由表6可知,谷氨酰胺对泥鳅肠道消化酶有极显著(P<0.01)或显著影响(P<0.05)。B~E组脂肪酶活力较对照组都有所升高,其中B~D组极显著高于对照组(P<0.01),E组显著高于对照组(P<0.05),F组活力较对照组有所下降;添加组(B~F组)的淀粉酶活力较对照组均极显著升高(P<0.01),整体趋势呈先升高后降低,其中D组数值最高,极显著高于对照组(P<0.01);添加组(B~F组)蛋白酶活力较对照组均有所升高,其中E组最高,极显著高于对照组(P<0.01),D组、F组显著高于对照组(P<0.05);在肠道中,γ-谷氨酰转移酶随添加量的增加而升高,F组极显著高于对照组(P<0.01),并显著高于其他添加组(P<0.05),B~E组较对照组显著升高(P<0.05)。
2.4 谷氨酰胺对泥鳅体表SOD、体表溶菌酶活力的影响(见表7)

表7 谷氨酰胺对泥鳅体表黏液SOD和溶菌酶活力的影响
从总体上看,添加组体表黏液SOD随添加量的增加而增加,C~F组显著高于对照组,其中E~F组极显著高于对照组(P<0.01);体表黏液溶菌酶活力C~E组极显著高于对照组(P<0.01),B、F组较对照组之间差异不显著(P>0.05),整体趋势呈先升高后降低。
3 讨论
3.1 不同谷氨酰胺水平与泥鳅的生长性能
本试验结果表明,当饲料中谷氨酰胺添加量从0.4%上升到1.6%时,泥鳅的增重率和特定生长率呈上升趋势,且显著高于对照组(P<0.05),饵料系数C~E组显著低于对照组(P<0.05)。说明泥鳅饲料中添加谷氨酰胺有明显的促生长的作用。目前饲料中添加谷氨酰胺的研究主要集中在陆生动物及少数水生动物上,对泥鳅的影响还未见报道。戴四发等[5]在1~4周龄肉鸡日粮中添加0.4%的谷氨酰胺,能够提高肉鸡的生产性能;蒋宗勇等[6]在4日龄新生仔猪中添加1.0%的谷氨酰胺,18 d后显著提高仔猪日增重率;杨奇慧等[7]在1~4周龄杂交罗非鱼中的试验表明,谷氨酰胺添加量从0.2%上升到0.8%时,杂交罗非鱼的增重率和SGR呈上升趋势;林燕[8]在鲤鱼饲料中添加谷氨酰胺,含量从0.0%提高到1.2%时,极显著提高了幼健鲤的增重率。这些都说明在日粮中添加谷氨酰胺对动物生长性能有较好的提高,谷氨酰胺促进动物生长的可能原因有以下几方面:谷氨酰胺为细胞内核苷酸的合成提供氮源[9],促进体内DNA的合成;在组织细胞培养中,谷氨酰胺是必需添加的氨基酸[10],谷氨酰胺的添加量与细胞增殖量在一定程度内正相关;谷氨酰胺通过转氨基作用调节体内氨基酸的代谢,促进肌肉蛋白质的合成和转运;Xin等[11]报道,谷氨酰胺可以明显增加大鼠血浆胰岛素生长因子Ⅰ水平,刺激动物生长,而胰岛素生长因子Ⅰ是一类多功能细胞增殖调控因子,在细胞分化、增殖及个体发育中具有关键的促进作用。
谷氨酰胺可影响机体器官发育,在鱼肠上皮组织体外培养中发现,谷氨酰胺可显著促进鱼肠上皮细胞的增殖和分化[12];黄冠庆[13]在肉仔鸡日粮中添加谷氨酰胺发现,0.8%的浓度可显著提高21日龄时的脾脏、肝脏和胰腺的重量;林燕[8]在鲤鱼日粮中添加谷氨酰胺,可显著提高鲤鱼肝胰脏、肠道和脾脏的重量。本试验中通过对泥鳅日粮中添加外源性谷氨酰胺,当添加量为0.8%时,可显著(P<0.05)或极显著(P<0.01)提高泥鳅肠体指数和肝胰脏指数,但对泥鳅肠长指数影响不显著,肠道长度的增强可以延长食物在肠道内停留的时间,但由于泥鳅肠道的特殊性,作为泥鳅的呼吸辅助器官,泥鳅肠道与其他动物(如建鲤)肠道结构差异较大,泥鳅肠道将食道和肛门连在一起形成一直管,肠长主要根据体长变化,因此谷氨酰胺对泥鳅肠长指数影响不大。肝胰脏和肠道为泥鳅体内重要的免疫消化器官,因此其发育程度直接影响了泥鳅整体的生长状况。
3.2 谷氨酰胺促进泥鳅对营养物质的消化、吸收
3.2.1 不同谷氨酰胺水平对泥鳅肠道结构的影响
动物肠道是利用谷氨酰胺最多的器官,且谷氨酰胺是肠道主要能源物质[14]。谷氨酰胺能显著增加肠道蛋白质的合成代谢[15],促进动物肠上皮分化增殖[16]。研究表明,外源性的谷氨酰胺可显著促进肉仔鸡小肠发育,维持肠黏膜的完整性,以及提高小肠的主动吸收功能[17];可促进鲤鱼肠道生长,提高绒毛高度[18];可使鼠肠绒毛密度显著高于对照组,细胞及微绒毛紧密排列的程度大于对照组[19]。绒毛高度是反映水生动物肠道生长发育和吸收能力的重要标识[20],肠道绒毛是由肠上皮和固有层向肠腔突起形成,其高度的增加大大增加了肠道内部的吸收面积,而绒毛密度是对高度的一个补充。本试验中泥鳅前肠与中肠绒毛高度及前肠绒毛密度随着添加量的增加而升高,但对中肠绒毛密度影响不显著(P>0.05)。结合泥鳅肠呼吸的特点及肠道组织切片分析,可能是因为:泥鳅肠道后肠绒毛不明显,且肠壁极薄,泥鳅肠道消化吸收部位主要位于前中肠[21];而后肠的结构特点对肠呼吸的气体交换有很大的作用,是泥鳅肠呼吸的主要部位;中肠与后肠无明显分界,因此中肠至后肠绒毛密度逐步趋于稳定,一定程度上保证了气体交换面积;泥鳅肠道整体绒毛高度的降低也利于气体及粪便的通过,便于肠呼吸的正常进行,因此分析泥鳅肠道绒毛的生长发育可能在肠道消化吸收作用与肠呼吸作用之间存在一个平衡点。
3.2.2 不同谷氨酰胺水平对泥鳅消化酶活力的影响
鱼体利用营养物质的能力依赖于消化酶,鱼体内消化酶主要分为蛋白酶、脂肪酶、淀粉酶,不同种类鱼体内三种酶的活力是不同的,主要取决于鱼类的食性以及生活习惯[22]。鱼体内部或外部条件改变使其消化酶活力发生变化,会直接影响鱼对饲料中营养物质的利用率,消化酶的变化可能跟饲料中添加物有关,当食物组分发生变化时,在一定程度内也会影响消化酶活力[23]。由于泥鳅是无胃鱼,因此肠道是其主要的消化场所,消化酶活力的高低可直接反映其对营养物质的吸收利用。
研究表明,谷氨酰胺可显著提高畜禽类及鲤鱼[4,8,24-26]消化道消化酶活力。本试验结果表明,日粮中添加谷氨酰胺,可提高泥鳅肠道内消化酶活力。蛋白酶活力的高低会影响水生动物体内蛋白质的消化程度,进而影响水生动物体内蛋白质沉积效率。γ-谷氨酰转移酶是参与γ-谷氨酰循环和氨基酸跨膜转运的关键酶,具有转肽等重要功能[27]。张军民等[28]通过对不同日龄仔猪的体内γ-谷氨酰转移酶含量的测定发现,在动物体内,当自身氨基酸合成不足时,需要通过外源摄取来补充自身蛋白质含量,因此γ-谷氨酰转移酶含量会发生变化,即逐步升高,当仔猪体内氨基酸合成逐渐满足日常需要时,γ-谷氨酰转移酶含量逐渐降低,因此γ-谷氨酰转移酶对外源氨基酸的吸收起关键作用。本试验中通过添加谷氨酰胺显著提高了泥鳅肠道γ-谷氨酰转移酶含量,因此对泥鳅肠道氨基酸和蛋白质的吸收、吸收和跨膜转运有显著的促进作用。
3.3 谷氨酰胺对泥鳅非特异性免疫的影响
溶菌酶(lysozyme,LYZ)是一类酶的总称,又称为胞壁质酶、N-乙酰胞壁质聚糖水解酶、G蛋白,能水解细菌胞壁N-乙酰葡糖胺和N-乙酰胞壁酸之间的β-1,4糖苷酸位点的肽聚糖,引起细胞裂解。溶菌酶是体内重要的非特异性免疫因子,具有与抗生素不同的抑菌机理,在杀死病原体时,不宜使其产生耐药性,并且具有无毒、无残留、活性稳定、抗菌谱广等优点[29]。在水生动物体内溶菌酶水平是非特异性免疫的重要指标,广泛存在于黏液、淋巴组织、血液等其他体液中。溶菌酶的活力与性别、年龄、季节、温度、pH值、外源病毒感染程度等因素有关[30]。本试验研究结果发现,泥鳅日粮中添加谷氨酰胺提高了泥鳅血浆和体表黏液溶菌酶活力,血浆溶菌酶B~D组活力呈上升趋势,D组极显著高于对照组(P<0.01),从E组开始,溶菌酶活力逐渐降低,与对照组相比差异不显著(P>0.05);黏液溶菌酶C~E组极显著高于对照组(P<0.01),但F组与对照组相比差异不显著(P>0.05);陈昌福等[31]、刘勇等[32]发现,草鱼、鲫鱼等黏液溶菌酶活力较血浆中高,本试验中也证明泥鳅黏液溶解酶活力比血浆溶菌酶活力高,这与前面学者研究结果一致,水生动物黏液可以说是第一道免疫防线。
鱼类体表黏液中含有大量能够抵抗外源病原体入侵的非特异性免疫因子,除溶菌酶外还含有可抑制病毒复制的干扰素、可作为调节细胞吞噬的C-反应蛋白等[33]。黏液除具有免疫功能外,还可以防止异物和病原体的侵入;维持体内渗透压;减少运动摩擦力等功能[33]。梁明山等[34]在泥鳅黏液中发现SOD,本试验进一步证明泥鳅黏液中含有SOD,而且泥鳅黏液中SOD的含量与梁明山等[34]所测结果6.5 U∕ml具有一致性。本试验表明,在泥鳅日粮中添加谷氨酰胺可以提高黏液SOD酶活力,并随着添加量的增加,SOD活力逐步升高。
结合泥鳅生长性能、消化吸收能力以及非特异性免疫试验结果及变化规律,本试验表明,添加高剂量(1.6%~3.2%)谷氨酰胺对泥鳅生长出现一定程度抑制作用。水生动物对氨基酸需要量根据动物的种类、年龄、食性、发育阶段、水温、水质、pH值以及饲料中蛋白含量的种类和饲喂方式不同而变化[35];另外氨基酸在水生动物肠道内的转运机制较为复杂,一种氨基酸的转运可能会兼有许多转运机制,而鱼体对游离氨基酸的吸收率相当高,但是日粮中结合态的蛋白质较游离氨基酸更能刺激鱼体的生长[36],因此推测谷氨酰胺添加量在一定范围内能显著增加肠道蛋白质[15]的合成代谢,促进肠道发育,但随着浓度的增高,添加的游离氨基酸先于饲料蛋白中分解氨基酸的吸收,同时还影响其他氨基酸吸收同步化,使氨基酸得不到平衡互补,进而影响泥鳅生长发育。
