尼古丁污染及其微生物降解研究进展
2014-01-14何厚龙柳华贵钟卫鸿
焦 洋,何厚龙,柳华贵,钟卫鸿
(浙江工业大学生物与环境工程学院,浙江 杭州310032)
尼古丁(nicotine),又名烟碱,广泛存在于茄科(Solanaceae)植物中。作为一种生物碱,尼古丁含量占烟草植物生物碱总量的95%,是烟草的重要成分,也是一种环境有毒物质。尼古丁易溶于醇、氯仿及石油醚等有机溶剂,是具有碱性的二元碱。根据N-甲基四氢吡咯在吡啶环上位置的不同,尼古丁可分为3种立体异构体:α-烟碱、β-烟碱和γ-烟碱,其中β-烟碱是尼古丁的主要存在形式。
一些微生物可以通过不同的代谢途径(如吡啶途径、吡咯途径、脱甲基途径等)利用尼古丁,具有研究和应用价值。近年来,利用微生物降解尼古丁已成为国内外研究的热点。作者在此对尼古丁的危害及污染来源、微生物降解途径等进行综述。
1 尼古丁的危害及污染来源
1.1 尼古丁的危害
尼古丁不仅对人类有致毒、致癌等副作用,而且影响土壤生态结构,污染地下水,对土壤和环境造成严重污染,扰乱生态平衡。1994年,尼古丁被美国环保局正式列入有毒物质释放目录(Toxics Release Inventory)[1]。大量尼古丁可以抑制中枢神经系统,造成呼吸停止,心脏麻痹。尼古丁一旦进入人体细胞,就会变为一种神经毒素,使人产生兴奋感,长期作用会导致依赖性,是吸烟成瘾的主要作用物质;还可以转化成多种具有高生物活性的物质(如NNN、NNK、cotinine、TSNA等),显著提高癌症(如肺癌等)的并发率[2]。
尼古丁在细胞水平上的影响还有:抑制细胞增殖和凋亡,造成姐妹染色单体交换和畸变,导致心脏休克蛋白表达加强[3]。流行病学研究表明,在妊娠期内,尼古丁的暴露可以显著提高“婴儿猝死综合症”(sudden infant death syndrome,SIDS)的发病风险[4]。尼古丁暴露可以对胎儿的生长发育产生严重影响,包括直接的致畸效应以及出生后表现出来的“印记”效应(妊娠期内,在胎儿器官和系统发育的敏感期内,某些发生在胎儿生长环境中的改变可能会对胎儿留下永久性的“印记”,在其出生后会影响内分泌系统、代谢系统、心脑血管等的正常发育)[5]。
1.2 尼古丁污染的来源
自然环境和人类生活环境中尼古丁的污染主要来源于烟草及其废弃物、烟碱型农药、尼古丁橡胶、尼古丁药剂等[6],尤其以烟草废弃物污染最为严重。
我国是世界上最大的烟草生产国和消费国,烟草产量占世界总产量的35%,卷烟产量占世界总产量的32%,烟民数量占世界总量的20%,均位居世界第一,2007年,全行业利税达3 880亿元[7]。在巨大消费市场下,烟草行业逐步发展成为我国支柱产业之一,在国民经济中占有很高的比例。然而,在烟草行业带来巨大经济效益的同时,由于烟草大规模广泛种植、大量烟草制品的后期加工以及含尼古丁废弃物的不合理排放,给自然环境和人类健康带来巨大隐患。据统计,我国每年烟叶总产量约450万~500万t,其中约90万~150万t烟草制品下脚料被废弃。这些烟草废弃物中尼古丁平均含量高达18g·(kg干重)-1,是欧盟规定有毒有害烟草废弃物中尼古丁含量控制标准的36倍[8]。由于尼古丁具有极好的水溶性,在自然环境中不易分解,所以很容易造成水体的污染,甚至造成地下水污染,对自然环境和人类健康带来巨大威胁。因此,尼古丁污染处理受到研究人员的高度重视,微生物降解尼古丁也成为研究的热点。
2 尼古丁的降解
尼古丁含量是影响卷烟制品品质的重要因素,含量过高会导致烟气的刺激性强、卷烟吸味差、安全性差。目前,在烟草生产期间和后期加工过程中严格控制尼古丁的含量是减轻尼古丁污染的主要途径,其方法主要有物理化学法和微生物降解法。
物理化学法成本较高、副产物多,影响烟草其它组分。如袁淑霞等[9]研究发现,添加HNO3改性活性炭可吸附烟气中的尼古丁;王怡红等[10]研究发现,NaCl等无机盐对卷烟中的尼古丁具有去除效果;高兴斋等[11]用气浮加过滤,在调节池加以射流曝气,对卷烟厂废水进行处理,污染物去除率高达85%~95%,达到了废水排放标准。
微生物降解法选择性高、副产物少、高效、经济且易操作,微生物资源丰富,代谢具有多样性,所以得到了越来越多的关注。
3 尼古丁降解微生物
早在1927年,Batham[12]在土壤中加入尼古丁后发现硝酸盐的含量有所增加,他认为这是土壤微生物作用的结果,这是最早的关于尼古丁降解菌的报道。此后,越来越多的尼古丁降解菌被发现,包括细菌、放线菌及真菌。迄今为止,已发现的可降解尼古丁的微生物主要有:细菌中的假单胞菌属(如Pseudomonas putida S16[13]、Pseudomonas putida ZUTSKD[14])、节杆菌属(如Arthrobacter oxidans[15])、纤维单胞菌属[16](如Cellulomonas sp.)、苍白杆菌属[17](如Ochrobactrum intermedium)等;放线菌中的Streptomyces platense、Streptomyces griseus等;真菌中的Cunninghamella echinulata、Aspergillus oryzae[18]等。已发现的尼古丁降解菌中,大部分菌株可以以尼古丁为唯一碳源、氮源和能源生长。
假单胞菌是应用最广泛的一类尼古丁降解微生物。最早关于降解尼古丁假单胞菌的报道是Wada等[19]在土壤中分离得到的尼古丁降解菌Pseudomonas sp.No.41,并开始对该菌株进行代谢途径的研究。随 后,Pseudomonas convexa Pc1[20]、Pseudomonas putida S16[13]、Pseudomonas sp.HF-1[8]、Pseudomonas putida ZUTSKD[14]、Pseudomonas sp.Nic22[21]、Pseudomonas putida ZB-16A[22]、Pseudomonas sp.HZN6[23]、Pseudomonas sp.CS3[24]等假单胞菌属尼古丁降解菌陆续被报道。在这些假单胞菌中,耐受尼古丁能力最强的是P.putida ZB-16A和P.putida S16,可耐受尼古丁浓度高达6g·L-1,其次是P.putida ZUTSKD,可耐受尼古丁浓度为5.8g·L-1,其它菌株耐受尼古丁浓度一般在4g·L-1左右。P.putida S16降解能力最强,13h即可将4g·L-1的尼古丁完全降解。大部分假单胞菌降解尼古丁的最适温度为30℃、最适pH值为7.0。随着研究的深入,越来越多的可降解尼古丁的假单胞菌被发现,大部分属于Pseudomonas putida,同时也有一些新发现的菌种,如Pseudomonas stutzeri ZCJ[25],该菌株降解尼古丁的最适温度为37℃、最适pH值为7.5、耐受尼古丁浓度达4.5g·L-1,在培养过程中,培养液会有颜色的变化。
4 微生物降解尼古丁的代谢途径
关于微生物降解尼古丁的代谢途径目前还处于研究阶段。已报道并经证实的途径主要有3种:广泛存在于假单胞菌属(Pseudomonas)和刺孢小克汗霉菌属(Cunninghamella echinulata)的 吡 咯 途 径(pyrrolidine pathway)、广泛存在于节杆菌属(Arthrobacter)的吡啶途径(pyridine pathway)、广泛存在于烟草植物及真菌(Pellicularia filamentosa)中的脱甲基途径(Me pathway)[22,26-27]。
4.1 吡咯途径(图1)
吡咯途径从尼古丁的吡咯烷脱氢开始,生成N-甲基麦思明(N-methymyosmine),接着水化成假氧化尼古丁(pseudooxynicotine),脱掉甲胺生成3-琥珀酰吡啶(3-succinoylpyridine,SP),SP通过羟基化生成6-羟基-3-琥珀酰吡啶(6-hydroxy-3-succinoylpyridine,HSP),进而生成2,5-二羟基吡啶(2,5-dihydroxy-pyridine,DHP),最后吡啶环打开,完成吡咯途径的代谢。
4.2 吡啶途径(图2)
吡啶途径从尼古丁在尼古丁脱氢酶(NDH)作用下吡啶环第6位羟基化生成6-羟基尼古丁(6-hydroxynicotine)开始,随后在6-羟基尼古丁氧化酶(6-HNO)催化作用下生成6-羟基-N-甲基麦思明(6-hydroxy-N-methylmyosmine),6-羟基-N-甲基麦思明在酶6-HLNO或6-HDNO的催化作用下生成[6-羟基吡啶基-(3)]-甲胺丙基酮[N-methylaminopropyl-(6-hydroxypyridyl-3)-ketone],而后在酮脱氢酶(KDH)催化作用下生成[2,6-二羟基吡啶基-(3)]-甲胺丙基酮[N-methylaminopropyl-(2,6-dihydroxypyridyl-3)-ketone],随后经2,6-二羟基假氧化尼古丁水解酶(DHPONH)的催化生成2,6-二羟基吡啶(2,6-dihydroxypyridine)和γ-甲胺基丁酸(γ-N-methylaminobutyrate),最后γ-甲胺基丁酸经N-甲胺基丁酸氧化酶(MABO)的催化生成γ-胺基丁酸(γ-aminobutyrate),2,6-二羟基吡啶经2,6-二羟基吡啶-3-羟化酶(2,6-DHPH)的催化生成2,3,6-三羟基吡啶(2,3,6-trihydroxy-pyridine)。值得注意的是,尼古丁代谢吡啶途径中培养液经常会出现颜色的变化,可能是由于在代谢过程中,一些小分子的代谢中间产物聚合成蓝色聚合物——尼古丁蓝(nicotine blue)所致。
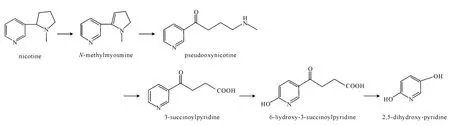
图1 尼古丁代谢的吡咯途径Fig.1 Pyrrolidine pathway of nicotine metabolism

图2 尼古丁代谢的吡啶途径Fig.2 Pyridine pathway of nicotine metabolism
4.3 脱甲基途径
脱甲基途径始于尼古丁吡咯烷的脱甲基化,先生成去甲基尼古丁(nornicotine),然后沿着去甲基尼古丁的代谢途径进一步代谢降解。
4.4 其它途径
随着尼古丁代谢途径研究的不断深入,一些新的尼古丁代谢途径及代谢产物被发现。利用GC-MS分析Pseudomonas sp.ZUTSKD对尼古丁的降解产物,检测到有可天宁、2,3′-二吡啶、3-(2,3,4-三氢-5-吡咯基)-吡啶和3-羧基-吡啶的生成,推断3-(2,3,4-三氢-5-吡咯基)-吡啶是由尼古丁经可天宁脱-CH3生成,这可能是尼古丁在Pseudomonas sp.ZUTSKD中降解代谢的初始步骤,不同于已知的微生物代谢尼古丁的途径。因此,推测Pseudomonas sp.ZUTSKD可能有着全新的尼古丁代谢途径[28]。
在对Pseudomonas sp.HF-1和Pseudomonas sp.Nic22尼古丁代谢中间产物进行分析时发现了可铁宁、去甲基尼古丁、麦思明等,在Pseudomonas sp.HF-1代谢尼古丁产物中还发现了尼古提林,通过HPLC、GC-MS、NMR和FTIR分析Pseudomonas plecoglossicida TND35降解尼古丁产物,发现了3种新的中间产物,这些均表明菌株中可能存在着其它的尼古丁代谢途径[22,29]。
5 结语
尼古丁不仅严重威胁人体健康,而且破坏土壤生态结构,扰乱生态平衡。微生物可以以尼古丁作为唯一的碳源、氮源和能源进行生长繁殖,最终将尼古丁通过不同的代谢途径降解成无害的物质。细菌中的假单胞菌属是降解尼古丁最广泛的微生物,其降解能力和耐受能力较强,具有较高的研究和实际应用价值。在未来的研究中,可以从酶学和分子生物学水平研究微生物代谢尼古丁的机理,找到关键酶及其基因,利用基因工程手段增强微生物对尼古丁的降解能力。
[1]NOVOTNY T E,ZHAO F.Consumption and production waste:Another externality of tobacco use[J].Tobacco Control,1999,8(1):75-80.
[2]CAMPAIN J A.Nicotine:Potentially a multifunctional carcinogen?[J].Toxicological Sciences,2004,79(1):1-3.
[3]AOSHIBA K,NAGAI A,YASUI S,et al.Nicotine prolongs neutrophil survival by suppressing apoptosis[J].Journal of Laboratory and Clinical Medicine,1996,127(2):186-194.
[4]RINTAHAKA P J,HIRVONEN J.The epidemiology of sudden infant death syndrome in Finland in 1969~1980[J].Forensic Science International,1986,30(2):219-233.
[5]BARKER D J,OSMOND C,GOLDING J,et al.Growth in utero,blood pressure in childhood and adult life,and mortality from cardiovascular disease[J].British Medical Journal,1989,298(6673):564-567.
[6]HEISHMA S J,TAYLOR R C,HENNINGFIELD J E.Nicotine and smoking:A review of effects on human performance[J].Experimental and Clinical Psychopharmacology,1994,2(4):345.
[7]黄英华,黄英梅,李宝香.中国烟草行业的市场定位及其发展方向的初步探讨[J].中国烟草学报,2008,14(6):57-61.
[8]RUAN A,MIN H,PENG X,et al.Isolation and characterization of Pseudomonas sp.strain HF-1,capable of degrading nicotine[J].Research in Microbiology,2005,156(5-6):700-706.
[9]袁淑霞,吕春祥,李永红,等.活性炭改性对滤嘴吸附性能的影响[J].太原理工大学学报,2008,38(6):509-513.
[10]王怡红,张征林,印秀堂,等.无机盐对香烟中焦油及尼古丁去除的研究[J].化工时刊,1999,13(1):17-20.
[11]高兴斋,刘育禾,钟巧云,等.用物化法处理卷烟厂综合废水的实践[J].环境科学研究,1998,11(5):55-57.
[12]BATHAM H N.Nitrification in soil[J].Soil Science,1927,24:187-200.
[13]WANG S N,XU P,TANG H Z,et al.Biodegradation and detoxification of nicotine in tobacco solid waste by aPseudomonas sp.[J].Biotechnology Letters,2004,26(19):1493-1496.
[14]ZHONG W,ZHU C,SHU M,et al.Degradation of nicotine in to-bacco waste extract by newly isolated Pseudomonas sp.ZUTSKD[J].Bioresource Technology,2010,101(18):6935-6941.
[15]SGUROS P L.Microbial transformation of the tobacco alkaloids I:Cultural and morphological characteristics of a nicotinophile[J].Journal of Bacteriology,1955,69:28-33.
[16]GEISS V L,GRAVELY L E,GREGORY C F.Process for reduction of nitrate and nicotine content of tobacco by microbial treatment:US,4557280[P].1985-12-10.
[17]袁勇军,陆兆新,黄丽金,等.烟碱降解细菌的分离、鉴定及其降解性能的初步研究[J].微生物学报,2005,45(2):181-184.
[18]MENG X J,LU L L,GU G F,et al.A novel pathway for nicotine degradation by Aspergillus oryzae 112822isolated from tobacco leaves[J].Research in Microbiology,2010,161(7):626-633.
[19]WADA E,YAMASAKI K.Degradation of nicotine by soil bacterial[J].Journal of the American Chemical Society,1954,76(1):155-157.
[20]THACKER R,RORVING O,KAHLON P,et al.NIC,A conjugative nicotine-nicotinate degradative plasmid in Pseudomonas convexa[J].Journal of Bacteriology,1978,135(1):289-290.
[21]CHEN C,LI X,YANG J,et al.Isolation of nicotine-degrading bacteriumPseudomonas sp.Nic22,and its potential application in tobacco processing[J].International Biodeterioration & Biodegradation,2008,62(3):226-231.
[22]LI H,LI X,DUAN Y,et al.Biotransformation of nicotine by microorganism:The case of Pseudomonas spp.[J].Applied Microbiology and Biotechnology,2010,86(1):11-17.
[23]QIU J,MA Y,CHEN L,et al.AsirA-like gene,sirA2,is essential for 3-succinoyl-pyridine metabolism in the newly isolated nicotine-degrading Pseudomonas sp.HZN6strain[J].Applied Microbiology and Biotechnology,2011,92(5):1023-1032.
[24]WANG H H,YIN B,PENG X X,et al.Biodegradation of nicotine by newly isolated Pseudomonas sp.CS3and its metabolites[J].Journal of Applied Microbiology,2012,112(2):258-268.
[25]ZHAO L,ZHU C,GAO Y,et al.Nicotine degradation enhancement by Pseudomonas stutzeri ZCJ during aging process of tobacco leaves[J].World Journal of Microbiology and Biotechnology,2012,28(5):2077-2086.
[26]EBERHARDT H J.The biological degradation of nicotine by nicotinophilic microorganisms[J].Beitragezur Tabakforschung International,1995,16(3):119-129.
[27]BRANDSCH R.Microbiology and biochemistry of nicotine degradation[J].Applied Microbiology and Biotechnology,2006,69(5):493-498.
[28]孙柯丹,朱晨静,钟卫鸿,等.一株尼古丁高效降解菌株的分离鉴定和降解特性[J].环境科学学报,2008,28(7):1294-1301.
[29]RAMAN G,MOHAN K N,MANOHAR V,et al.Biodegradation of nicotine by a novel nicotine-degrading bacterium,Pseudomonas plecoglossicida TND35and its new biotransformation intermediates[J].Biodegradation,2014,25(1):95-107.
