苋菜试管苗amaNACA2-like基因克隆及生物信息学分析
2014-01-14刘生财匡华琴谢礼洋赖钟雄
刘生财,匡华琴,谢礼洋,赖钟雄
(福建农林大学园艺植物生物工程研究所,福州 350002)
苋菜试管苗amaNACA2-like基因克隆及生物信息学分析
刘生财,匡华琴,谢礼洋,赖钟雄*
(福建农林大学园艺植物生物工程研究所,福州 350002)
以苋菜试管苗为材料,采用RT-PCR结合RACE技术进行amaNACA-2 like基因全长cDNA序列克隆,对其进行生物信息学分析。结果表明,苋菜amaNACA-2 like基因cDNA全长为896 bp,其中5'UTR 66 bp,3'UTR 216 bp,poly A尾长17 bp,并包含一个615 bp开放阅读框,编码205个氨基酸。GenBank登录号为KC588966。生物信息学分析表明,该蛋白是一种亲水性细胞质蛋白,包括10个磷酸化位点和8个功能位点;该蛋白属于NAC蛋白超级家族(NAC superfamily),包含NAC功能位点,同时包括一段UBA超家族结构域;其二级结构由α螺旋、无规则卷曲结构、延伸链和β转折组成;系统进化树分析表明苋菜amaNACA2-like蛋白独立形成一个分支,但与其他物种间亲缘关系较近,说明NACA 2-like蛋白在生物进化过程中比较保守。研究amaNACA-2 like基因克隆和生物信息学分析,可为深入研究其功能奠定基础。
苋菜;试管苗;amaNACA-2 like;克隆;生物信息学
核糖体是细胞质中不同环境条件下合成多肽的分子机器,核糖体上新合成的多肽与新生多肽关联复合体(Nascent polypeptide associated complex,NAC)结合可保护新合成的蛋白质不与不适宜的细胞质因素相互作用[1-3],NAC可直接和信号识别粒子(Signal recognition particle,SRP)互作保证新合成的蛋白质运转到内质网中,阻止非分泌蛋白运转[4-5],这表明NAC在保证蛋白质准确运转,维持生命正常活动中起重要作用[6]。NAC是一个由α和β两个亚基组成的异源二聚体,是在高等真核生物中鉴定的新生多肽链的核糖体关联因子[2,7]。在转录调控和细胞分化过程中,NAC两个亚基表达不均衡,暗示这两个亚基有其各自功能[8-10]。α亚基具有转录激活及与DNA、rRNA和tRNA结合功能[11];而β亚基可以和RNA聚合酶II型结合,与转录起始有密切关系[12],在植物中有广泛研究[13-14]。有关α亚基的研究主要是在动物、微生物及人类中较多[10,15-17],对于植物鲜有报道。
苋菜(Amaranthus tricolorL.)属于C4一年生草本植物,离体培养周期短,大约60 d开花结籽,可作为研究植物生长发育的理想材料。NACA在植物生长发育过程中调控作用机制尚无报道。因此,本研究中以苋菜试管苗为试验材料,通过克隆nascent polypeptide-associated compleχ subunit alpha-likeprotein 2-like基因(amaNACA2-like),并对所获得的基因进行生物信息学分析,为探讨NACA基因对苋菜试管苗乃至其他植物的转录调控机制奠定基础。
1 材料与方法
1.1 材料
以福建农林大学园艺植物生物工程研究所提供的苋菜(Amaranthus tricolorL.syn.A.mangostanus)无菌试管苗为试验材料。
1.2 方法
1.2.1 苋菜试管苗总RNA提取及质量检测
使用多糖多酚植物总RNA快速提取试剂盒(百泰克公司)提取苋菜试管苗总RNA。利用琼脂糖凝胶电泳检测RNA完整性,利用超微量分光光度计(Thermo公司)测定RNA纯度和浓度。
1.2.2 苋菜试管苗amaNACA2-like基因cDNA合成
采用Fermentas公司RevertAidTMFirst-Strand cDNA Synthesis Kit试剂盒合成保守区域及3'RACE cDNA第一链,采用Clontech公司Super SMARTerTMRACE cDNA Amplificaion Kit试剂盒合成5'RACE cDNA第一链。
1.2.3 苋菜amaNACA2-like基因引物设计及PCR扩增
根据NCBI登陆的拟南芥、烟草、蓖麻、苜蓿、大豆等NACA基因序列比对结果,根据保守区域设计上下游引物;3'RACE特异引物是以克隆得到的苋菜amaNACA2-like基因保守区设计两条正向引物;5'RACE特异引物是在保守区和3'端拼接后的相对特异区域设计的2条反向引物;根据拼接得到的苋菜amaNACA2-like基因的全长设计ORF引物。委托北京六合华大基因科技股份有限公司合成引物。本试验所用引物见表1。根据扩增不同的目的片段,设定相应的PCR扩增程序,PCR扩增体系为25 μL。
1.2.4 目的片段的回收、克隆及测序
利用本实验室琼脂糖凝胶DNA回收试剂盒,以pMD18-T作为载体,转化到感受态细胞DH5α中,挑取单克隆进行摇菌培养,通过菌液PCR初步验证阳性克隆,将菌液送交北京六合华大基因科技股份有限公司测序。
1.2.5 苋菜amaNACA2-like基因序列生物信息学分析
使用DNAMAN 7.0进行序列拼接及氨基酸序列推导;使用MEGA 5.0软件构建不同物种间NACA的进化树;使用在线软件对该基因进行生物信息学分析。
2 结果与分析
2.1 苋菜amaNACA2-like基因全长的克隆
通过对苋菜amaNACA2-like基因cDNA进行PCR扩增(见图1),得到片段大小为381 bp的保守序列,299 bp的3'端序列,383 bp的5'端序列。通过DNAMAN 7.0软件进行序列拼接获得全长cDNA,经过开放阅读框验证及分析,起始密码子ATG上游-3位核苷酸为A,+4位为G,为典型的Kozak结构,3'UTR有加尾信号“GATAA”,说明该基因为完整的一条基因(登陆号为KC588966)。经分析显示该基因全长为896 bp,其中5'UTR 66 bp,3' UTR 216 bp,poly A尾长17 bp,并包含一个615 bp的开放阅读框(见图2),编码205个氨基酸。

表1 试验所用引物及其序列Table 1 Primers and sequences were used in the study

图1 苋菜amaNACA2-like基因cDNA扩增产物电泳图Fig.1 Amplification electrophoretogram ofAmaranthus tricolorL.amaNACA2-likecDNA
2.2 苋菜amaNACA2-like基因生物信息学分析
2.2.1 苋菜amaNACA2-like编码的蛋白质理化性质预测与分析
利用ProtParam对苋菜amaNACA2-like编码的蛋白质理化性质进行预测与分析。结果显示,该蛋白分子式为C1157H1845N305O388S9,由3 704个原子构成,包括242个氨基酸,相对分子质量为26.5247 ku,理论等电点(pI)4.43,带正电的氨基酸有25(Arg+Lys)个,带负电的氨基酸有49(Asp+Glu)个,总平均疏水性为-0.386,属于亲水蛋白,但不是稳定蛋白(不稳定系数为53.26)。
2.2.2 苋菜amaNACA2-like蛋白保守结构域的预测与分析
根据NCBI-CDS在线分析工具对苋菜ama-NACA 2-like蛋白的结构域进行分析,该蛋白属于NAC蛋白超级家族(NAC superfamily),包含NAC功能位点。另通过比对预测该蛋白序列包括一段UBA超家族结构域(见图3),这与大豆氨基酸序列预测结果相同。
2.2.3 苋菜amaNACA2-like蛋白质信号肽、跨膜区及亚细胞定位的预测
采用在线分析工具SignaIP 4.1 Server(http://www.cbs.dtu.dk/services/SignalP/)对苋菜amaNACA2-like蛋白进行分析,结果表明该蛋白没有明显的信号肽,说明该蛋白可能在胞质中起作用。利用跨膜结构分析在线分析软件TMpred(http://vit-embnet.unil. ch/software/TMPRED-form.html),对苋菜amaNACA2-like蛋白进行跨膜结构预测,结果表明其多肽链上不存在由内向外或由外向内的螺旋,因此推测该蛋白不属于跨膜蛋白,有可能在细胞质中起作用。应用PSORT对该蛋白的亚细胞定位进行预测,结果表明该蛋白最可能定位于细胞质中,可能性为0.650。通过以上分析,推断苋菜amaNACA2-like属于一种胞质蛋白。

图2 苋菜amaNACA2-like基因cDNA全长(彩图)Fig.2 Full-length sequence ofAmaranthus tricolorL.amaNACA2-likecDNA
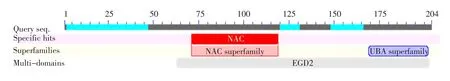
图3 NCBI中amaNACA2-like蛋白氨基酸序列的保守结构域检索Fig.3 Search for conserved domain ofamaNACA2-likeprotein amino acid sequence ofAmaranthus tricolorL.by NCBI
2.2.4 苋菜amaNACA2-like蛋白质磷酸化位点及功能位点预测
蛋白质磷酸化作用在细胞信号传递过程中占有极其重要地位。利用NetPhos 2.0 Serve在线磷酸化位点分析工具(http://www.cbs.dtu.dk/services/ NetPhos/)对苋菜amaNACA2-like蛋白的磷酸化位点进行预测。从图4中可以看出,整个多肽链存在10个氨基酸磷酸化位点(>0.5),其中丝氨酸(Ser)可能发生磷酸化的有7个,苏氨酸(Thr)可能发生磷酸化的有2个,酪氨酸(Tyr)可能发生磷酸化的有1个。可以看出,丝氨酸磷酸化在苋菜ama-NACA 2-like蛋白中占主导地位,推测丝氨酸磷酸化可能在苋菜amaNACA2-like蛋白行使功能中有重要作用。

图4 苋菜amaNACA2-like蛋白磷酸化修饰预测Fig.4 Predicted phosphorylation sites in amino acid sequence ofamaNACA2-like
蛋白质功能位点和相互作用位点的预测对于认识蛋白质功能机制有重大意义。通过Predict-Protein在线软件分析(https://www.predictprotein.org// submit-basic.php)。结果表明amaNACA2-like蛋白存在3个蛋白激酶C磷酸化位点:SEK、SRK、TIK,3个酪蛋白激酶II磷酸化位点:SRSE、SKPD、TGVE,2个酰基化位点:GSAEGT、GVSRSK。
2.2.5 苋菜amaNACA2-like蛋白质卷曲以及保守结构域预测
利用Coils分析工具,对amaNACA2-like形成卷曲螺旋的倾向性进行预测,以window=14、21和28为参数,按照几率>50%就可形成螺旋的规则,在没有权重和有权重情况下的分析结果一致的区段很有可能形成卷曲螺旋,3种参数及有无权重的分析结果显示:苋菜amaNACA2-like在windows= 14时极易形成卷曲螺旋,在windows=21时可能形成卷曲螺旋,而在windows=28时不能形成卷曲螺旋结构(见图5)。

图5 苋菜amaNACA2-like氨基酸序列卷曲螺旋的预测Fig.5 Predicted amino acid sequence of the winded helix ofamaNACA2-like
2.2.6 苋菜amaNACA2-like蛋白质二级结构与三级结构预测分析
利用在线分析工具PHD(http://npsa-pbil.ibcp.fr/ cgi-bin/npsa-automat.pl?page=/NPSA/npsa-phd.html)对苋菜amaNACA2-like蛋白进行二级结构预测。amaNACA2-like蛋白二级结构以α螺旋结构为主,所占比例为42.65%,其次为无规则卷曲结构37.25%,而延伸链和β转折占的比例较少,分别为13.24%和6.86%(见图6)。
利用SWISS-MODEL在线软件预测该基因编码蛋白的三级结构,发现该蛋白由β折叠和无规则卷曲构成(见图7)。
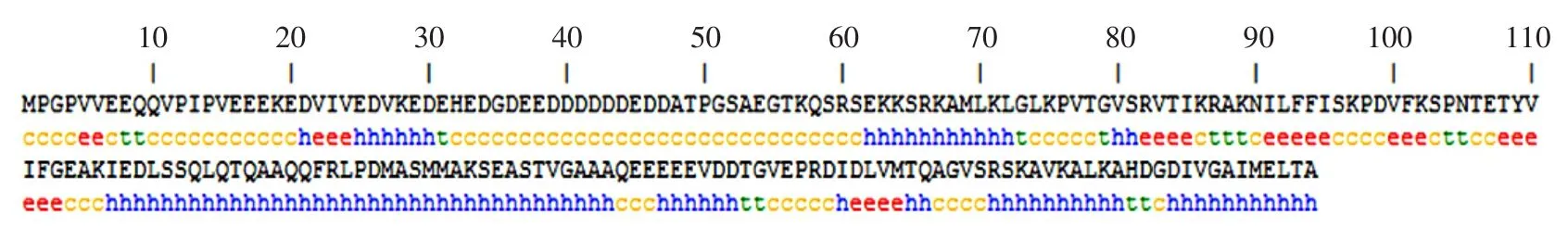
图6 amaNACA2-like蛋白序列二级结构PHD预测结果Fig.6 Secondary structure prediction by PHD foramaNACA2-like

图7 amaNACA2-like蛋白的三维结构预测Fig.7 Three-dimensional structure ofamaNACA2-likebasing on SWISS-MODEL
2.2.7 苋菜amaNACA2-like蛋白系统进化树构建
本研究通过对苋菜amaNACA2-like基因编码的氨基酸序列在线BLAST的结果,选取拟南芥、蓖麻、葡萄、玉米、苜蓿、烟草等其他9种植物的amaNACA2-like基因编码的氨基酸序列,运用MEGA 5.0软件中的邻位相连法(Neighbor-Joining,NJ),将Bootstrap检验值设置为1 000,建立其编码区氨基酸序列之间的系统关系树(见图8)。
从图8中可以清楚看出,苋菜amaNACA2-like蛋白独立形成一个分支,但与其他物种间亲缘关系比较近,可信度达77%,特别是拟南芥间的可信度达100%。说明NACA 2-like蛋白在生物进化过程中比较保守。

图8 苋菜amaNACA2-like系统进化树Fig.8 Phylogenetic tree foramaNACA2-like
3 讨论与结论
新生多肽关联复合体是一个由αNAC和βNAC组成NAC的异源二聚体,可能是第一个与核糖体产生的新生肽链相结合的细胞质蛋白,在蛋白质折叠和运输早期起关键作用[18],保证新生链进入内质网中[19]。通过对保守结构域及系统进化树研究分析,本研究获得苋菜NACA基因序列。通过对NACA蛋白质信号肽、跨膜区及亚细胞定位进行预测,结果表明其在细胞质中起到作用,可能属于细胞质蛋白,暗示其在蛋白正确转运到内质网中可能起作用,但还需进一步研究。大量研究表明,NACA可能在动物、微生物或者人类的转录过程中起作用[17-20]。本研究进行的amaNACA-2 like基因克隆和生物信息学分析可为深入研究其功能奠定基础。
[1]Panasenko O O,David F P,Collart M A.Ribosome association and stability of the nascent polypeptide-associated complex is dependent upon its own ubiquitination[J].Genetics,2009,181(2): 447-460.
[2]Wiedmann B,Sakai H,Davis T A,et al.A protein complex required for signal-sequence specific sorting and translocation[J]. Nature,1994,370:434-440.
[3]Rospert S,Dubaquie Y,Gautschi M.Nascent-polypeptide-associated complex[J].Cell Mol Life Sci,2002,59(10):1632-1639.
[4]Lauring B,Kreibich G,Wiedmann M.The intrinsic ability of ribosomes to bind to endoplasmic reticulum membranes is regulated by signal recognition particle and nascent-polypeptide-associated complex[J].PNAS,1995,92(21):9435-9439.
[5]Moller I,Jung M,Beatrix B,et al.A general mechanism for regulation of access to the translocon:Competition for a membrane attachment site on ribosomes[J].PNAS,1998,95(23):13425-13430.
[6]Karan R,Subudhi P K.Overexpression of a nascent polypeptide associated complex gene(SabNAC)of Spartina alterniflora improves tolerance to salinity and drought in transgenic Arabidopsis[J].Biochemical and Biophysical Research Communications, 2012,424:747-752
[7]Huh S U,Kim K J,Paek K H.Capsicum annuum basic transcription factor 3(CaBtf3)regulates transcription of pathogenesis-relatedgenesduringhypersensitiveresponseupon tobacco mosaic virus infection[J].Biochem Biophys Res Commun, 2012,417(2):910-917.
[8]Quelo I,Hurtubise M,St-Arnaud R.Alpha NAC requires an interaction with c-Jun to exert its transcriptional coactivation[J]. Gene Expr,2002,10(5-6):255-262.
[9]Akhouayri O,Quelo I,St-Arnaud R.Sequence-specific DNA binding by the alpha NAC coactivator is required for potentiation of c-Jun-dependent transcription of the osteocalcin gene[J].Mol Cell Biol,2005,25(9):3452-3460.
[10]Lopez S,Stuhl L,Fichelson S,et al.NACA is a positive regulator of human erythroid-cell differentiation[J].J Cell Sci,2005,118 (8):1595-1605.
[11]Beatrix B,Sakai H,Wiedmann M.The α and β subunit of the nascent polypeptide-associated complex have distinct functions [J].The Journal of Biological Chemistry,2000,275(48):37838-37845.
[12]Markesich D C,Gajewski K M,Nazimiec M E,et al.Bicaudal encodes the Drosophila beta NAC homolog a component of the ribosomal translational machinery[J].Development,2000,127: 559-572.
[13]李广旭.受水稻白叶枯病菌诱导的水稻转录因子基因OsBTF3的功能分析[D].北京:中国农业科学院,2009.
[14]张中保,张登峰,李会勇,等.玉米干旱诱导表达基因ZmBTF3b的克隆与表达分析[J].中国农业科学,2010,43(16):3280-3287.
[15]Munz B,Wiedmann M,Lochmuller H,et al.Cloning of novel injury-regulated genes.implications for an important role of the muscle-specific protein skNAC in muscle repair[J].J Biol Chem, 1999,274:13305-13310.
[16]Reimann B,Bradsher J,Franke J,et al.Initial characterization ofthenascent polypeptide-associated complex in yeast[J]. Yeast,1999,15:397-407.
[17]Li H Q,Willianm R R,Du S J.skNAC(skeletal Naca),a musclespecific isoform of Naca(nascent polypeptide-associated complex alpha),is required for myofibril organization[J].The FASEB Journal,2009,23:1-13.
[18]Wang S,Sakai H,Wiedmann M.NAC covers ribosome-associated nascent chains thereby forming a protective environment for regions of nascent chains just emerging from the peptidyl transferase center[J].J Cell Biol,1995,130(3):519-528.
[19]Moller I,Beatrix B,Kreibich G,et al.Unregulated exposure of the ribosomal M-site caused by NAC depletion results in delivery of non-secretory polypeptides to the Sec61 complex[J].FEBS Lett, 1998,441(1):1-5.
[20]Moreau A,Yotov W V,Glorieux F H,et al.Bone-specific expression of the alpha chain of the nascent polypeptideassociated complex,a coactivator potentiating c-Jun-mediated transcription[J].Mol Cell Biol,1998,18:1312-1321.
Cloning and bioinformatics analysis ofamaNACA2-likefromin vitro
plantlets inAmaranthus tricolorL.
LIU Shengcai,KUANG Huaqin,XIE Liyang,LAI Zhongxiong
(Institute of Horticultural Biotechnology,Fujian Agriculture and Forestry University, Fuzhou 350002,China)
The full-lengthamaNACA2-likecDNA sequence was cloned using RT-PCR and RACE techniques and the bioinformatics was analyzed fromin vitroplantlets ofAmaranthus tricolorin this experiment.The full-length cDNA sequenceamaNACA2-likewas 896 bp in length,containing an open reading frame of 615 bp(encoding 205 animo acids),flanked by 66 bp 5'UTR and 216 bp 3'UTR that contained a 17 bp polyA.The accession number was KC588966 in GenBank.Bioinformatics analysis showed that the protein was hydrophilic cytoplasmic protein,with 10 phosphorylation sites and eight functional sites.The protein belongs to NAC superfamily with a NAC functional site.In addition,a UBA superfamily domain was contained in the sequence.The secondary structure was consisted of alphahelix,coil,strand chain and beta turn.The phylogenetic tree indicated thatamaNACA2-likeprotein constituted a branch alone,but had a close relationship with other species,which indicated NACA 2-likeprotein possesses high conservatism during species evolution.This study was helpful for further study of plantNACA2-likefunctions.
Amaranthus tricolor;in vitroplantlets;amaNACA2-like;cloning;bioinformatics
S572
A
1005-9369(2014)01-0083-07
2013-08-08
高等学校博士学科点专项新教师类科研基金项目(20123515120009);福建省自然科学基金项目(2013J05045);福建省教育厅A类项目(JA12098);福建农林大学园艺学博士后基金项目(132300102);福建农林大学园艺学院青年学术骨干培养基金项目(FAFU2012 YYPY10)
刘生财(1980-),男,讲师,博士,研究方向为园艺植物生物技术与遗传资源。E-mail:1215698900@qq.com
*通讯作者:赖钟雄,研究员,博士生导师,研究方向为园艺植物生物技术与遗传资源。E-mail:Laizx01@163.com
时间2014-1-9 20:19:46[URL]http://www.cnki.net/kcms/detail/23.1391.S.20140109.2019.006.html
刘生财,匡华琴,谢礼洋,等.苋菜试管苗amaNACA2-like基因克隆及生物信息学分析[J].东北农业大学学报,2014,45(1): 83-89.
Liu Shengcai,Kuang Huaqin,Xie Liyang,et al.Cloning and bioinformatics analysis ofamaNACA2-likefromin vitroplantlets inAmaranthus tricolorL.[J].Journal of Northeast Agricultural University,2014,45(1):83-89.(in Chinese with English abstract)
