低温胁迫下冬小麦东农冬麦1号分蘖节相关EST-SSR信息分析
2014-01-14苍晶范博宋扬许贺牟永潮于晶张达王军虹孟婧朱祥春
苍晶,范博,宋扬,许贺,牟永潮,于晶,张达,王军虹,孟婧,朱祥春
(东北农业大学生命科学学院,哈尔滨 150030)
低温胁迫下冬小麦东农冬麦1号分蘖节相关EST-SSR信息分析
苍晶,范博,宋扬,许贺,牟永潮,于晶,张达,王军虹,孟婧,朱祥春
(东北农业大学生命科学学院,哈尔滨 150030)
利用抑制消减杂交技术构建东农冬麦1号分蘖节低温胁迫相关基因的cDNA正反文库,获得表达序列标签EST(Expressed sequence tag),经分析和前期处理后拼接得到736条非重复序列(Unigenes)。运用生物信息学方法对低温胁迫下诱导的冬小麦东农冬麦1号分蘖节相关EST-SSRs分布频率和碱基重复特点进行归纳分析。结果表明,736条Unigenes共检索到59个EST-SSRs,EST-SSRs出现频率为8.02%。EST-SSRs包括24种重复单元,其主要以三核苷酸和二核苷酸重复基元为主,二者分别占总SSRs 47.46%和44.07%,在EST中出现频率分别为3.80%和3.53%,其中以二核苷酸CA和AG重复基元为主,三核苷酸重复基元以AAC重复基元所占比例最高。为冬小麦EST资源开发和抗寒基因挖掘奠定基础。
冬小麦;东农冬麦1号;低温胁迫;分蘖节;EST;SSR
东农冬麦1号是首例能在黑龙江省高寒地区越冬的强抗寒冬小麦品种,可耐-30℃低温,其返青率可达85%以上。
EST(Expressed sequence tag)和cDNA中均含有SSR,基于EST的SSR(Simple sequence repeat)标记是近年发展起来的新型分子标记,与传统SSR相比,可反映基因的编码部分,直接获得基因表达信息,为功能基因提供绝对标记,使得对决定重要性状的基因进行直接鉴定成为可能,从而强化SSR标记在遗传研究中的应用。在众多分子遗传标记方法中,EST-SSR方法具备成本低且简单、有效,重复性好、多态性高等优点[1]。有关小麦ESTSSR分析已有报道[2],但关于冬小麦抗寒相关的EST-SSR尚未见报道。本研究对低温胁迫下东农冬麦1号分蘖节相关的EST-SSR进行分析,为挖掘冬小麦抗寒基因、辅助抗寒品种选育提供生物信息学基础。探究东农冬麦1号高抗寒的分子机制及其遗传背景,对于进一步分子育种,获得更适宜的优质抗寒新材料,具有理论意义和应用前景。
1 材料与方法
1.1 东农冬麦1号分蘖节相关EST来源
东农冬麦1号分蘖节相关EST来源于东北农业大学植物生理生化与分子生物学研究室构建的低温胁迫下东农冬麦1号分蘖节的抑制消减杂交文库。
1.2 东农冬麦1号分蘖节相关EST的前期处理
从培养基上挑取白色菌斑进行序列测定。挑取正反文库的阳性克隆进行序列测定,成功回收789个EST,序列长度介于103~844 bp之间,平均355bp。
1.3 东农冬麦1号分蘖节相关EST的聚类与去冗余
在处理过程中可能存在多个EST代表相同转录产物的情况,这可能是由于特定组织的EST序列会代表随机取样的各种转录产物。为避免产生冗余ESTs以及使聚类组成获得质量高的长序列,可使用程序对聚类进行拼接。使用CAP3程序获得736条unigenes,占全部序列的93.28%。
1.4 东农冬麦1号分蘖节EST序列的信息分析
运用SSRI软件http://www.gramene.org/db/searches/ssrtool在线搜索EST-SSR。搜索标准为:二核苷酸、三核苷酸、四核苷酸、五核苷酸、六核苷酸重复序列的重复次数大于或等于5。
2 结果与分析
2.1 东农冬麦1号分蘖节相关EST中SSR的出现频率与分布
东农冬麦1号分蘖节相关的EST-SSR类型丰富,二核苷酸至六核苷酸均可被检索到,但各种重复类型所占比例和出现频率各不相同。其中二核苷酸和三核苷酸是主要的重复类型,分别占总SSR的44.07%和47.46%,四核苷酸、五核苷酸和六核苷酸所占总SSR比例非常小,分别为0.41%、0.14%和0.14%,三者在EST中的出现频率均未超过1%(见表1)。

表1 东农冬麦1号分蘖节相关EST-SSRs出现频率Table 1 Frequency of EST-SSRs occurred in tillering nodes of Dongnongdongmai 1
2.2 东农冬麦1号分蘖节相关EST-SSR中重复基元的特点
在检索到的59个EST-SSR中,共包括24种核苷酸重复基元。其中三核苷酸重复基元类型最为丰富,共11种,占总核苷酸基元类型的45.83%;其次是二核苷酸基元类型,共8种,占总核苷酸基元类型的33.33%;四核苷酸、五核苷酸和六核苷酸的基元类型均为1种。二核苷酸基元类型主要以CA为主,AG次之,TC、AT依次减少,TA、GA、CT、GT检索数量均为1(见表2)。三核苷酸基元类型主要以AAC比例最高,占三核苷酸总基元含量的28.75%;其次是ACA,占三核苷酸总基元含量的17.86%。其他各基元类型所占比例较分散,由于基元类型较多,各基元所占比例均较低。四、五、六核苷酸基元类型共有5种,每一种基元数目都较少,因此,在整个EST中出现频率也很低。
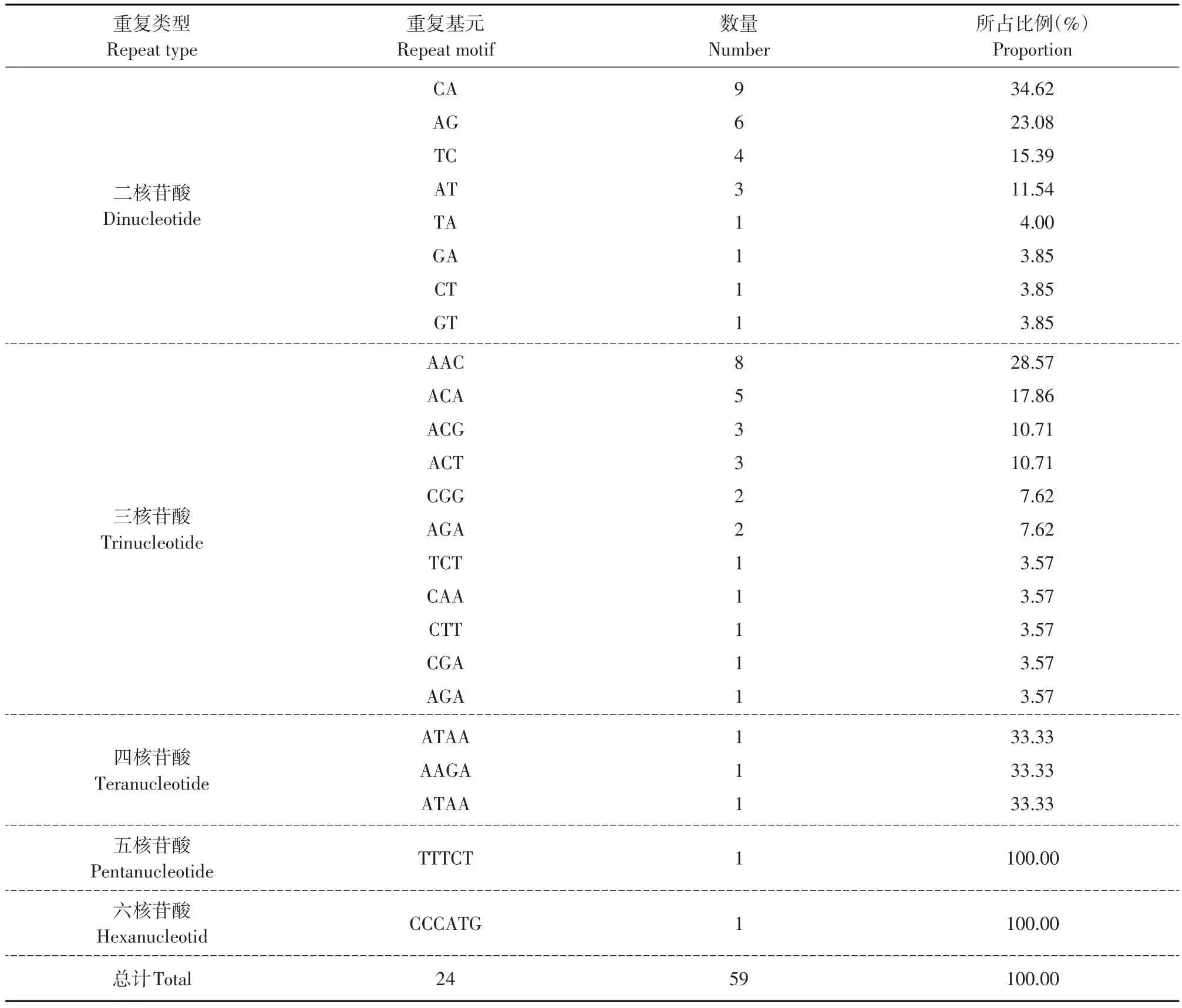
表2 东农冬麦1号分蘖节EST-SSRs重复类型的出现情况Table 2 Repeat type occurred in EST-SSRs of tillering nodes of Dongnongdongmai 1
2.3 东农冬麦1号分蘖节相关EST-SSR中重复基元的分布特点
东农冬麦1号分蘖节相关EST-SSR多态性主要是由于重复基元的重复次数、重复类型以及SSR长度不同造成。
如表3所示,由于东农冬麦1号分蘖节相关EST-SSR重复基元的重复次数不同致使其EST-SSR呈现多态性,从而获得分蘖节相关EST-SSR不同数目。对所获得的EST进行SSR检索时,以重复基元最低重复次数5为标准,进行检索共获得9种重复次数。最高重复次数达到13次重复。重复5次获得EST-SSR数目最多,共29个,占总SSR数49.15%,重复6次获得EST-SSR数目为13个,占总SSR数22.03%,重复7次获得EST-SSR数目为7个,占总SSR数11.86%。其他重复次数获得ESTSSR较少。

表3 东农冬麦1号分蘖节相关EST-SSRs重复次数的出现情况Table 3 Number of repeat occurred in EST-SSR of tillering nodes of Dongnongdongmai 1
3 讨论与结论
对低温胁迫下构建的东农冬麦1号分蘖节的抑制消减杂交文库,经处理成功得到789条EST,经分析和前期处理后拼接得到736条序列中共检索到ESR-SSR 59个,出现频率为8.02%,与其他已研究物种的EST-SSR出现频率相近。例如水稻4.7%[3]、普通小麦5.4%[4]、银杏5.97%[5]、百合5.98%[6]、黄瓜7.47%[7]、蝴蝶兰7.65%[8]、白菜10.34%[9]等。其出现频率均低于15%。但与某些物种反差较大,例如西瓜53.4%[10]。造成EST-SSR出现频率不同的原因主要有:SSR检索时长度的不同、物种间SSR间的真实差异、同一物种间EST来源于不同组织和器官、不同研究收集的EST数量不同、现有EST数据不足以覆盖基因组等。
东农冬麦1号分蘖节相关的EST-SSR类型主要以二核甘酸和三核苷酸为主,分别占总SSR的44.07%和47.46%,这与大多数报道一致。但由于物种的不同,重复基元会存在差异[2]。东农冬麦1号分蘖节相关EST-SSR分析表明,三核苷酸重复基元略高于二核苷酸重复基元,这与梅[11]、甘蔗[12]、烟草[13]、西洋参[14]、中国板栗[15]、花生栽培种[16]等研究结果一致。但与荔枝[17]等研究结果相反。在三核苷酸重复基元中,主要以AAC和ACA重复类型为主,分别占三核苷酸重复基元的28.57%和17.86%,不同植物的三核苷酸重复基元变化较大。本研究表明,三核苷酸主要是AAC和ACA类型为主,而在水稻和高粱中则以GGC和GCC为主,鹅掌楸则以AAG和CTT为主要重复基元[18]。二核苷酸重复基元主要以CA和AG重复类型居多,分别占二核苷酸重复基元的34.62%和23.08%,研究表明,这些占优势的重复基元可能与其编码相应蛋白质的使用频率较高有关[19]。GC重复基元是一种难以检索到的重复基元,本文同样未检索到GC重复基元。Rota曾提出若EST-SSR数目足够庞大和无偏倚性,即在于理想状态下,由4种不同碱基随机组合,以数学概率统计将会产生2种单核苷酸、4种二核苷酸、10种三核苷酸、33种四核苷酸、102种五核苷酸和350种六核苷酸基本重复基元类型[2]。本研究对EST-SSR的分析表明,很多重复基元出现频率很低,例如四、五、六核苷酸的重复基元占全部EST-SSR的比例均不足1%,分别为0.41%、0.14%和0.14%。这些相对高级基元的核苷酸重复都不是东农冬麦1号分蘖节相关的EST-SSR重复基元的主导类型,表明其EST-SSR确实存在一定偏差。
结果分析表明,东农冬麦1号分蘖节相关EST-SSR中,重复的基元类型不是很多,共24种,其中三核苷酸基元重复类型最为丰富达到11种,占核苷酸基元重复类型的45.83%,二核苷酸重复类型8种,四核苷酸3种,五核苷酸1种,六核苷酸1种。
冬小麦EST序列数量较大,本研究只针对低温胁迫下东农冬麦1号分蘖节相关EST-SSR信息分析,不足以反映冬小麦EST-SSR分布频率特点及规律。但研究结果为进一步开发与东农冬麦1号分蘖节相关EST-SSR的信息位点奠定理论基础,随着研究深入,对冬小麦总EST-SSR进行全面生物信息学分析,加快冬小麦EST资源开发和利用、分子标记、遗传资源评价、遗传作图、特定形状的辅助选择及育种等方面具有重要意义。
[1]李小白,崔海瑞,张明龙.EST分子标记开发及在比较基因组学中的应用[J].生物多态性,2006,14(6):541-547.
[2]Rota L R,Kantety R V,Yu J K.Nonrandom distribution and frequencies of genomic and EST-derived microsatellite markers in rice,wheat and barley[J].BMC Genomics,2005,6(1):23.
[3]Cordeiro G V,Rota M L,Matthews D E,et al.Data mining for simple sequence repeats in expressed sequence tags from barley, maize,rice,sorghum and wheat[J].Plant Molecular Biology,2002, 48:501-510.
[4]Gupta P K,Rustgi S,Sharma S,et al.Transferable EST-SSR markers for the study of polymorphism and genetic diversity in bread wheat[J].Molecular Genetics and Genomics,2003,270: 315-323.
[5]樊洪泓,李廷春,李正鹏,等.银杏EST序列中微卫星的分布特征[J].基因组学与应用生物学,2009,28(5):869-873.
[6]杨素丽,明军,刘春,等.基于EST信息的百合SSR标记的建立[J].园艺学报,2008,35(7):1069-1074.
[7]王丽娟,孙彩玉,秦智伟.黄瓜抗霜霉病相关ERT-SSR信息分析[J].东北农业大学学报,2011,42(10):52-56.
[8]张君毅,陈瑞凤.蝴蝶兰EST-SSRs分析[J].植物生理学通讯, 2010,46(6):559-563.
[9]忻雅,崔海瑞,卢美贞,等.白菜EST-SSR信息分析与标记的建立[J].园艺学报,2006,33(3):549-554.
[10]冯建明,郭绍贵,吕桂云,等.西瓜抗枯萎病相关EST-SSR的信息分析[J].华北农学报,2009,24(3):87-91.
[11]上官凌飞,李晓颖,宋长年,等.梅EST-SSR标记的开发及利用[J].西北植物学报,2010,30(9):1766-1772.
[12]闫学兵,阙友雄,许莉萍,等.甘蔗EST序列的SSR信息分析[J].热带作物学报,2010,31(9):1497-1501.
[13]胡重怡,蔡刘体,陈兴江.烟草ESTs资源的SSR信息分析[J].生物技术通讯,2009(7):81-85.
[14]杨维泽,金航,赵振玲,等.西洋参EST资源的SSR信息分析[J].西南农业学报,2011,24(1):275-278.
[15]向闱,宁祖林,康明,等.中国板栗EST-SSR信息分析及其通用性[J].热带亚热带植物学报,2010,18(6):665-669.
[16]梁炫强,洪彦彬,陈小平,等.花生栽培种EST-SSRs分布特征及应用研究[J].作物学报,2009,35(2):246-254.
[17]孙清明,马文朝,马帅鹏,等.荔枝EST资源的SSR信息分析及EST-SSR标记开发[J].中国农业科学,2011,44(19):4037-4049.
[18]Xu M,Sun Y G,Li H G.EST-SSRs development and paternity analysis for Liriodendronspp.[J].NewForests,2010,40(3):361-382.
[19]范三红,郭蔼光,单丽伟,等.拟南芥基因密码子偏爱性分析[J].生物化学与生物物理进展,2003,30(2):221-225.
Analysis of EST-SSR information in tillering nodes of winter wheat Dongnongdongmai1 under low temperature stress
CANG Jing,FAN Bo,SONG Yang,XU He,MU Yongchao,YU Jing,ZHANG Da,WANG Junhong,MENG Jing,ZHU Xiangchung
(School of Life Sciences,Northeast Agricultural University,Harbin 150030,China)
Expressed sequence tags(EST)were obtaineded from positive and negative cDNA library of low temperature stress related genes in Dongnongdongmai1,which was constructed by suppression subtractive hybridization technique.After analysis and pretreatment, the EST were assembled and 736 unigenes were acquired.The distribution frequency and the characteristics of base-repeat of EST-SSRs in tillering nodes of Dongnongdongmai1 under low temperature were summarized and analyzed using the bioinformatics method.The results showed that All of 56 EST-SSRs distributed in 736 unigenes.The frequency of EST-SSRs occurences was 8.02%.These EST-SSRs were composed of 24 kinds repeat motifs and they were basically trinucleotide or dinucleotide repeat motifs.In the total SSRs,the trinucleotide repeat motifs accounted for 47.46%and the dinucleotide repeat motifs accounted for 44.07%.In the total EST frequency,the trinucleotide repeat motifs accountedfor 3.80%and the the dinucleotide repeat motifs accounted for 3.53%.The repeat motif on CA and AG in dinucleotide repeat motifs were the most common motif,and The repeat motif on AAC in trinucleotide repeat motifs accounted for the greatest proportion.All these conclusions will establish the theoretic foundationfordevelopment EST resourceof winterwheat andcoldresistancegeneexcavation.
winter wheat;Dongnongdongmai1;low temperature stress;tillering nodes;EST;SSR
S512.1+1
A
1005-9369(2014)01-0001-05
2012-01-12
高等学校博士学科点专项科研基金课题(20112325110003);黑龙江省教育厅科学技术研究项目;黑龙江省教育厅青年基金项目(11551067)
苍晶(1963-),女,教授,博士生导师,研究方向为植物生理生化与分子生物学。E-mail:cangjing2003@163.com
时间2014-1-9 22:49:15[URL]http://www.cnki.net/kcms/detail/23.1391.S.20140109.2249.014.html
苍晶,范博,宋扬,等.低温胁迫下冬小麦东农冬麦1号分蘖节相关EST-SSR信息分析[J].东北农业大学学报,2014,45(1):1-5. Cang Jing,Fan Bo,Song Yang,et al.Analysis of EST-SSR information in tillering nodes of winter wheat Dongnongdongmai1 under low temperature stress[J].Journal of Northeast Agricultural University,2014,45(1):1-5.(in Chinese with English abstract)
