水稻Os03g19020基因的克隆及亚细胞定位分析
2014-01-14柏锡贾蓓吕德康朱延明才华纪巍
柏锡,贾蓓,吕德康,朱延明,才华,纪巍
(东北农业大学生命科学学院,哈尔滨 150030)
水稻Os03g19020基因的克隆及亚细胞定位分析
柏锡,贾蓓,吕德康,朱延明,才华,纪巍
(东北农业大学生命科学学院,哈尔滨 150030)
Os03g19020基因是从水稻苗期冷害基因表达谱芯片中筛选到一个冷胁迫诱导表达的基因,Real-time PCR验证。结果表明,Os03g19020基因受冷诱导表达,响应迅速;氨基酸序列分析发现Os03g19020编码PHD-finger家族蛋白,具有典型的C4HC3保守结构域;同源性分析结果显示,03g19020与籼稻品种OsI-11202蛋白相似性达98%。将03g19020基因融合GFP基因,转化烟草原生质体细胞,结果表明,03g19020蛋白主要定位在细胞核内。
水稻;Os03g19020基因;PHD-finger;亚细胞定位
PHD-finger结构域是14种已知锌指结构域(Zinc-bingding motif)中的一种,是真核生物中一种进化保守的锌指结构,具有典型的C4HC3(cys4-His-cys3)保守结构域[1]。该结构域长约50~80个氨基酸,其半胱氨酸间距相对保守[2],在最后两个半胱氨酸之前存在一个色氨酸或其他芳香族氨基酸残基[3]。目前,已在多种植物中发现PHD-finger类基因,它们主要参与植物发育及生长过程调控[4-7]。此外,PHD-finger类基因还参与非生物胁迫调控。刘雨等通过过量表达OsPHD1基因证明该基因可以显著提高水稻对低温、高盐和干旱胁迫的耐受性[8]。水稻Os03g19020基因(PHD-finger family protein)编码的蛋白质属于植物同源结构域(Plant homeodomain,PHD结构域)家族,前期研究中,从水稻苗期冷害基因表达谱中发现Os03g19020基因响应低温胁迫。因此,本研究将对Os03g19020基因克隆并进行氨基酸序列分析,并对其蛋白进行亚细胞定位分析,为下一步分析提供参考。
1 材料与方法
1.1 材料
水稻品种采用粳稻品种空育131(Oryza sativaKongyu131),大肠杆菌(Escherichia coli)菌种为DH5α。克隆载体pGEM-T购自Promega公司;亚细胞定位载体为pBSK-eGFP。
1.2 总RNA提取与cDNA合成
水稻种子在42℃下处理3~5 d打破休眠。5% NaClO灭菌30 min,蒸馏水清洗后浸种1 d。置于湿滤纸上,暗处30℃催芽1 d。萌动后转移到Yoshida溶液中,28℃14 h光照(光照强度250 μmol·m-2·s-1),25℃10 h黑暗培养至三叶期。将幼苗置于4℃条件下处理,处理时间为0、1、3、6、12、24 h,取地上部组织。采用Trizol试剂(Invitrogen)提取水稻的总RNA,DNA酶Ⅰ处理后备用。以Oligod(T)18为反转录引物,用Super-Script III反转录酶(Invitrogen)试剂盒合成cDNA第一链。
1.3 OsGAMYB基因的冷胁迫条件下的表达分析
根据ABI PRISM 7500 Sequence Detection System对实时荧光定量PCR体系的要求设计引物。以水稻cDNA为模板,进行real-time PCR反应,筛选出能够获得锐利单峰溶解曲线的引物。用筛选出的引物对各样品进行Real-time PCR反应,每个样品设置3次技术重复。引物序列为:RT-Os03g190 20-sp:5'CAGCTCTAAGGTGGCCAGTC 3';RTOs03g19020-asp 5'ACTGGTGTCGGAGGATCAAC 3'。以Ef1-α基因为对照,引物序列为:Ef1-α sp 5'AGCCTCCTCCTCTCGCCAT 3';Ef1-α sp:5'TGT TCATCTCAGCGGCTTC 3'。实时定量PCR体系共25 μL。反应程序为:50℃预热2 min,95℃预变性2 min;95℃变性15 s,60℃退火30 s,40个循环,3次重复,采用2-ΔΔCt法进行定量分析。
1.4 Os03g19020基因的氨基酸序列分析与同源性分析
从NCBI下载Os03g19020的蛋白同源序列,运用Clustalx1.83及MEG 4.0软件进行氨基酸序列进行序列比对和进化树分析。
1.5 Os03g19020基因亚细胞定位载体的构建
根据Os03g19020基因cds区设计引物sense:5' CCATCGATATGGGGAAGGGAGGGGAAGG 3';antisense:5'CGGGATCCGACACCTTCAGTTCCTTGGAA GTTG 3',引物设计时应保证Os03g19020基因与GFP融合后不发生移码突变。PCR反应体系为:10 μL 10×KOD plus Buffer(Mg2+free),4 μL MgCl2(25 mmol·L-1),10 μL dNTP mix(2.0 mmol·L-1each),1.5 μL 5'PCR Primer(10 μmol·L-1),1.5 μL 3'PCR Primer(10 μmol·L-1),0.4 μg水稻cDNA,2 μL KOD plus Polymerase,ddH2O补足体积(总体积100 μL)。PCR反应条件为:94℃预变性5 min,94℃变性15 s,57℃退火30 s,68℃延伸90 s,共35个循环。将扩增出的PCR产物构建与表达载体pBSK-eGFP连接,转化DH5α,利用Qiagen max plastmid extraction kit(Qiagen)制备无内毒素质粒,调整质粒浓度至10 μg·μL-1。
1.6 原生质体制备及Os03g19020基因的亚细胞定位分析
取幼嫩烟草植物叶片,切成约0.5 mm的细丝,用预质壁分离液(组成为0.4 mol·L-1甘露醇和10 mmol·L-1氯化钙)分离1 h,加入10 mL酶液(组成为1%的纤维素酶R10、0.25%的离析酶MR10、6.8 mmol·L-1氯化钙、0.4 mol·L-1甘露醇,pH 5.5)于25℃对材料进行静置酶解2~3 h。用过滤除网去未完全消化的残渣,200 g离心3 min,弃上清,加入5 mL漂洗液W5(154 mmol·L-1NaCl、125 mmol·L-1CaCl2、5 mmol·L-1KCl、2 mmol·L-1MES),重复3次,用1 mL MMG溶液(0.4 mol·L-1甘露醇、15 mmol·L-1MgCl2、2 mmol·L-1MES)重悬。具体方法参见文献[9]。取20 μL原生质体溶液滴加于血球计数板上计数原生质体数,并换算成每克鲜组织所分离的原生质体数,最终调整浓度为105个·mL-1。PEG法转化原生质体,将100 μL原生质体溶液、10 μL质粒与110 μL 50%PEG溶液混匀后静置30 min,W5溶液清洗3次,25℃对材料进行静置恢复,16 h后观察。激光共聚焦显微镜下观察GFP的表达情况,GFP的激发波长为488 nm。
2 结果与分析
2.1 冷胁迫下Os03g19020基因的转录水平分析
粳稻品种空育131幼苗经4℃冷处理0、1、3、6、12和24 h后,总RNA提取结果见图1,Real-time PCR分析Os03g19020在冷胁迫条件下表达结果见图2。

图1 总RNA变性凝胶电泳检测Fig.1 Total RNA examined on formaldehyde agarose gel

图2 Real-time PCR分析Os03g19020在冷胁迫条件下表达Fig.2 Expression of Os03g19020 under cold stress by Real-time PCR
由图1可见,从左至右依次为冷胁迫处理0、1、3、6、12、24 h的变性凝胶电泳检测结果,在28S和18S区域有明显条带,亮度比例接近2∶1,初步判断其完整性良好,可进行反转录试验,用于下步Real-time PCR分析。三叶期水稻经冷处理时,Os03g19020基因随着处理时间的增加,表达量逐渐增加,12 h达到最高峰,以后又有下降趋势(见图2),说明Os03g19020是冷胁迫早期应答基因。
2.2 Os03g19020的氨基酸序列分析与同源性分析
在NCBI网站上以Os03g19020为靶序列进行BlastP比对,下载比对结果中与Os03g19020相似度高的基因蛋白序列,用Clustal X进行蛋白的多重序列比对,并用MEG4构建进化树,结果如图3A所示。进化树结果显示,与Os03g19020相似的蛋白来自多个物种,其中与Os03g19020相似性最高的是籼稻品种OsI-11202蛋白。OsI-11202被推断为是一种PHD finger蛋白。另外,利用Clustal X进行氨基酸序列分析。结果由图3B所示,PHD finger家族蛋白都含有典型的C4HC3(cys4-His-cys3)保守结构域,并且最后两个半胱氨酸之前存在一个色氨酸。

图3 Os03g19020与其他PHD finger蛋白的系统进化树分析与氨基酸序列比对Fig.3 Alignment of OsMYB17 with other PHD finger proteins and phylogenetic tree analysis
2.3 Os03g19020基因的克隆与亚细胞定位载体的构建
为研究Os03g19020分子功能,根据Os03g19020序列设计引物,以水稻cDNA为模板,扩增并克隆到终止密码子突变的CDS序列,长2 397 bp。将Os03g19020基因CDS片段连接到pBSK-eGFP载体上,转化大肠杆菌DH5α,取PCR鉴定为阳性克隆(结果未显示)的质粒,用ClaⅠ和Bam HⅠ进行双酶切鉴定,酶切片段在2 000 bp(见图4)。经测序证明,插入没引起读码框变化,表明载体构建成功,Os03g19020 CDS片段已经连接到亚细胞定位载体pBSK-eGFP上。
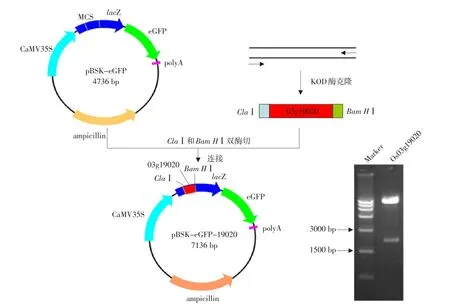
图4 Os03g19020亚细胞定位载体的构建Fig.4 Construction of protein sub-cellular location vectors for Os03g19020
2.4 Os03g19020蛋白的亚细胞定位分析
利用PEG法将构建的载体转染烟草原生质体,激光共聚焦显微镜下观察GFP的表达情况。在转Os03g19020+GFP基因的原生质体中,绿色荧光出现在细胞核中,而细胞质和其他部位没有看到GFP的表达(见图5);在只转GFP基因的对照中,绿色荧光在细胞核和细胞质中均有分布,说明Os03g19020蛋白是一种核定位蛋白。
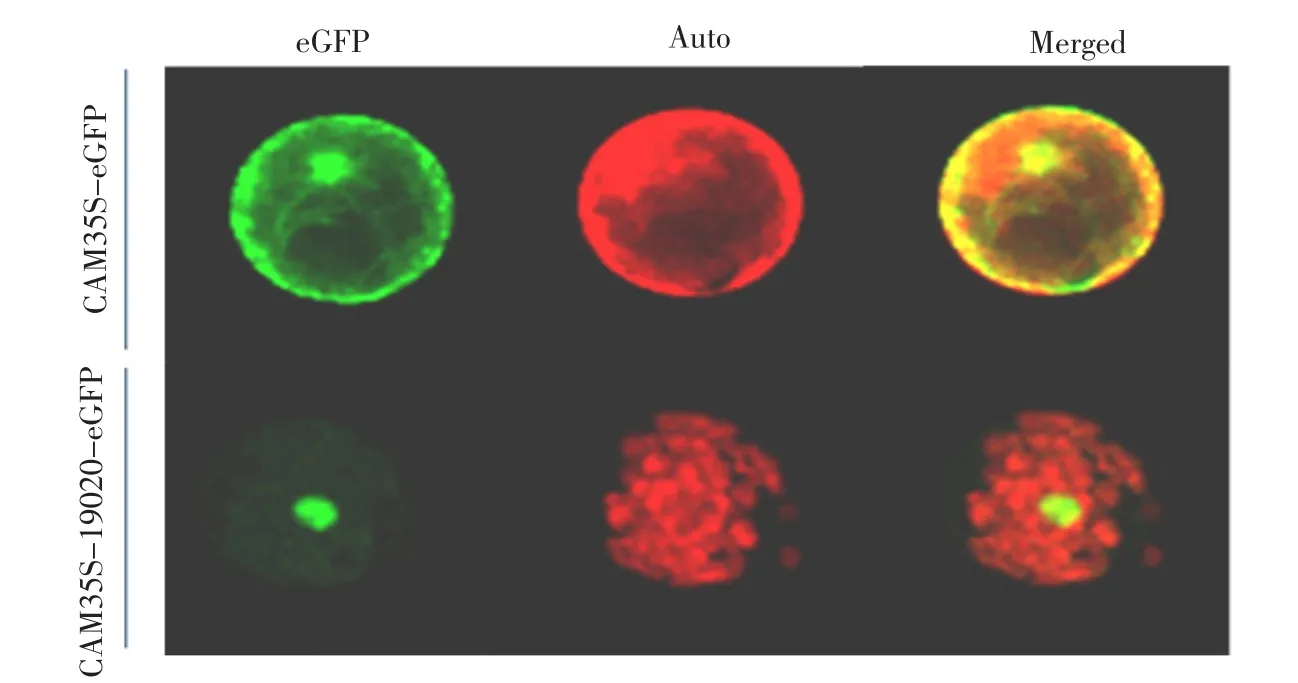
图5 Os03g19020的亚细胞定位Fig.5 Subcellular location of Os03g19020 in protoplast
3 讨论与结论
PHD-finger结构域是真核生物中一种进化保守的锌指结构域,通常参与蛋白质之间的相互作用,特别是对核小体组蛋白进行甲基化、乙酰化、磷酸化等修饰[10]。通过识别结合组蛋白甲基化密码,参与核内不同的生物学进程,如转录调控、细胞周期、凋亡等[11]。在植物中关于PHD-finger转录因子家族的功能研究主要集中在拟南芥上,Thomas等发现VIN3和VRN5对春化作用敏感,为春化作用所需[4];胡功铃等认为在春化处理过程中,VIN3及其同源基因编码的蛋白都具有PHD-finger结构域,该结构域通过对开花抑制基因FLOWERING LOCUS C染色质组蛋白进行H3K9、H3K27甲基化、H3K9和H3K14去乙酰化等修饰,调节FLC染色质结构状态,使其从松弛状态转变为高度凝缩状态而关闭其功能,影响FLC转录活性进而促进开花[12];Shinozaki等发现MS1蛋白是花粉成熟过程不可缺少的核信号分子[6]。
通过对水稻苗期胁迫处理发现Os03g19020基因是冷胁迫早期应答基因,推测该基因可能对苗期水稻耐性的提高具有一定作用。通过氨基酸序列分析,Os03g19020具有典型的C4HC3结构域,通过同源性分析,该蛋白与一些已有的PHD-finger蛋白具有很高同源性;亚细胞定位分析,Os03g19 020定位在细胞核内。预测该基因编码的蛋白可能与组蛋白结合,参与到核内的生物学过程,调控基因的转录,进而提高植物冷胁迫耐性,为下一步研究提供依据。
在基因的功能研究过程中,基因表达产物的功能与其在宿主细胞中的定位有重要关系,特别是对于真核细胞,蛋白质的亚细胞定位与其功能有着密切联系,蛋白质在生物体细胞核糖体内合成后,除分泌蛋白被分泌到细胞外,有些蛋白存在于细胞质中,其他蛋白都要按其功能精确地定位到相应的细胞器或细胞区域,如细胞核、细胞膜、内质网等[13]。蛋白质位于不同的细胞部位所行使的功能也不同,所以研究其在宿主细胞中的亚细胞定位,对研究外源基因表达产物的功能或未知新基因的功能具有很重要意义。目前被广泛应用的能进行亚细胞定位的报告基因主要是绿色荧光蛋白(Green fluorescent protein,GFP)。绿色荧光蛋白是细胞生物学和分子生物学研究中的一个重要工具,绿色荧光蛋白相对分子质量很小,能与多种不同的蛋白质N端或C端融合而保持其天然蛋白的特性,因此是一种直观性很强的遗传标记物[13-14]。外源基因通过与某单一序列的融合,可特异地进行细胞核、线粒体、质体、内质网等细胞器定位。
[1]Schindler U,Beckmann H,Cashmore A R.HAT3,a novel Arabidopsis homeodomain protein containing a conservedcysteinerich region[J].Plant J,1993,4(1):137-150.
[2]Kalkhoven E,Teunissen H,Houweling A,et al.The PHD type zinc finger is anintegral part of the CBP acetyltransferase domain [J].Mol Cell Biol,2002,22(7):1961-1970.
[3]Korfhage U,Trezzini G,Meier I,et al.Plant homedomain protein involved in t ranscriptional regulation of a pathogen defenserelated gene[J].Plant Cell,1994,6:695-708.
[4]Thomas G,Mylne J S,Pedro C,et al.The PHD Finger Protein VRN5 functions in the epigenetic silencing of Arabidopsis FLC[J]. Current Biology,2007,17:73-78.
[5]Ito T,Shinozaki K,The male sterility1 gene of Arabidopsis, encoding a nuclear protein with a PHD-finger motif,is expressed in tapetal cells and is required for pollen maturation[J].Plant Cell Physiology,2002,43(11):1285-1292.
[6]冯英,刘庆坡,薛庆中.水稻与拟南芥PHD-finger蛋白的系统分析[J].遗传学报,2004,31(11):1284-1293.
[7]Yukihiro I,Atsushi C,Mitsugu E,et al.Radial axis differentiation in a globular embryo is marked by HAZ1,a PHD-finger homeobox gene of rice[J].Gene,2004,331:9-15.
[8]刘雨,刘春霞,李早霞,等.植物同源结构域(PHD)-finger家族转录因子OsPHD1的过量表达可提高水稻的耐逆性能[J].农业生物技术学报,2011,19(3):462-469.
[9]Yoo S D,Cho Y H,Sheen J.Arabidopsis mesophyll protoplasts:A versatile cell system for transient gene expression analysis[J]. Nature Protocols,2007,2(7):1565-1572.
[10]Mariann Bienz The PHD finger,a nuclear protein-interaction domain[J].Trends Biochem Sci,2006,31(1):35-40.
[11]马红辉,方存磊,曾平耀.植物同源结构域(PHD结构域)——组蛋白密码的解读器[J].生物化学与生物物理进展,2008,35 (6):625-630.
[12]胡功铃,陈国平,胡宗利,等.植物同源结构域指蛋白在拟南芥等十字花科植物春化作用途径中的功能[J].生物工程学报2010,1(25-26):1-8.
[13]周善跃.三个烟草品种瞬时表达OsWAK1::GFP的差异[J].中国农学通报,2009,25(8):58-61.
[14]朱丹,王希,朱延明,等.植物亚细胞定位载体卡盒pCEG的构建及验证[J].东北农业大学学报,2011,42(4):83-87.
Cloning and subcellular localization analysis ofOs03g19020 gene in rice
BAI Xi,JIA Bei,LV Dekang,ZHU Yanming,CAI Hua,JI Wei
(School of Life Sciences, Northeast Agricultural University,Harbin 150030,China)
Os03g19020 was identified as a cold-responsive gene based on genome profiles and Real-time PCR analysis in response to cold stress.The Os03g19020 mRNA showed rapid up-regulation following cold-stress treatment.The amino acid sequence analysis revealed that Os03g19020 encoding PHD-finger family proteins,with a typical C4HC3 conserved domain;homology analysis showed that 03g19020 was similar to indica rice OsI-11202 protein,and the serial similarity was about 98%.The subcellular localization mainly in the nucleus suggested that Os03g19020 was an transcription factor.
rice;Os03g19020 gene;PHD-finger;subcellular localization
S572
A
1005-9369(2014)01-0029-05
2012-02-28
黑龙江省高校科技创新团队建设计划(2011TD005);东北农业大学博士启动基金(2009RC06)
柏锡(1975-),男,副教授,博士,硕士生导师,研究方向为植物抗逆分子生物学。E-mail:baixi@neau.edu.cn
时间2014-1-9 20:20:34[URL]http://www.cnki.net/kcms/detail/23.1391.S.20140109.2020.007.html
柏锡,贾蓓,吕德康,等.水稻Os03g19020基因的克隆及亚细胞定位分析[J].东北农业大学学报,2014,45(1):29-33.
Bai Xi,Jie Bei,Lv Dekang,et al.Cloning and subcellular localization analysis ofOs03g19020 gene in rice[J].Journal of Northeast Agricultural University,2014,45(1):29-33.(in Chinese with English abstract)
