不同育苗基质的团花根际微生物群落功能多样性特征
2014-01-04聂艳丽刘金凤
聂艳丽 ,陆 斌 ,刘金凤 ,童 清
(1.云南省林业技术推广总站,云南 昆明 650224;2. 普洱市林业科学研究所,云南 普洱 665000)
不同育苗基质的团花根际微生物群落功能多样性特征
聂艳丽1,陆 斌1,刘金凤1,童 清2
(1.云南省林业技术推广总站,云南 昆明 650224;2. 普洱市林业科学研究所,云南 普洱 665000)
利用Biolog-ECO技术对不同育苗基质培育下的团花根际微生物群落功能多样性进行研究。结果表明:不同育苗基质的团花根际微生物群落功能多样性有显著差异,13#甘蔗渣堆肥用作团花育苗基质能显著增强微生物对碳源的利用,13#育苗基质培育的团花根际微生物总体表现较高的活性,并且根际微生物利用碳源的类型以糖类、羧酸类和氨基酸类化合物相对较多,如:D-半乳糖酸γ--内脂、D-甘露醇、D-纤维二糖、衣康酸、L-精氨酸、L-天门冬酰胺和L-丝氨酸。
团花;根际微生物;微生物多样性;碳源利用
团花Anthocephalus chinensis,又称大叶黄梁木、毛老鹰树、埋格冬(傣语)、咪更昔(壮语),茜草科团花属,是热带和南亚热带的珍贵速生阔叶树种。对其育苗技术的研究早在1977年就已开始,但目前团花仍采用传统的土壤袋装育苗,苗期较难管理,根腐病等土传病害严重。随着天然林保护工程、退耕还林工程以及云南省林业建设六大工程的启动和实施,对热区珍贵阔叶树种的育苗技术,提出了越来越高的要求。
Biolog系统是Garland和Miss于1991年建立的一套用于研究土壤微生物群落结构和功能多样性的方法,通过对所得数据进行统计分析可以获得土壤微生物群落方面的信息[1]。该方法在国际上已成为研究土壤微生物群落流行的常规方法[2],已成功用于比较不同土壤类型[1],不同管理策略下的农业土壤[3],以及不同植被根际的土壤微生物群落代谢多样性。 Biolog-ECO微平板技术是测定土壤微生物对不同碳源利用能力和其代谢差异,表征土壤微生物功能多样性或结构多样性[4],反映土壤微生物群落结构[5]。该方法在以树木为研究对象的应用研究中几乎没有,团花根际微生物功能多样性的研究还未见报道。本研究利用Biolog-ECO技术研究不同育苗基质培育下的团花根际微生物群落功能多样性差异,及其对碳源的利用规律,对深入研究团花基质育苗有理论价值,特别是在不同育苗基质对团花土传病害的抑制机理研究奠定基础。在实施天然林保护、生态治理和西部大开发中,有计划地培育出优质苗木,提高人工林的质量,对保护天然林,解决用材也具有特殊意义。
1 材料与方法
1.1 供试材料
供试植物为云南热区珍贵速生阔叶树种团花Anthocephalus chinensis。供试的育苗基质是以甘蔗渣为主要原料进行堆制而成,采用堆肥化处理后的育苗基质13#、6#和1#,其理化性质见表1。供试的对照为森林表土,土壤的理化性状为:全碳含量 9.6 g·kg-1,全氮含量 0.2 g·kg-1,碳氮比(C/N)为39.9,铵态氮含量34.7 mg·kg-1,硝态氮含量 33.0 mg·kg-1,速效磷含量 24.8 mg·kg-1,速效钾含量 38.6 mg·kg-1,pH 值 7.4,EC 值 209 µs·cm-1。
1.2 团花生长条件
采用盆栽试验于2009年佛罗里达大学(北纬 25°51.2′,东经 -80°49.9′)进行。设 4 个处理,
表1 供试育苗基质的物理化学性质†

† 该表的数据来源于先前的研究成果[6]。
Table 1 Physical and chemical characteristics of different substrates每个处理5个重复。4个处理的编号为13#、6#、1#,和以不加堆肥的森林表土作为对照CK。盆的口径为12.5 cm,高为10 cm,放置在玻璃温室内。将团花种子消毒后,每盆均匀播撒60颗种子在各盆中,并在生长期间使其持水量保持在70%左右。待出苗后14 d定苗至30株,并统计出苗率。
1.3 样品采集
待幼苗长到2~3叶龄时,将团花根系从土或基质中取出,能够抖落掉的土称为土体土,附着在团花根系上,不能抖落的土为根际土,用小刷子轻轻扫掉,收集,置于-20℃冰箱保存,待测。
1.4 Biolog-ECO分析
微生物群落结构多样性采用微生物生态板Biolog-ECO方法分析,微平板购自美国BIOLOG公司,按化学基团的性质将ECO板上的31种碳源划分为6类,即糖类、氨基酸类、羧酸类、聚合物类、胺类和酚类化合物。每块板3个重复。
1.4.1 生态板接种液制备
称取团花根际附着基质5.0 g,放入三角瓶中,加入45 mL无菌0.1 mol·L-1磷酸缓冲溶液(K2HPO4/KH2PO4,pH值7.0),封口后在摇床上260 r·min-1震荡 30 min;取上清液,2 500 r·min-1离心 5 min;取上清液,12 000 r·min-1离心20 min;弃上清液,加5 mL 0.85%的无菌NaCl于离心管中溶解沉淀;0.85%NaCl稀释接种液使其浊度为99%[7]。
1.4.2 上板ELISA反应及数据记录
用移液器将接种液分别加在Biolog ECO平板的各孔中,每孔150 μL,每个样32孔,每块板3个重复,置于25℃恒温培养;培养过程中分别在4、24、48、72、96、120、144和168 h记录590 nm波长的数据。
1.4.3 每孔颜色平均变化率的分析
Biolog ECO 微平板ELISA反应一般采用每孔颜色平均变化率AWCD(Average well color development)来描述,计算不同时间的平均颜色变化率。计算公式为:
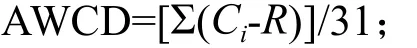
其中,Ci是除对照孔外各孔吸光度值,R是对照孔吸光度值。
采用SAS GLM Duncan多重比较进行多样性指数差异显著分析(P<0.05)。
1.4.4 群落参数的统计分析
分析确定Biolog微平板培养最佳统计时间[8],目前大部分学者应用培养72 h的数据进行土壤微生物群落功能多样性的研究[1-2,7,9],因为大部分微生物在72 h处于对数生长期。本研究不同处理的育苗基质Biolog微生态板培养时,72 h微生物刚刚进入旺盛生长期,在96 h碳源利用度快,效果

Shannon-Wiener 碳源均匀度指数(E):评价物种的均匀度,由H得出,其计算公式为:最明显,在120 h 碳源利用就进入衰退期了。故本研究采用培养96 h的数据,将碳源分为6类,分析团花根际微生物群落对31种碳源的利用情况,即根际微生物群落代谢功能多样性。
以该统计时间的数据计算多样性指数(H)、碳源均匀度指数(E)和微生物丰富度指数(S)。用这3个指数来表示根际微生物群落代谢功能多样性[2]。
Shannon-Wiener 多样性指数(H):估计物种的丰富度,其计算公式为:
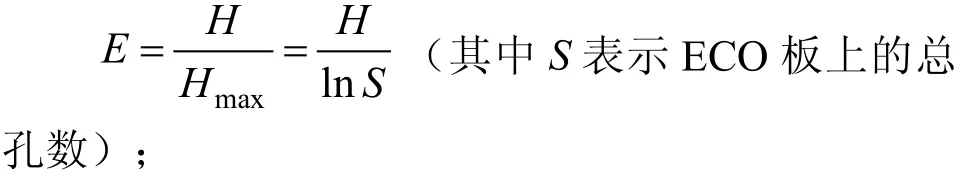
微生物丰富度指数(S):微生物群落利用碳源种类的数目,即颜色变化孔数。
BIOLOG测定及数据处理按相关分析方法进行[1,10-11],采用SAS中PCA进行主成分分析(SAS Institute Inc., Cary, NC),统计软件应用SAS for Windows V6.12。
2 结果与分析
2.1 不同育苗基质对根际微生物碳源利用的动力学特征
图1为31种碳源的AWCD值,不同育苗基质的根际微生物活性均随培养时间的延长,微生物利用碳源量呈逐渐增加的趋势。13#根际利用碳源总量在整个培养过程都高于其它处理,说明13#根际微生物利用碳源能力强,总体表现较高的活性。培养144 h时,对照CK的根际微生物AWCD值显著低于其它3个处理,13#基质的根际微生物群落AWCD值均高于其它处理,并且差异显著,13#处理根际微生物的AWCD值分别高于CK、1#、和6#处理的50.0%,21.2%和8.5%,13#、6#和1#处理根际微生物的AWCD值分别高于CK的50.0%、38.2%和23.8%。
2.2 不同育苗基质对根际微生物群落多样性的影响
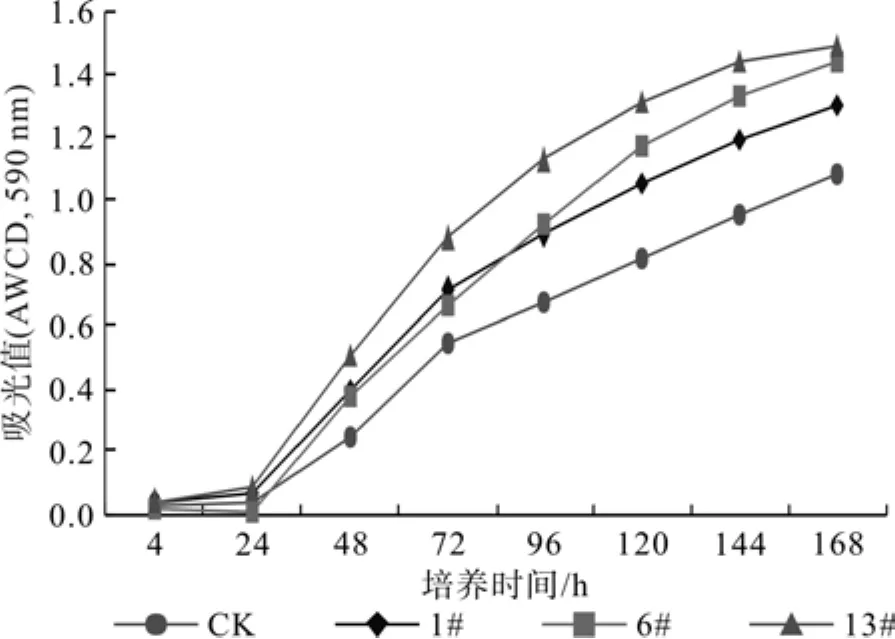
图1 不同育苗基质对团花根际微生物群落的每孔颜色平均变化率Fig.1 AWCD of rhizosphere microbial community of different substrates
不同育苗基质的根际微生物群落代谢多样性指数、均匀度指数和丰富度指数见表2。由表2可以看出13#基质的Shannon多样性指数最高,并与6#基质和CK差异显著,13#基质的微生物多样性指数是CK的1.01倍,6#基质的微生物多样性指数最低,1#基质的微生物多样性指数较6#基质有增加趋势,但差异不显著。从不同处理的Shannon均匀度结果来看,1#基质的微生物均匀度指数最高,显著高于6#和13#基质。从不同育苗基质的丰富度指数结果来看,13#基质微生物的丰富度显著高于1#。13#基质微生物多样性高更可能的原因是综合作用的结果,与营养、环境、微生物种群代谢速度等关系密切。
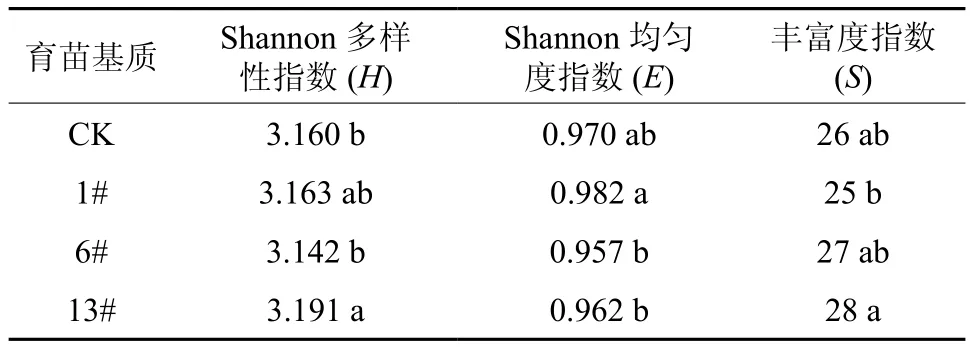
表2 不同育苗基质对根际微生物群落多样性的影响†Table 2 Effects of different substrates on microbial biodiversity indexes
2.3 不同育苗基质的根际微生物对不同类型碳源利用的差异
团花根际微生物多样性反映了微生物群落总体的变化,但不能反映微生物群落代谢的具体信息。在本研究中通过对4种不同育苗基质的团花根际微生物群落代谢多样性类型的比较,分析不同育苗基质的团花根际微生物对这6类碳源利用率上的差异,从而解析不同育苗基质对微生物群落的影响。
表3表示不同育苗基质培育下的团花根际微生物群落对Biolog ECO微平板中6类碳源利用差异。由表3可知,13#和6#与CK相比根际微生物碳源利用情况发生改变最为显著,13#与CK相比对聚合物类、糖类、羧酸类、氨基酸类、胺类和酚类化合物的利用率分别升高了19.2%、50.1%、63.0%、156.8%、137.8% 和 190.7%;6#与CK相比对聚合物类、糖类、羧酸类、氨基酸类、胺类和酚类化合物这6类碳源的利用率分别升高了0.7%、26.4%、26.5%、107.8%、141.9%和99.1%。13#基质对各种碳源的利用均显著增强,对团花苗木起到更大的调节作用。
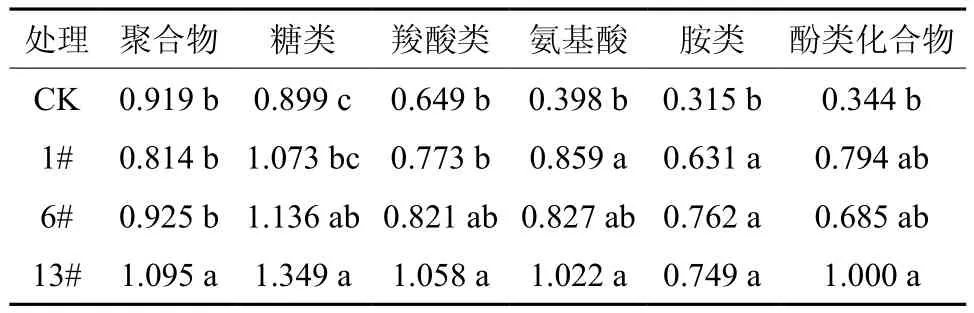
表3 不同育苗基质的根际微生物对6类碳源的利用†Table 3 Effects of different substrates on carbon source utilization by rhizosphere microbial community
2.4 不同育苗基质的根际微生物群落代谢功能主成分分析
通过主成分分析可知不同育苗基质对团花根际微生物利用碳源能力的影响。图2为不同育苗基质的根际微生物群落代谢功能主成分分析,由图2可知,不同育苗基质在主成分1(PC1)轴和主成分2(PC2)轴上都分异明显,尤其是13#育苗基质与其它处理差异显著。13#与其它育苗基质分异显著,说明13#育苗基质根际微生物的碳源利用能力比其它处理增强,达显著水平;13#育苗基质根际微生物的碳源利用能力比6#增强,但不显著。

图2 不同育苗基质对根际微生物群落代谢功能主成分分析Fig. 2 Principal component analysis of rhizosphere microbial community of different substrates
对PC1和PC2的贡献率在(±0.2~0.4)的碳源能反映微生物对主要碳源的利用信息[12]。表
4为不同育苗基质对团花根际微生物群落代谢功能主成分分析,由表4可知:对PC1影响大的是主要是糖类、羧酸类和氨基酸类,对PC2影响大的主要是糖类。在主成分1中,聚合物中的α-环式糊精,糖类中的D-半乳糖酸γ-内脂、D-甘露醇、D-纤维二糖,羧酸类中的D-半乳糖醛酸、衣康酸、α-丁酮酸、D-苹果酸,氨基酸中的L-精氨酸、L-天门冬酰胺及L-丝氨酸,胺类中的苯乙胺,酚类化合物中的4-羟基苯甲酸,各类成分贡献率占20%以上;在主成分2中,聚合物中的吐温80,糖类中的β-甲基-D-葡萄糖苷、α-D-乳糖、1-磷酸葡萄糖、D,L-α-磷酸甘油,羧酸类中的丙酮酸甲酯,氨基酸中的L-苯丙氨酸、L-苏氨酸、甘氨酸-L-谷氨酸,各类成分贡献率占23%~35%。

表4 PC1和PC2贡献率较高的部分碳源†Table 4 Carbon sources of higher contribution rate to PC1 and PC2
3 讨 论
根际微生物的变化受多种因素的影响,如育苗基质理化性、育苗基质中的其它生物因素及育苗管理措施等。在本研究中采用4种不同育苗基质,包括森林表土对照CK,育苗基质是主要的影响因子,13#处理使得根际微生物的活性和多样性显著增强(表2)。但究竟是因为13#育苗基质改善了微生物的生存环境,还是丰富了微生物所需的营养物质,还有待于进一步的研究。
每孔颜色平均变化率(AWCD)反应根际微生物活性[1],微生物对碳源利用的AWCD值主要反映微生物群落结构和活性的差异[13-20]。从理论上分析,根际微生物个体数量多且种群丰富,AWCD值可达到较大值,若根际微生物个体数量少而种群丰富,则开始AWCD值较小,但随着培养时间延长,微盘中丰富的碳源使微生物不断繁殖,所以AWCD值逐渐增加;若种群丰富度差(即种类少),而某些种类的微生物数量多,则培养开始AWCD值增加较快,但较早达到最大恒定值,因为当能被利用的碳源消耗尽后,AWCD值不再增加。由此易知,根际不同的微生物群落结构会产生不同的碳源利用模式。
AWCD和微生物群落多样性指数、均匀度指数和丰富度指数反映的是微生物活性和功能多样性[14]。本研究应用Biolog ECO微孔板中有31种培养基,对不同处理的育苗基质或森林表土进行微孔颜色平均变化率测定,结果显示:13#处理根际微生物的AWCD值分别高于CK、1#、和6#处理的50.0%,21.2%和8.5%,说明13#基质的根际微生物群落AWCD值均高于其它处理,并且差异显著,并表明作物根际微生物代谢多样性与基质环境密切相关。13#育苗基质的微生物活性、群落多样性指数、均匀度指数和丰富度指数均显著高于对照CK,团花苗木的根际微生物以糖类代谢群为优势类群,13#育苗基质的根际微生物群落结构相对稳定,根际微生物群落对糖类、羧酸类和氨基酸类碳源的利用率显著高于聚合物类、胺/氨类和其它类碳源。应用培养法研究土壤微生物群落时,糖类、羧酸类和氨基酸类碳源可优先考虑。
不同育苗基质对团花根际微生物群落多样性产生显著影响,研究表明糖类、羧酸类和氨基酸类是区分土壤微生物群落功能多样性的主要碳源类型,处理间的差异主要是根际微生物群落对糖类、羧酸类和氨基酸类碳源利用的差异引起的,这3类碳源是区分不同根际微生物群落的最敏感碳源。不同育苗基质对团花根际微生物的数量、活性、各种多样性指标、碳源代谢能力等的影响都有显著差异,进而对苗木的生长发育产生了较大影响。但具体是哪些微生物群落发生了变化,是一类主导或是几类综合影响,目前还不清楚。因此,应进一步研究发生显著变化的微生物类群,才能更好的阐明根际微生物群落变化与不同机制类型相互关系。
[1] Garland J L, Milis A L. Classification and characterization of heterotrophic microbial communities of on the basis of patterns of community- level sole- carbon- source utilization [J]. Appli.Environ. Microbiol., 1991, 57: 2351-2359.
[2] Schutter M, Dick R. Shifts in substrate utilization potential and structure of soil microbial communities in response to carbon substrates[J]. Soil Biology and Biochemistry, 2001,33(11): 1481-1491.
[3] Buyer J S, Drinkwater L E. Comparison of substrate utilization assay and fatty acid analysis of soil microbial communities [J].Journal of Microbiological Methods, 1997, 30(1): 3-11.
[4] 徐华勤, 肖润林, 邹冬生,等. 长期施肥对茶园土壤微生物群落功能多样性的影响[J]. 生态学报, 2007, 27(8): 3355-3361.
[5] Garland J L. Analytical approaches to characterization of samples of microbial communities using sole C source utilization patterns [J].Soil Biology & Biochemistry, 1996, 28: 213-221.
[6] 聂艳丽, 周跃华, 李 娅, 等. 甘蔗渣堆肥化处理及用作团花育苗基质的研究[J].农业环境科学学报, 2009, 28(2):380-387
[7] 范成明. 土壤生物消毒对大白菜根肿病的防效评估及防治机理研究[D].昆明:云南农业大学, 2007.
[8] Grove J A, Kautola H, Javadpour S, et al. Assessment of changes in the microorganism community in a biof i lter [J]. Biochemical Engineering Journal, 2004, 18(2):111-114.
[9] Bending G D, Turner M K, Jones J E. Interactions between crop residue and soil organic matter quality and the functional diversity of soil microbial communities [J]. Soil Biology and Biochemistry, 2002, 34(8): 1073-1082.
[10] 孔维栋,刘可星,廖宗文,等.不同腐熟程度有机物料对土壤微生物群落功能多样性的影响[J].生态学报,2005,(9):2291-2296.
[11] 白学慧, 姬广海, 李成云,等. 魔芋与玉米间栽对魔芋根际微生物群落代谢功能多样性的影响[J].云南农业大学学报,2008, (6): 736-740.
[12] 蔡燕飞, 廖宗文, 董 春,等. 番茄青枯病的土壤微生态防治研究[J].农业环境保护, 2002, (5): 417-420.
[13] 陈云兰, 姬广海, 董 坤. 抗生素溶杆菌13-1对魔芋土壤微生物群落的影响[J].云南农业大学学报:自然科学版, 2010,(4): 487-493.
[14] Liu A M, Huang W Y, Microbial activities and functional diversity of community in soils polluted with copper tmling after cultivate [J]. Ecology and Environment, 2005, 14(6): 876-879.
[15] 李 蓉,周德明,吴 毅,等.杉木根际溶磷菌筛选及其部分特性的初步研究[J].中南林业科技大学学报,2012,32(4):95-99.
[16] 牛芳华,李志辉,周德明,等.尾巨桉幼苗根际土壤微生物分布特点及酶活性[J].中南林业科技大学学报,2011,31(3):151-155.
[17] 尹晓宁,马 明,张 坤,等.不同覆盖条件对陇东旱塬苹果园土壤水分及果实品质的影响[J].经济林研究,2012,30(1):34-39.
[18] 闫秀婧,王惠玲,吴 霞,等.天水市麦积区经济林种植土壤中 NPK 的空间变异分析[J].经济林研究,2012,30(1):62-66.
[19] 陈炜青,姜成英,吴文俊,等.不同插穗处理对油橄榄温床扦插生根率的影响[J].经济林研究,2012,30(4):60-63.
[20] 邓小梅,欧阳昆唏,张 倩,等.团花研究现状及发展思考[J].中南林业科技大学学报,2011,31(11):90-95.
Diversity characteristics of rhizospheric microorganism community function of Anthocephalus chinensis cultured with different substrates
NIE Yan-li1, LU Bin1, LIU Jin-feng1, TONG Qing2
(1. Forestry Department of Yunnan Province, Kunming 650224, Yunnan, China; 2. Puer Forestry Academy, Puer 665000, Yunnan, China)
The microbial community functional diversity of rhizospheric microorganism ofAnthocephalus chinensiscultured with different substrates were researched by applying the Biolog ECO method. The results show that the microbial community functional diversity in rhizosphere soil was signif i cant difference with different substrates; the microbes in rhizosphere utilized more carbons with 13# sugarcane bagasse as the substrate, which the microbe in rhizosphere had relatively higher activity of metabolism than that of others treatments. Furthermore, the types of carbon source utilized by microbial community in rhizosphere were carbohydrates, carboxylic acid,and amino acid such as D-Galactonic Acid γ-Lactone, D-Mannitol, D-Cellobiose, Itaconic acid, L-Arginine, L-Asparagine, and L-Serine.
Anthocephalus chinensis; rhizospheric microorganism; microbial diversity; carbon source utilization
S718.4
A
1673-923X(2014)01-0007-05
2013-09-20
云南省自然科学基金委资助项目(2004C0027Q);中央财政林业科技推广示范项目[2011]TQYN03;云南省技术创新人才培养项目(2010CI092)
聂艳丽(1974-),女,黑龙江人,副研究员,博士,主要从事林木基质育苗和林木组培的研究与推广工作;E-mail:nieyanli@163.com
童 清(1964-),男,云南景谷人,正高级工程师,主要从事森林培育研究;E-mail:pelkstq@163.com
[本文编校:吴 毅]
