青海湖北岸高寒草甸草原非生长季土壤呼吸对温度和湿度的响应
2014-01-02陈骥曹军骥魏永林刘吉宏马扶林陈迪超冯嘉裕夏瑶岑燕
陈骥,曹军骥,魏永林,刘吉宏,马扶林,陈迪超,冯嘉裕,夏瑶,岑燕
(1.中国科学院地球环境研究所 黄土与第四纪地质国家重点实验室,陕西 西安710075;2.中国科学院大学,北京100039;3.西安交通大学全球环境变化研究院,陕西 西安710075;4.青海省海北州海北牧业气象试验站,青海 海北810200;5.琼州学院理工学院,海南 三亚572022)
青藏高原雄居亚洲大陆的中部,总面积250万km2以上,有“地球第三极”之称。青海湖流域位于青藏高原东北部,是西风、东南季风和西南季风的交汇带,属于全球气候变化敏感区域和脆弱地区,是典型的高原半干旱高寒气候,具有常年干寒、多风、温差较大、太阳辐射强等特点。青海湖流域坐落于海拔3000m以上,地处高海拔高纬度,以其独特的地形成为全球变化的敏感区域,因而倍受世界各国政府和科学家的普遍关注[1]。作为青海湖流域较为典型的植被类型,高寒草甸化草原生态系统具有典型的地带性分布特征,其生态系统功能极其脆弱[2]。放牧与围栏封育是该地区草地利用的最主要方式之一,在退化土地恢复和植被重建过程中,围栏封育以其投资少、见效快的特点,已经成为退化土地恢复的重要措施之一,并被世界各国广泛采用。封育后,由于地上生物量和盖度的变化从而引起相应的土壤温度与湿度的变化[3],进而引起相应的土壤生物化学过程的变化[4]。
土壤呼吸是陆地生态系统碳循环的重要组成部分,大约把植被通过光合作用所固定的碳的1/4返还到大气中,是土壤、大气、植被中碳交换的重要过程,对陆地生态系统碳循环有巨大的作用,土壤CO2呼吸大约占到土壤呼吸的3/4,是地下碳循环的重要体现[5]。非生长季土壤呼吸是非生长季生态系统释放CO2的一个重要组成部分,显著地影响着碳收支[6-7]。相关研究表明陆地生态系统碳汇具有明显的季节变化和年际变化[8]。然而,已往的大多数研究都集中在对生长季土壤呼吸的测定,对年土壤呼吸量的估算大多基于非生长季土壤呼吸为0的假设[6]。近期的研究表明,非生长季土壤呼吸不仅不为0而且能占到年土壤呼吸总量的14%~30%,并且受多种因素影响,例如降雪厚度[7]、土壤温度和含水量[4]以及土壤微生物的活性[9]。土壤呼吸的温度敏感性在相当大的程度上决定着全球气候变暖背景下土壤与大气中CO2的交换能力。在以往的生态学模型中大多采用单一不变的Q10值进行模拟土壤与大气之间的CO2交换,并在此基础上估计全球温度的变化。但实际上,近些年的研究表明,土壤呼吸的温度敏感性是随着温度的变化而变化的,在不同的环境条件下土壤呼吸往往具有不同的温度敏感性。然而,一方面由于非生长季青藏高原恶劣的工作条件;另一方面由于以往的研究大多认为非生长季的土壤呼吸对全年土壤呼吸量的贡献不重要,甚至认为非生长季土壤呼吸为0,使得目前关于青藏高原地区非生长季土壤呼吸的研究和报道较少。使得我们对非生长季土壤呼吸对年土壤呼吸的贡献以及非生长季土壤呼吸对温湿度的响应的认识存在一定的局限。
因此,对非生长季土壤呼吸的测定和研究对于更加精确估算生态系统的碳循环和土壤碳固存能力及其对温室效应的贡献有重要意义;同时对非生长季土壤呼吸大小及其影响因素的研究有利于改善区域和全球的碳循环模型、对预测生态系统对全球变化的响应及其相互作用也具有重要意义。本文基于短期围栏封育的基础研究了非生长季土壤呼吸的变化,旨在:1)得出非生长季土壤呼吸占年土壤呼吸量的比例;2)讨论在非生长季各环境因子对土壤呼吸的贡献;3)讨论非生长季的土壤呼吸温度敏感性。
1 材料与方法
1.1 研究样地
研究样地位于中国气象局海北草地生态实验站(100°51′E,36°57′N,海拔3140m),隶属于青海省海北州海北牧业气象试验站,主要为冬季牧场。研究区地处欧亚大陆腹地,属典型的高原大陆性气候。年平均气温和年降水量分别为0.8℃和398.2mm,年日照时间为2912.7h,平均无霜期为48d,土壤为砂壤土。草地植被为典型的草甸化草原植物,主要有西北针茅(Stipasareptanavar.krylovii)、紫花针茅(S.purpurea)、洽草(Koeleria cristata)、冷地早熟禾(Poacrymophila)、矮生嵩草(Kobresiahumilis)、猪毛蒿(Artemisiascoparia)、紫菀(Astertataricus)和花苜蓿(Medicagoruthenica)等。实验站地形平坦,植被分布均匀(冠层高度为20~30cm),可基本代表环青海湖地区草甸草原的特征。研究样地非生长季时间为10月20日到次年4月20日。
1.2 实验设计
选取于2008年(GE5)和2010年(GE3)围栏封育的草地为研究对象,围栏分别于2007和2009年非生长季完成,以围栏外自由放牧地为对照样地(FG)。分别在自由放牧(FG),2010年围栏封育(GE3)和2008年围栏封育(GE5)的草地中选取地势平坦大约为200m×200m的样地作为研究样地,各研究样地间有5~10m的缓冲区。围封草地终年禁牧,栏外自由放牧,放牧强度大约为0.5只牦牛/hm2和2.5只羊/hm2。在围栏封育之前各样地均为自由放牧样地。在每个选取的作为实验对象的样地里,选取6个样点(1m×1m),彼此互为重复,各样地内的样点选取要考虑到空间异质性所引起的差异,所以互为重复的样点的选取尽量保持较高的相似性,为了更好地反映各样地的实际情况,同一样地内各样点间的距离至少为3m。
1.3 土壤呼吸测定方法
内径为20cm,高度为5cm的PVC圈子在实验设计之初就已经插入土壤中,插入深度为2~3cm。PVC圈子内活的生物体至少于土壤呼吸测定的前1d全剪掉。剪掉的草留在PVC圈子内自然分解,并尽可能地不扰动地表凋落物[9]。在年季变化上,生长季每月至少测定2次土壤呼吸值,非生长季每月至少测定1次。生长季内同时还要测定土壤呼吸的日变化,每2h测定1次,从早上8点测到晚上20点。土壤呼吸的测定采用West system的Li-8100,并且在晴朗的日子里完成。非生长季土壤呼吸测定的具体时间分别为2012年10月14日,11月13日和2013年2月14日,3月23日,4月20日。
1.4 土壤温、湿度测定方法
采用HOBO自动数据采集器(Onset Computer Company,Pocasset,MA,USA)记录10cm土壤温、湿度变化。数据器每min采集1次数据,每5min记录1次平均值。土壤温、湿度测定仪器尽量放置在样地周边,以免人为踩踏影响植物的正常生长[3]。最终数据处理为小时平均值和日平均值,用和土壤呼吸相匹配的瞬时土壤温、湿度值来计算他们的关系。
1.5 数据处理方法与作图
利用Excel 2003和SPSS 13.0完成所有统计分析,应用单因素方差分析(One-way ANOVA)和最小显著性差异(LSD)方法检验差异性,应用Origin 8.0作图。应用R=aebT模型拟合土壤呼吸的温度敏感性,再根据Q10=e10b来计算Q10。其中R为土壤呼吸速率,T为10cm土壤温度,a为0℃时土壤呼吸的截距,b为常数[10]。对于生长季的土壤呼吸采用R=aebT(W)分别对温度(T)和湿度(W)进行拟合,在非生长季采用R=aT(W)+b模型对土壤异养呼吸与温、湿度的关系进行拟合。应用相邻2次间的间隔天数和呼吸速率来计算年、季节呼吸量[11]。
2 结果与分析
2.1 研究样地微气象对围栏封育的响应

图1 全年、生长季和非生长季的降雨和蒸散发的变化Fig.1 The variation of precipitation and moisture evaporation(ME)over annual scale,growing season and dormant season
如图1所示,全年的降雨都基本发生在生长季,非生长季降水量为23.7mm,仅占全年降水量的4.9%,非生长季的蒸发量为503.5mm,占全年蒸发量的34.5%。全年、生长季和非生长季蒸发量分别是降水量的3.0,2.1和21.2倍,所以该研究区域有较为干旱环境,尤其是在非生长季。在非生长季,土壤具有较低的体积含水率,这和非生长季具有较少的降雨是相一致的。在整个非生长季内,围栏封育增加了土壤体积含水率但未达显著水平(图2C),自由放牧、3年围栏封育和5年围栏封育平均土壤体积含水率分别为2.5%,2.8%和3.4%。
在非生长季土壤温度的变化趋势和大气温度的变化趋势相一致,最低温均出现在1-2月之间(图2A,B)。大气最低温度为-27.8℃,自由放牧、3年围栏封育和5年围栏封育土壤最低温度分别为-12.2,-11.0和-10.4℃。方差分析结果表明,非生长季土壤温度的差异在自由放牧、3年围栏封育和5年围栏封育样地间未达到显著差异水平,但围栏封育降低了土壤温度。在整个非生长季内,自由放牧、3年围栏封育和5年围栏封育平均土壤温度分别为-0.1,-0.4和-0.9℃。
2.2 非生长季土壤呼吸对短期围栏封育的响应
非生长季土壤呼吸速率的变化表现出和大气温度、土壤温度、土壤湿度相一致的单峰变化规律(图2D)。非生长季土壤呼吸速率的变化范围为0.42~1.25,0.40~1.17,0.28~1.01μmol/(m2·s),峰值出现在1到2月之间,此时也具有较低的温度和湿度。显著性检验结果表明,非生长季2012年10月14日和2013年4月20日土壤呼吸在FG,GE3和GE5之间有显著性差异(P<0.05),但在FG和GE3之间无显著性差异。
如图3所示,自由放牧样地土壤呼吸平均速率在全年、生长季和非生长季分别为2.7,4.1和0.9μmol/(m2·s);3年围栏封育样地土壤呼吸平均速率在全年、生长季和非生长季分别为2.5,3.7和0.8μmol/(m2·s);5年围栏封育样地土壤呼吸平均速率在全年、生长季和非生长季分别为2.3,3.5和0.7μmol/(m2·s)。方差分析结果表明,自由放牧、封育3年和封育5年在全年、生长季和非生长季尺度上土壤呼吸速率均有显著性差异(P<0.05);在全年尺度上,土壤呼吸速率仅在GE5和FG,GE3间有显著差异,在生长季尺度上土壤呼吸在FG、GE3、GE5间均有显著性差异,在非生长季尺度上土壤呼吸无显著性差异(P<0.05)。短期围栏封育作用对非生长季土壤呼吸作用影响不明显,但略微降低了非生长季的平均土壤呼吸速率。自由放牧、3年围栏封育和5年围栏封育样地非生长季土壤呼吸平均速率分别是全年土壤呼吸平均速率的0.21,0.22和0.19倍,是生长季土壤呼吸平均速率的0.32,0.33和0.30倍。

图2 非生长季气温,10cm土壤温度,10cm土壤湿度,土壤呼吸速率对围栏封育的响应Fig.2 Response of air temperature(AT,A),10cm soil temperature(ST,B),10cm soil moisture(SM,C),and soil respiration rate(Rs,D)to grazing exclusion

图3 全年、生长季和非生长季平均土壤呼吸速率(Rs)对围栏封育的响应Fig.3 Response of average soil respiration rate(Rs)to grazing exclusion over annual scale,growing season,and dormant season
在较长的非生长季中,研究样地虽然具有较低的土壤呼吸速率,但因土壤呼吸所排放的碳量并不少。自由放牧、3年围栏封育和5年围栏封育样地非生长季因土壤呼吸所排放的碳量分别为117.2,109.2和100.7g C/m2,占全年因土壤呼吸所排放碳量的21.2%,22.3%和23.2%(图4)。方差分析结果表明,自由放牧、封育3年和封育5年在全年、生长季和非生长季尺度上土壤呼吸排碳量均有显著性差异(P<0.05);在全年尺度上,土壤呼吸排碳量仅在GE5和FG、GE3间有显著差异,在生长季尺度上土壤呼吸在FG、GE3、GE5间均有显著性差异,在非生长季尺度上土壤呼吸无显著性差异(P<0.05)。
2.3 非生长季土壤呼吸与土壤温、湿度的关系
如图5所示,在非生长季土壤呼吸具有较低的温度敏感性,自由放牧、3年围栏封育和5年围栏封育样地的土壤呼吸温度敏感性分别为1.5,1.8和1.7。在全年尺度上和生长季尺度上土壤呼吸都具有较高的温度敏感性。在非生长季土壤温度和土壤体积含水率对土壤呼吸的解释率都较低,土壤温度解释了自由放牧、3年围栏封育和5年围栏封育样地土壤呼吸变化的36.8%,53.0%和34.8%;土壤湿度解释了36.2%,51.7%和37.9%(图6)。

图4 全年、生长季和非生长季土壤呼吸(Rs)所排放的碳对围栏封育的响应Fig.4 The amount of carbon released by soil respiration(Rs)over annual scale,growing season,and dormant season
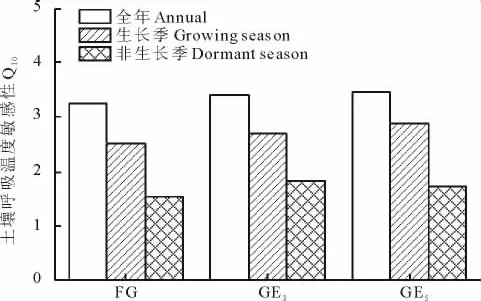
图5 全年、生长季和非生长季土壤呼吸温度敏感性(Q10)对围栏封育的响应Fig.5 The response of Q10to grazing exclusion over annual scale,growing season,and dormant season

图6 非生长季土壤呼吸与土壤温度和土壤湿度间的线性关系Fig.6 Linear relationship between soil respiration(Rs)and soil temperature(A,B,C),and soil moisture(D,E,F)during dormant season
同时为了区分土壤湿度在非生长季对土壤呼吸的重要性,将围栏封育导致的差异性忽略掉,选取一定的土壤湿度范围(土壤体积含水率小于1.5%、土壤体积含水率在1.5%~3.0%和土壤体积含水率在3.0%~5.0%),在此范围内讨论土壤温度与土壤呼吸的变化。如图7所示,当土壤体积含水率小于1.5%时,土壤呼吸与温度间的关系不明显(F=0.296,P>0.5,R2=0.022);当土壤体积含水率在1.5%~3.0%时,土壤呼吸随温度的增加而显著增加(F=80.138,P<0.001,R2=0.684);当土壤体积含水率在3.0%~5.0%时,土壤呼吸与温度具有更好的相关性(F=79.831,P<0.001,R2=0.807)。说明一定的土壤湿度是保证土壤呼吸过程进行的必要条件。
3 讨论
3.1 非生长季土壤呼吸的重要性

图7 不同范围土壤体积含水率下土壤呼吸随温度的变化Fig.7 Response of soil respiration to soil temperature over different range of soil volume water content
土壤呼吸是陆地生态系统碳收支中最大的通量之一,是国际生物圈地圈碳循环的重要组成部分,研究和分析土壤呼吸机制,精确估算因其引起的碳收支问题具有重要意义。放牧与围栏封育是我国青藏高原地区草地的主要利用方式之一,其强烈的影响着草地微气象环境和草地碳循环过程。由于非生长季的干旱环境,在以往的研究中大多集中于生长季和土壤呼吸作用,对非生长季的土壤呼吸作用大多认为0或者认为非生长季的土壤呼吸作用不重要。但近期的研究结果表明陆地生态系统碳汇具有明显的季节变化和年际变化[8],非生长季土壤呼吸是非生长季生态系统释放CO2的一个重要组成部分,显著地影响着碳收支[6]。本研究结果表明,非生长季土壤呼吸平均速率分别是全年土壤呼吸平均速率的0.21,0.22和0.19倍,是生长季土壤呼吸平均速率的0.32,0.33和0.30倍,但在非生长季因土壤呼吸所排放的碳量占全年因土壤呼吸所排放的碳量的21%左右。这与近期的研究结果非生长季因土壤呼吸而排放的碳占全年的14%~30%相一致[6]。因此,非生长季土壤呼吸是非生长季生态系统释放CO2的一个重要组成部分,显著地影响着区域碳收支和生态系统碳循环。目前关于非生长季土壤呼吸的研究并不多,已有的研究结果大多集中在极地苔原地区和亚高山森林[12]。他们的研究结果表明,极地苔原和森林冬季土壤呼吸速率分别为0.002~1.359和0.22~0.67μmol/(m2·s);土壤呼吸的 CO2释放量分别为0.55~26.37和22.4~152.0g C/m2[6,12]。本研究结果与其相一致。
3.2 非生长季土壤呼吸的温度敏感性
土壤呼吸的温度敏感系数(Q10)是表征呼吸作用的重要指标,它是指温度每增加10℃土壤呼吸增加的倍数,这一指标反映了温度变化后呼吸作用的变化规律,对于了解未来气候变化后土壤呼吸的响应具有一定的参考价值。对土壤呼吸温度敏感性的深刻理解不仅可以揭示生态过程对气候变化和人类活动引起的气候变化的响应和适应,还有助于改进全球碳循环模型,以更准确的计算生态系统碳循环和区域碳收支,为气候变化做出更准确的预测[13]。以往的研究得出,陆地生态系统土壤呼吸的Q10值变化在1.3~5.6之间,这取决于生态系统的类型及其地理分布[14]。总体上Q10与温度呈负相关,在温度上升相同幅度下低温地区比在高温地区有着更大的Q10。相关的野外实验表明,土壤呼吸的温度敏感性在非生长季要大于生长季,并且这种敏感性随着温度的升高而降低[15]。实验室培养实验的结果也表明土壤呼吸的温度敏感性随着温度的增加而下降,并认为这主要是温度的直接作用造成的[16]。相关的整合分析也证明这一结论[17]。然而这些结论都基本基于土壤水分不受限的情况而得出的。
本研究结果表明,非生长季具有较低的土壤呼吸温度敏感性,这和以往的大多数研究结果是不相一致的。这可能是由于Q10具有一定的水分依赖性,不同生态系统中水分对Q10的影响会有很大的差别[13],一般而言,土壤干旱会降低土壤呼吸的温度敏感性,一定范围内随着土壤含水量的增加,土壤呼吸对温度的敏感性也会增加[17]。水分会通过对基质扩散的影响而作用于土壤呼吸温度敏感性。当水分含量低时会抑制酶、呼吸底物以及微生物的活动,从而影响到土壤呼吸对温度的响应;而在水分不构成土壤中基本生物化学过程的限制条件时,温度升高会加快物质扩散,因此土壤呼吸的温度敏感性就较高[18-19]。同时实验室的培养实验也表明土壤含水量适宜时温度敏感性最高,而土壤含水量较低和较高时温度敏感性下降[16]。研究样地非生长季土壤水分基本在7%以下,相关研究表明土壤水分低于7%这个临界值会显著抑制土壤呼吸[15],本研究中非生长季较低的Q10值可能就归因于非生长季较低的土壤含水率。放牧与围栏封育对非生长季土壤呼吸温度敏感性的影响较小,自由放牧、3年围栏封育和5年围栏封育样地的Q10值分别为1.5,1.8和1.7。
相关的其他研究结果还表明,通过分析不同水分条件状态下土壤呼吸的温度敏感性得出,在中国温带草原地区温度升高对较湿润区域土壤呼吸的影响大于较干旱区域,这是因为在湿润的年份里土壤呼吸具有更高的温度敏感性[20];冻融交替过程可能降低土壤呼吸及其对温度的敏感性,这种土壤呼吸与温度变化的不一致说明在冻融过程中土壤呼吸还受到其他生态因子的影响,比如微生物活性、氮等营养元素的矿化以及土壤水分等[21]。因此土壤呼吸温度敏感性的大小不仅仅是由温度来控制的,而与之相关的其他因素所导致的土壤呼吸的直接响应,和由这些因素相关的间接过程都会作用于土壤呼吸的温度敏感性。
3.3 非生长季土壤呼吸与温度和水分的关系
传统的研究认为在青藏高原这样的高寒地区,温度是包括土壤呼吸在内的生态系统生物化学过程的主要控制因素[4]。但本研究结果表明,在非生长季土壤温度和土壤湿度对土壤呼吸的解释率相当,甚至土壤湿度对土壤呼吸的解释率更高(土壤温度对非生长季土壤呼吸的平均解释率为41.53%,土壤湿度对土壤呼吸的平均解释为41.93%)。而且一定的土壤体积含水率是保证土壤呼吸对温度响应的必要条件。这可能是由于在非生长季较低的土壤含水率,使得土壤呼吸更多地受到土壤含水率的限制。当土壤含水量较低时,土壤呼吸速率随温度变化不明显;但当土壤含水量较高时,土壤呼吸速率随温度的升高而增大[22]。在非生长季为维持一定的土壤生物化学过程,一定的体积含水率是必要的。这和大部分控制实验所得到的结果是相一致的,土壤呼吸通常对最限制它的因子产生响应,即当土壤湿度成为限制因子时,土壤呼吸对温度的敏感性也会降低[23]。这和近期的研究结果限制寒冷气候下的生命活动的因素是水分的可获取性,而不是温度是相一致的[24]。与此相关的研究得出:土壤温度在-5℃以上时均可以检测到土壤微生物的活性;在-10℃仍然有非冻结的土壤水分和土壤呼吸作用[25];甚至在-39℃时仍有土壤呼吸的作用[26]。所有这些研究结果都表明,冬季土壤呼吸可能受温度以外的其他因素的影响作用更大,例如本研究中的土壤水分就是一个主要的影响因子。土壤呼吸是一个受多因子影响的复杂CO2释放过程,温度是影响土壤呼吸的一个主要因子,有研究结果表明,温度只有在极高或者极低时才会限制到土壤呼吸。除此之外非生长季土壤呼吸还受到降雪,土壤微生物活性等的影响。
[1]陈骥,曹军骥,金钊,等.模拟增温对青海湖鸟岛高寒草原群落结构影响初步研究[J].干旱区资源与环境,2014,28(5):127-133.
[2]王建兵,张德罡,曹广民,等.青藏高原高寒草甸退化演替的分区特征[J].草业学报,2013,22(2):1-10.
[3]Klein J A,Harte J,Zhao X Q.Dynamic and complex microclimate responses to warming and grazing manipulations[J].Global Change Biology,2005,11(9):1440-1451.
[4]Cao G M,Tang Y H,Mo W H,etal.Grazing intensity alters soil respiration in an alpine meadow on the Tibetan plateau[J].Soil Biology & Biochemistry,2004,36(2):237-243.
[5]Riley W J.Influence of terrestrial ecosystems and topography on coastal CO2measurements:A case study at Trinidad Head,California[J].Journal of Geophysical Research,2005,doi:10.1029/2004JG000007.
[6]王娓,汪涛,彭书时,等.冬季土壤呼吸:不可忽视的地气CO2交换过程[J].植物生态学报,2007,31(3):394-402.
[7]Monson R K,Burns S P,Williams M W,etal.The contribution of beneath-snow soil respiration to total ecosystem respiration in a high-elevation,subalpine forest[J].Global Biogeochemical Cycles,2006,doi:10.1029/2005GB002684.
[8]方精云,郭兆迪.寻找失去的陆地碳汇[J].自然杂志,2007,29(1):1-6.
[9]熊沛,徐振锋,林波,等.岷江上游华山松林冬季土壤呼吸对模拟增温的短期响应[J].植物生态学报,2010,34(12):1369-1376.
[10]Zhou X,Wan S Q,Luo Y Q.Source components and interannual variability of soil CO2efflux under experimental warming and clipping in a grassland ecosystem[J].Global Change Biology,2007,13(4):761-775.
[11]Niu S,Wu M,Han Y,etal.Nitrogen effects on net ecosystem carbon exchange in a temperate steppe[J].Global Change Bi-ology,2010,16(1):144-155.
[12]Wang C K,Bond-Lamberty B,Gower S T.Soil surface CO2flux in a boreal black spruce fire chronosequence[J].Journal of Geophysical Research-Atmospheres,2002,doi:10.1029/2001JD000861.
[13]杨庆朋,徐明,刘洪升,等.土壤呼吸温度敏感性的影响因素和不确定性[J].生态学报,2011,31(8):2301-2311.
[14]Zhou T,Shi P,Hui D,etal.Global pattern of temperature sensitivity of soil heterotrophic respiration(Q10)and its implications for carbon-climate feedback[J].Journal of Geophysical Research,2009,doi:10.1029/2008JG000850.
[15]Luo Y Q,Wan S Q,Hui D F,etal.Acclimatization of soil respiration to warming in a tall grass prairie[J].Nature,2001,413:622-625.
[16]Leifeld J,Steffens M,Galego-Sala A.Sensitivity of peatland carbon loss to organic matter quality[J].Geophysical Research Letters,2012,39:doi:10.1029/2012GL051856.
[17]Kirschbaum M U F.The temperature dependence of organic-matter decomposition-still a topic of debate[J].Soil Biology &Biochemistry,2006,38(9):2510-2518.
[18]Almagro M,López J,Querejeta J,etal.Temperature dependence of soil CO2efflux is strongly modulated by seasonal patterns of moisture availability in a Mediterranean ecosystem[J].Soil Biology and Biochemistry,2009,41(3):594-605.
[19]陈思宇,于惠,冯琦胜,等.2002-2010年青藏高原植被含水量微波遥感动态监测[J].草业学报,2013,22(5):1-10.
[20]陈全胜,李凌浩,韩兴国,等.典型温带草原群落土壤呼吸温度敏感性与土壤水分的关系[J].生态学报,2004,24(4):831-836.
[21]Mikan C J,Schimel J P,Doyle A P.Temperature controls of microbial respiration in arctic tundra soils above and below freezing[J].Soil Biology and Biochemistry,2002,34(11):1785-1795.
[22]邓钰,柳小妮,闫瑞瑞,等.呼伦贝尔草甸草原土壤呼吸及其影响因子对不同放牧强度的响应[J].草业学报,2013,22(2):22-29.
[23]杨阳,韩国栋,李元恒,等.内蒙古不同草原类型土壤呼吸对放牧强度及水热因子的响应[J].草业学报,2012,21(6):8-14.
[24]Fisk M C,Schmidt S K,Seastedt T R.Topographic patterns of above-and belowground production and nitrogen cycling in alpine tundra[J].Ecology,1998,79(7):2253-2266.
[25]Brooks P D,Schmidt S K,Williams M W.Winter production of CO2and N2O from Alpine tundra:Environmental controls and relationship to inter-system C and N fluxes[J].Oecologia,1997,110(3):403-413.
[26]Panikov N S,Flanagan P W,Oechel W C,etal.Microbial activity in soils frozen to below -39℃[J].Soil Biology and Biochemistry,2006,38(4):785-794.
