拉萨河谷山地灌丛草地物种多样性随海拔升高的变化特征
2014-01-02罗黎鸣苗彦军武建双潘影土艳丽余成群赵延赵贯锋武俊喜
罗黎鸣,苗彦军,武建双,潘影,土艳丽,余成群,赵延,赵贯锋,武俊喜*
(1.西藏大学农牧学院,西藏 林芝860000;2.中国科学院地理科学与资源研究所 生态系统网络观测与模拟重点实验室拉萨高原生态试验站,北京100101;3.西藏自治区高原生物研究所,西藏 拉萨850001)
*物种多样性随海拔升高的变化特征是生物多样性研究的重要议题,研究物种多样性的变化特征及其环境控制因子是保护生物学研究的基础[1]。海拔梯度综合了气候、土壤、光照等多种环境因子,因此物种多样性随海拔升高的变化特征可以为预测全球气候变化和人类干扰背景下物种分布提供参考[2-4]。贺金生和陈伟烈[5]将物种多样性与海拔梯度关系系统归纳为5种形式:负相关、单峰型、U型、正相关和无规律性。物种多样性及其变化对草地生态系统结构和功能的稳定性有显著影响[6-7]。功能性状具有表征生态系统功能的生态指示作用,对生态系统产生实质性影响[8-9]。
20世纪90年代中期开始,中国山地植物物种多样性调查计划(PKU-PSD计划)对国内寒温带、温带、暖温带、亚热带、热带、干旱(半干旱)各类型生态系统山地物种多样性的变化特征进行了系列报道[10-12],而世界“第三极”青藏高原报道不多。近年青藏高原物种多样性随海拔升高变化特征的研究[13-14]多集中在高寒草甸区,如王长庭等[13]在青海玛沁县和段敏杰等[14]在西藏那曲县的研究表明,物种多样性随海拔的升高先增加后降低,即物种丰富度、均匀度和多样性在中间海拔梯度最大,验证“中间膨胀假说”。而针对青藏高原河谷地区山地灌丛草地的研究较为薄弱。“一江两河”(指雅鲁藏布江、拉萨河、年楚河)地区属于藏南及喜马拉雅中段生态安全屏障区中的雅鲁藏布江中游宽谷土地沙化和水土流失控制及经济重点发展亚区[15]。既是西藏最重要的山原宽谷区,也是西藏粮食主产区和农区畜牧业发展重点区域[16]。受气候变暖、过度放牧和樵采等人类活动的多重干扰[17-18],河谷地区天然草地退化形势严峻[19],致使水土流失日益加剧、生态环境破坏严重[20],从而破坏了生态和经济的良性循环[21],制约该地区的跨越式发展。生物多样性是生态安全屏障功能监测与评估的重要内容之一[22],研究灌丛草地物种多样性随海拔升高的变化特征可为西藏河谷农区畜牧业可持续发展以及生态安全屏障建设提供科学依据。
“一江两河”地区属高原季风温带半干旱气候区,年平均气温在2.4~8.2℃,年降水量300~500mm。土地面积6.65万km2,占西藏总面积的5.41%;天然草地4.75万km2,占该地区土地面积的71.44%;人口总量达108.1万,占西藏总人口的36%[23]。拉萨河谷是其重要山原河谷区,山体连绵,河谷纵宽,沟域众多。本研究以拉萨河谷典型的山地灌丛草地为样地,通过研究植物群落物种Shannon-Wiener多样性指数和功能性状(株高)Rao指数变化特征,验证其物种多样性随海拔升高的“单峰曲线”变化特征,探究功能性状随海拔升高的变化特征,初步探讨造成物种多样性与功能性状随海拔变化的原因,为该地区山地灌丛草地物种多样性的保护、生态恢复和生态安全屏障建设提供科学依据和理论基础。
1 材料与方法
1.1 研究区域概况
研究区位于拉萨河流域下游的藏南谷地南岸,处于念青唐古拉山脉中段南翼,隶属于西藏自治区拉萨市达孜县(29°29′~30°02′N、91°15′~91°41′E)。平均海拔4500m,年均温约为7.5℃,日照充足,年均日照时数为3065 h,年均降水量450mm左右,90%以上集中6-9月,属于高原季风温带半干旱气候[24]。样地位于新仓村沟域的阴坡上,土壤类型由下至上分别是亚高山草甸草原土、亚高山灌丛草甸土、高山草甸土;植被类型由下至上分别是亚高山草甸草原、亚高山灌丛草甸、高山草甸;灌丛优势种为微毛樱草杜鹃(Rhododendronprimuliflorum),草地优势种为高山嵩草(Kobresiapygmaea)、青藏苔草(Carexmoorcroftii)等。
1.2 试验方法
于2012年7月,在草地生长季生物量高峰期对达孜县新仓村沟域的山地灌丛草地地上生物量、物种多样性和功能性状沿海拔梯度进行调查分析。由山脚3992m至山顶4940m沿坡面按植被类型差异性,每隔150~300 m设置1个样地,共设5个样地,每样地内平行设3个采样区,每个采样区在约为5m×5m的区域随机选取3个面积为0.5m×0.5m的样方,共计9个重复。先记录每个样方各个物种的高度、盖度和密度,然后齐地面分种收集地上生物量,以65℃烘干48h称重;每个采样区随机选取1个面积10m×10m的大样方,记录样方中灌丛数量、盖度,再随机记录10株灌丛的灌高、灌幅宽、灌幅窄,然后随机剪取其中3株,以65℃烘干48h称重。
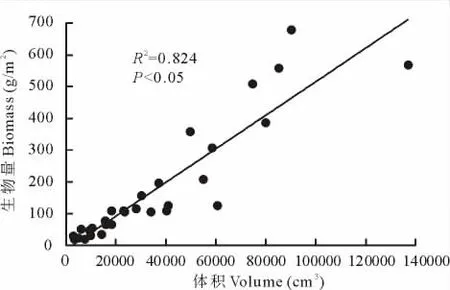
图1 灌丛生物量与灌丛体积的关系Fig.1 Relationship between aboveground biomass and shrub volume
1.3 数据处理
假定灌丛冠幅为长方体,灌丛体积(V)=灌高×灌幅宽×灌幅窄。灌丛的生物量(SB)与体积(V)建立线性回归方程(图1):y=0.0053x-12.728(R2=0.824,P<0.05)。采样区灌丛生物量(SB)=(d/n)×ym=(d/n)×(y1+y2+…+y10)/10,式中:d为采样区灌丛总株数;n为采样区面积,ym为单株平均生物量。

本研究中,物种多样性的测定采用物种丰富度指数、Shannon-Wiener多样性指数(H)表征。物种丰富度指数用9个0.25m2样方内出现的物种数表示;Shannon-Wiener多样性指数H=-∑PilnPi,式中,Pi为物种的相对重要值。
功能性状(functional trait)采用Rao指数[25]表示,R,式中,S为样方中的物种数;Pi、Pj分别为第i、j个物种在群落中相对盖度;dij为种i与种j之间的性状差额,本研究以株高差距计算。株高是植物对综合生境条件长期适应而在外貌上表现的重要特性之一,代表资源竞争力,反映生态系统功能[26]。
1.4 数据分析
将各海拔梯度上所采的样方数据计算均值,通过SPSS 19.0软件,利用单因子方差分析(ANOVA)的最小显著差数法(LSD)对不同海拔梯度下各指标进行比较和差异显著性检验,作图采用Origin 8.0软件。
2 结果与分析
2.1 随海拔升高山地灌丛草地植物群落地上生物量及其盖度的变化特征
随着海拔的升高,草地的盖度(71±14)%和生物量(49.18±19.17)g/m2差异不显著(P>0.05);灌丛的盖度和生物量呈先增加后降低的变化趋势;山体植物群落总盖度和总生物量呈先增加后降低的趋势。海拔4050m处无灌丛发育,草地的盖度和生物量分别为70%和53.65g/m2(表1)。海拔4550m处草地和灌丛的盖度和生物量最高,草地分别为77%和63.40g/m2;灌丛分别为40%和1104.42g/m2。海拔4550m处灌丛的2个采样区4470和4645m盖度分别为55%和25%,生物量分别为1893.06和315.79g/m2。

表1 随海拔升高山地灌丛草地植物群落地上生物量及其盖度的比较Table 1 Comparisons of aboveground biomass and total coverage of shrub grassland communities along an altitudinal gradient
2.2 随海拔升高山地灌丛草地植物物种组成及其重要值的变化特征
随着海拔升高山地灌丛草地植物群落物种组成及重要值存在一定的差异(表2)。高山嵩草为优势种,且重要值(0.591±0.034)变化不大。海拔4050m处共优种狼毒,次优势种青藏苔草,重要值分别为0.542和0.307。海拔4200m处共优种狼毒,次优势种青藏苔草,劲直黄芪,重要值分别为0.405,0.272和0.244,出现禾草类植物,莎草类和杂草类重要值总和在该梯度的比例明显增加。海拔4350m处次优势种青藏苔草,紫花针茅,重要值分别为0.268和0.223,杂草类中,劲直黄芪,狼毒不再出现,垫状点地梅开始出现,其重要值为0.092。海拔4550和4850m处,杂草类的重要值表现出增加的趋势。
2.3 拉萨河谷山地灌丛草地植物群落物种多样性随海拔升高的变化特征
随着海拔升高山地灌丛草地植物群落物种丰富度、多样性、功能性状Rao指数差异显著(P<0.05)(图2)。物种丰富度(图2A)随海拔的升高先增加后降低,整体呈“单峰曲线”变化特征。海拔4050m处物种数最少(13种),海拔4200m处物种最多(23种)。Shannon-Wiener指数(图2B)随海拔的升高先增加后降低,整体呈“单峰曲线”变化特征,在海拔4350m处最高(1.700),在海拔4850m处最低(1.479)。功能性状Rao指数(图2C)随海拔的升高而降低,在海拔4050m处最大(1.624),在海拔4850m处最小(0.540)。

表2 随海拔升高山地灌丛草地植物物种组成及其重要值Table 2 Species composition and important value of the shrub grasslands along an altitudinal gradient
3 讨论
3.1 山地灌丛草地植物群落随海拔升高的变化特征
青海玛沁县[13]和西藏那曲县[14]的研究表明,高寒草甸植物群落生物量和盖度随海拔的升高先增加后降低。岷江上游干旱河谷[27]的研究表明,灌丛群落随海拔的升高而增加。本研究表明,灌丛草地的灌丛生物量和盖度随海拔升高为先增加后降低;而草地生物量和盖度随海拔升高无显著变化。灌丛受海拔因素分异明显,而草地受海拔因素分异不明显,可能是海拔梯度引起的环境因子变化对灌丛生长发育影响显著,而草地群落由于灌丛群落小生境的庇护,影响不显著。Badano等[28]在安第斯山脉的研究表明,在高山环境下护士植物(nurse plant)对微环境的改善能频繁减弱环境压力对植物的负面影响。灌丛一方面物理隔离了畜群的采食和践踏;一方面改善了局部光、温、水因子的分配,为草地群落的发育提供了空的生态位,减少了物种间的竞争。发育在山体中上部海拔4350~4700m的陡坡灌丛带对防控该地区山地水土流失发挥了重要的作用。

图2 物种丰富度、香侬-维纳指数以及以株高测算的功能性状的Rao指数随海拔升高的变化Fig.2 Variations of species richness,Shannon-Wiener index and Rao’s index based on functional trait-height along an altitudinal gradient
拉萨河谷山地灌丛草地的灌丛种类和数量与岷江、伊犁河谷[27,29]差异很大,可能是海拔和气候差异较大造成的。本研究样地灌丛种类只有3种,优势种微毛樱草杜鹃占据绝对优势,伴生种雪层杜鹃(R.nivale)和金露梅(P.fruticosa)零星分布。草地群落组成及其重要值与段敏杰等[14]藏北高寒草地的结果不一致。共优种为高山嵩草和青藏苔草,伴生种为圆穗蓼、钉柱委陵菜,各物种重要值差异不显著,可能是由各自的生态位[30]所决定。海拔较低处(4050~4200m)杂草类中的劲直黄芪,狼毒出现且物种重要值相对较高。这说明毒杂草增加、群落结构变化、植被已呈退化趋势,很大程度上是人类频繁活动、牲畜采食行为造成的。但海拔4200m处灌丛小生境为物种繁育起到了庇护作用,草地物种数并没有减少反而最高[28]。由此推断,拉萨河谷山地灌丛草地中灌丛对草地生态系统结构和生产功能的稳定性具有重要作用。
3.2 物种多样性和功能性状随海拔升高的变化特征
青藏高原草甸区的研究[13-14]表明,物种多样性随海拔的变化呈“单峰曲线”变化特征,即随海拔梯度的升高,物种丰富度、Shannon-Wiener多样性指数、Pielous均匀度指数先增加后降低。疏勒河上游地区[31]不同类型草地植被物种多样性随海拔的变化呈“单峰曲线”变化特征,验证“中间膨胀假说”。受焚风效应的缘故,岷江上游干旱河谷[32]物种多样性随海拔升高而降低。伊犁河谷北坡[29]植物群落多样性呈一种特殊的双峰变化特征,即随海拔梯度的升高,物种丰富度、Shannon-Wiener多样性指数、Pielous均匀度指数呈先增加后降低再增加的特征,这是由于研究区存在特殊的“逆温层”地带,本质上同样符合“中间膨胀假说”。本研究结果验证了物种多样性随海拔升高呈“单峰曲线”变化特征,可能是低海拔受水分制约,高海拔受温度制约,中海拔常有云雾覆盖、水热条件相对均衡等气候差异对物种多样性分布特征有显著影响。
以株高测算的功能性状Rao指数随海拔的升高而降低,充分反映草地植被株高随海拔分异明显。低海拔Rao指数大,植被对资源利用的互补性强,株高功能性状趋异;高海拔Rao指数小,植被对资源利用的互补性弱,株高功能性状趋同。随着海拔的升高环境恶劣程度加剧,不适应的物种被淘汰,适应的物种功能趋同,这是环境对物种选择的结果[33]。山地灌丛草地沿海拔梯度升高,物种多样性呈“单峰曲线”变化特征,以株高测算的功能性状Rao指数则呈下降趋势,这既是物种适应不同海拔的温度、水分和光照等要素组合的综合结果,也是物种共存和相互作用的综合结果。
4 结论
拉萨河谷山地灌丛对维持草地生态系统物种多样性、生产力和稳定性起一定作用。随海拔升高灌丛生物量和盖度呈先增加后降低的变化趋势,但草地盖度(71±14)%和生物量(49.18±19.17)g/m2变化不明显(P>0.05)。随海拔升高群落物种组成存在差异,但共优种高山嵩草(0.591±0.034)和青藏苔草(0.326±0.061)的优势度无显著变化(P>0.05)。随海拔升高草地物种丰富度、Shannon-Wiener多样性指数呈先增加后降低的“单峰曲线”变化特征;以株高测算的草地群落功能性状Rao指数随海拔升高而降低。物种丰富度、功能性状以及地上净初级生产力沿海拔升高的变化特征存在差异,为维持高原河谷山地灌丛草地的生态安全,未来应重视三者相互关系及其生态学机制的相关研究。
该地区山地灌木种类仅有3种,其中微毛樱草杜鹃占据绝对优势,所以该地区灌丛物种多样性保护不是首要的,但发育在山体中上部海拔4350~4700m的陡坡灌丛带对防控该地区山地水土流失发挥了重要作用。所以在日常生产生活中,人类樵砍灌丛行为要适度,要保证灌丛群落的完整性和再生产能力,以免灌丛群落结构和功能严重受损,进而导致草地生态系统物种多样性减少、生态系统稳定性变差、生态安全屏障功能减弱。
[1]Noss R F.Indicators for monitoring biodiversity:a hierarchical approach[J].Conservation Biology,1990,4(4):355-364.
[2]Wu J,Shen Z,Zhang X.Precipitation and species composition primarily determine the diversity-productivity relationship of alpine grasslands on the Northern Tibetan Plateau[J].Alpine Botany,2014,124(1):13-25.
[3]Gaston K J.Global patterns in biodiversity[J].Nature,2000,405:220-227.
[4]Oommen M A,Shanker K.Elevational species richness patterns emerge from multiple local mechanisms in Himalayan woody plants[J].Ecology,2005,86(11):3039-3047.
[5]贺金生,陈伟烈.陆地植物群落物种多样性的梯度变化特征[J].生态学报,1997,17(1):91-99.
[6]Tilman D,Reich P B,Knops J,etal.Diversity and productivity in a long-term grassland experiment[J].Science,2001,294:843-845.
[7]江小雷,岳静,张卫国,等.生物多样性,生态系统功能与时空尺度[J].草业学报,2010,19(1):219-225.
[8]Díaz S,Cabido M.Vive la difference:plant functional diversity matters to ecosystem processes[J].Trends in Ecology & Evolution,2001,16(11):646-655.
[9]孟婷婷,倪健,王国宏.植物功能性状与环境和生态系统功能[J].植物生态学报,2007,31(1):150-165.
[10]方精云,沈泽昊,唐志尧,等.“中国山地植物物种多样性调查计划”及若干技术规范[J].生物多样性,2004,12(1):5-9.
[11]唐志尧,方精云.植物物种多样性的垂直分布格局[J].生物多样性,2004,12(1):20-28.
[12]方精云.探索中国山地植物多样性的分布规律[J].生物多样性,2004,12(1):1-4.
[13]王长庭,王启基,龙瑞军,等.高寒草甸群落植物多样性和初级生产力沿海拔梯度变化的研究[J].植物生态学报,2004,28(2):240-245.
[14]段敏杰,高清竹,郭亚奇,等.藏北高寒草地植物群落物种多样性沿海拔梯度的分布格局[J].草业科学,2011,28(10):1845-1850.
[15]钟祥浩,刘淑珍,王小丹,等.西藏高原国家生态安全屏障保护与建设[J].山地学报,2006,24(2):129-136.
[16]钟祥浩,刘淑珍,王小丹,等.西藏高原生态安全研究[J].山地学报,2010,(1):1-10.
[17]韩国军,王玉兰,房世波.近50年青藏高原气候变化及其对农牧业的影响[J].资源科学,2011,33(10):1969-1975.
[18]钱拴,毛留喜,侯英雨,等.青藏高原载畜能力及草畜平衡状况研究[J].自然资源学报,2007,22(3):389-397.
[19]蔡晓布.西藏“一江两河”地区土壤退化特征[J].土壤肥料,2003,40(3):4-7.
[20]陶和平,高攀,钟祥浩.区域生态环境脆弱性评价——以西藏“一江两河”地区为例[J].山地学报,2006,24(6):761-768.
[21]黄黔.我国的生态建设与生态现代化[J].草业学报,2008,17(2):1-8.
[22]孙鸿烈,郑度,姚檀栋,等.青藏高原国家生态安全屏障保护与建设[J].地理学报,2012,67(1):3-12.
[23]西藏自治区第六次全国人口普查领导小组办公室.西藏自治区2010年人口普查资料[M].北京:中国统计出版社,2012:6.
[24]赵海珍,李文华,马爱进,等.拉萨河谷山地灌丛草地生态系统服务价值评价——以拉萨达孜县为例[J].草业科学,2011,28(12):27-31.
[25]Leps J,De Bello F,Lavorel S,etal.Quantifying and interpreting functional diversity of natural communities:practical considerations matter[J].Preslia,2006,78(4):481-501.
[26]Vojtech E,Loreau M,Yachi S,etal.Light partitioning in experimental grass communities[J].Oikos,2008,117(9):1351-1361.
[27]陈泓,黎燕琼,郑绍伟,等.岷江上游干旱河谷灌丛生物量与坡向及海拔梯度相关性研究[J].成都大学学报(自然科学版),2007,26(1):14-18.
[28]Badano E I,Marquet P A,Cavieres L A.Predicting effects of ecosystem engineering on species richness along primary productivity gradients[J].Acta Oecologica,2010,36(1):46-54.
[29]田中平,庄丽,李建贵.伊犁河谷北坡垂直分布格局及其与环境的关系——一种特殊的双峰分布格局[J].生态学报,2012,32(4):1151-1162.
[30]Pulliam H R.On the relationship between niche and distribution[J].Ecology Letters,2000,3(4):349-361.
[31]陈生云,刘文杰,叶柏生,等.疏勒河上游地区植被物种多样性和生物量及其与环境因子的关系[J].草业学报,2011,20(3):70-83.
[32]林勇明,王道杰,周麟,等.岷江干旱河谷植物群落分布特征及梯度效应[J].福建林学院学报,2009,29(4):332-335.
[33]Wu J,Zhang X,Shen Z,etal.Grazing-exclusion effects on aboveground biomass and water-use efficiency of alpine grasslands on the Northern Tibetan Plateau[J].Rangeland Ecology & Management,2013,66(4):454-461.
