黄土区苜蓿人工草地群落生态与生产功能演替
2014-01-02王书转郝明德普琼吴振海
王书转,郝明德,普琼,吴振海
(1.中国科学院水利部水土保持研究所,陕西 杨凌712100;2.中国科学院大学,北京100039;3.西藏自治区农业技术推广服务中心,西藏 拉萨850000;4.西北农林科技大学生命科学学院,陕西 杨凌712100)
人工草地建设是现代生态学的重点研究领域[1]。黄土高原地区是我国水土流失最为严重的地区,植被建设是遏制水土流失和防止土壤退化的重要途径[2]。改善和优化黄土高原地区脆弱的生态环境很大程度上有赖于对草地植被的保护、发展与建设[3],再加上黄土高原地区干旱少雨,土壤营养条件差,而该区草地面积与分布范围远大于森林,草地覆盖和保护土地的功能又显著优于荒漠,故建植多年生人工草地不仅有利于该区农牧业经济可持续发展,同时也有助于指导退化草地生态系统恢复和重建,改善和优化该地区生态环境[4-7]。多年生豆科牧草紫花苜蓿(Medicagosativa),在黄土高原地区种植历史悠久,生态适应性强,耐旱、耐土壤贫瘠,适口性好,营养价值高,具有很好的经济效益[8]。紫花苜蓿不仅有利于畜牧业发展,它还具有土壤改良和水土保持的功能。甘肃天水水土保持站通过9年的观测资料证明,苜蓿地要比作物地减少土壤冲刷1.6倍;绥德站的观测资料也表明,种植苜蓿小区要比种植谷子减少土壤冲刷5~12倍[2],可见紫花苜蓿人工草地具有较好的生产和生态功能,如何将其建设成为一种生态上健全可靠、经济和生产上又合理可行的优化生态——生产人工草地将具有非常强的现实指导意义[3]。本研究基于时间序列研究方法对苜蓿人工草地群落生态和生产功能演替过程进行了研究,以期为退化草地恢复与重建提供科学依据,同时延长苜蓿人工草地的有效利用年限,更好地发挥苜蓿草地的生产、生态功能,实现其生产、生态功能的协调和可持续发展。
1 材料与方法
1.1 研究区概况
试验区域位于甘肃省镇原县上肖乡,地处北纬35°30′,东经107°29′,海拔1200m,属黄土高原沟壑区,以塬地为主。属北方半湿润易旱区,降水主要集中在7-9月,季节短而集中,年均降水量520mm,年平均气温8.7℃,年辐射量554.3~565.2kJ/cm2,≥0℃年积温3435℃,≥10℃年积温2722℃,全年无霜期165d。主要土壤类型为黑垆土和黄绵土,土层深厚。研究区域为黄土高原旱塬区,水资源极度贫乏,塬高水低,不利于水资源的存贮,农业耕作主要利用天然降水,属典型的旱作农业区。
1.2 试验设计
共设置10块试验样地,每块样地大小约10m×20m,样地平整均一,距离很近,即使最大距离也在1km范围之内,试验区域为塬面,地形平坦,起伏很小,可以不考虑样地海拔差异所带来的影响,土壤均为黑垆土,各样地立地条件较一致,具有可比性。选取的10块样地最初是耕地,后被当地居民弃耕,为了不荒废耕地同时又便于饲养牲畜,当地居民便在这些弃耕地上种植当地乡土品种紫花苜蓿,种植期间从不施肥,也无田间管理措施,只在每年收获季节茬割2次,比较适合研究平坦塬地上人工草地的演替变化。样地确立之后与当地居民签订协议,同时确定每块苜蓿草地的准确种植年限。本研究数据于2010年6月底、9月初采集种植时间分别为2,3,4,5,6,7,8,16,26和31年的苜蓿草地进行研究。在每块苜蓿草地随机选取4个1m×1m样方(远离样地边缘50cm以减少边缘效应所带来的试验误差),记录每个样方内的植物种类、株数、株高以及每种植物在样方内的株数或丛数、频度和盖度。
1.3 数据计算方法
群落中植物种的重要值(IV)=1/3[相对频度(%)+相对盖度(%)+相对密度(%)][9-12],其中,相对密度为样地内各种植物的个体数占全部植物种个体数的百分比。相对盖度为群落中某一物种的种盖度占所有种盖度之和的百分比,盖度以离地面1英寸(2.54cm)高度的断面积计算[9]。相对频度为各种植物在全部样方中的频度与所有植物种频度和之比。
物种多样性测定选用 Margalef指数R1、Gleason指数R2表征物种丰富程度、Shannon-Wiener指数(H′)和Pielou指数(J′)表征群落物种多样性特征[12-14]。

式中,R为物种丰富度指数,S为群落中的植物种类数,N为群落中所有种的总个体数,A为单位面积。

式中,H′为多样性指数,S为群落中的植物种数量,Pi为第i种植物的重要值占所有植物种重要值总和的比值。

式中,J′为均匀度指数,H′为多样性指数,S为群落中的植物种数量。

式中,C′为生态优势度,N为群落中所有种的重要值总和,ni为第i种植物的重要值。
草地生产力测算:将每个样方中所有草的干重相加为总的地上生物量。
1.4 数据处理
通径分析可处理较为复杂的变量关系,如自变量与因变量之间的相关关系或因果关系,或者某些自变量之间的间接关系对因变量产生的影响等,从而便于对各因子变量之间的关系进行解释,因此使用较广。本研究运用SPSS软件对数据进行通径分析和相关分析。
2 结果与分析
2.1 群落生态功能演替
2.1.1 群落演替阶段分析 建群种的更替是草地植被演替阶段的象征,优良牧草和杂类草之间的竞争将决定人工草地群落演替的方向[15-16]。重要值是综合数量指标,它以综合数值表示植物物种在群落中的相对重要性。表1按物种重要值排序列出了不同生长年限苜蓿草地群落主要物种组成及其密度值,结果显示,苜蓿种群重要值随其生长年限呈先增加后降低的变化趋势,在第6年达到最高值,之后苜蓿种群重要值开始逐渐降低,到31年左右达到最低,由此将苜蓿人工草地群落生态功能演替过程大致分为3个阶段,即紫花苜蓿草地阶段、苜蓿人工草地向长芒草天然草地演替的过渡阶段、长芒草次生天然草地阶段。
1)紫花苜蓿人工草地演替阶段,大致包括2~6龄苜蓿草地,在此演替阶段,不同生长年限的苜蓿草地群落的优势种皆为苜蓿,其重要值和密度皆呈上升趋势,附地菜为群落的亚优势种,具有相对较大的重要值,茵陈蒿、赖草、夏至草等为主要伴生种。
2)苜蓿人工草地向长芒草天然草地演替的过渡阶段,大致包括8~31龄,群落的优势种仍为苜蓿,而各伴生种附地菜、夏至草、绢毛委陵菜、五爪金龙和长芒草等种间竞争较为激烈,密度和重要值变化复杂,较为明显的是附地菜逐渐演变为群落的主要伴生种,长芒草在16年之后开始大量入侵人工草地,且地位变化很快,重要值开始上升,直到31龄达到最高,最终替代苜蓿成为群落的优势种。
3)长芒草次生天然草地阶段,31龄之后,群落结构变化明显,苜蓿草地已经完全退化演替为长芒草群落,群落的优势种已被长芒草所代替,苜蓿则演变为亚优势种,形成长芒草+苜蓿+五爪金龙为主的群落类型。
从上述演替进程可以看出,种群组成在不同的阶段变化显著,杂类草之间的竞争较为复杂,这可以从主要伴生种附地菜、长芒草、五爪金龙、夏至草和绢毛委陵菜等重要值和密度值的变化趋势上看出来。重要值降低,表明该种植物在群落中的地位和作用降低。随苜蓿生长年限的延长,附地菜的重要值下降趋势明显,但又不同于其他伴生种,它在不同的演替阶段均为重要的伴生种,这是由于附地菜繁殖力较强,且耐旱、耐土壤贫瘠,适合黄土高原特殊的自然环境,因此成为研究区域苜蓿人工草地主要的入侵杂草。五爪金龙和绢毛委陵菜在群落演替后期重要值较高,成为该时期主要的竞争杂草。与之相反的是蒙古蒲公英和夏至草,它们的重要值在群落演替的前期和中期较高,为群落前期和中期的主要竞争杂草。

表1 苜蓿人工草地群落主要物种组成、密度与重要值的动态变化Table 1 The dynamics changes of main species’components,density and importance value of alfalfa artificial grassland
苜蓿人工草地群落物种组成的演替过程表明,随演替年限的增加,群落结构的复杂化程度不断提高,与天然植被群落结构的相似度越来越大,说明群落演化逐步倾向于生态稳定性群落[11],人工草地“自然化”趋势越来越大。
2.1.2 种群演替分析 种群密度、高度、盖度和频度是植物种最基本的数量特征,这些数量特征的动态变化情况可在很大程度上反映不同种群在群落内的地位、作用、竞争力以及群落的演替特征。在苜蓿人工草地群落演替过程中,长芒草和苜蓿种群的变化都极为显著(表1,表2)。
1)苜蓿种群的变化。随着演替时间的增加,苜蓿种群的高度、盖度、密度和重要值都表现为先上升后下降的变化趋势。群落演替的第6年,苜蓿种群的数量特征值均达到最大值,重要值、高度、密度、盖度增幅分别达87.26%,18.35%,29.28%和56.41%。随苜蓿植株生长速率的不断加快,苜蓿植株长势逐渐旺盛,种群盖度增幅较为明显、其次是密度和高度,这都使得苜蓿具有较强的资源利用能力和竞争力,在群落内比率增大,逐渐占据优势地位,重要值也相应地增大,从而使杂类草尤其是植株高大杂类草生长受到抑制。从第6年开始苜蓿种群各数量特征值逐年减少,且随着人工草地退化程度的加重下降显著。群落演替的第26年,苜蓿种群的数量特征值均降到最低,重要值、高度、密度、盖度降幅分别达58.47%,37.03%,60.60%,60.33%。到群落演替的第31年,苜蓿种群完全退化,草地群落内不但杂草种类繁多,且大多数为植株较高大的植物,如长芒草。

表2 黄土区苜蓿人工草地种群动态变化Table 2 The population dynamics changes of alfalfa grasslands on the Loess Plateau
2)长芒草种群的变化。虽然长芒草仅在16龄苜蓿草地出现,但其种群的重要值、高度、盖度、密度和频度增长很快,增幅分别为230.10%,36.27%,157.39%,560%,36.27%。长芒草为黄土高原原生地带性植被,为黄土高原的顶级演替群落[12],这与长芒草自身的生物特性有关,其生态幅度比较广,生态适应性强,容易适应不同的生境条件,并形成不同植物群落的优势和建群种,所以虽然它仅在苜蓿演替后期出现,却对苜蓿草地具有很强的入侵能力,能排除其他杂草,替代苜蓿而成为群落的优势种,引起苜蓿草地退化。
2.1.3 物种多样性和生态优势度的动态变化 植物群落结构动态变化能在一定程度上反映植被演替进程的特点[17]。随演替年限延长,苜蓿草地群落的多样性指数、均匀度指数和生态优势度指数的变化趋势基本一致,均呈先上升后下降的变化趋势,丰富度指数的变化趋势则相反,先下降后上升 (图1)。群落演替前期(2~6年),是建群种苜蓿生长旺期,苜蓿重要值、高度、盖度和密度不断增大,竞争力越来越强,充分利用光、热、水等资源,优势度相对增加,建群种的地位逐步提高,生态优势度指数稳步上升;因为人工草地不同于天然草地,其建群种是人为建植,所以其他杂草是随着草地生长时间而逐渐入侵,所以群落的多样性指数也呈上升趋势。随着苜蓿竞争能力的增强,其对杂类草的抑制也越来越强,致使群落植物种类数降低,而群落中所有种的个体数,尤其是苜蓿的株树越来越多,最终导致均匀度指数逐渐提高,而丰富度指数逐渐下降,群落的多样性指数、均匀度指数和生态优势度指数在草地6龄时达到最大,分别为5.23,2.95,0.53,相反,丰富度指数达到最小值3.00。群落演替中期(6~16年),苜蓿种群的重要值开始逐渐下降,生态优势度指数也随之逐渐减小,使得杂类草大量入侵,此时群落种间竞争尤其是杂草之间的竞争和抑制作用非常强烈,故群落多样性指数、均匀度指数和优势度指数呈缓慢下降趋势,物种丰富度指数则逐年增加,草地出现退化态势。苜蓿草地建植16年之后,生态适应强的长芒草开始入侵,苜蓿种群重要值迅速降低,生态优势度指数也快速下降,种群之间呈此消彼长的态势,致使多样性指数、均匀度指数下降较快,而丰富度指数快速上升,草地快速退化。上述变化趋势表明,没有田间管理措施的情况下,人工苜蓿草地盛产期时间短暂,草地的生产力稳定性差[11]。
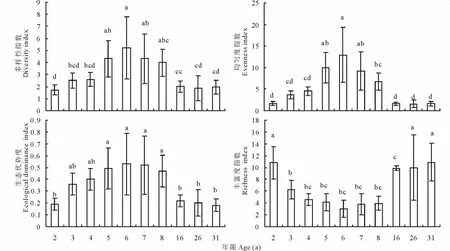
图1 群落特征值的变化Fig.1 Changes of community characteristic values
2.2 群落生产功能的动态变化
2.2.1 生物量的动态变化 在一定条件下,植被生物量的动态变化也可反映人工草地演替的方向及其退化状况。2茬苜蓿草地生物量、年生物量的变化趋势皆是先升后降(图2),且变化明显。草地演替的2~6年,苜蓿草地生物量随演替时间而增大,至6龄时达到最大,其中第1茬生物量为777.3g/m2,比2龄334.8g/m2增加了132.2%,第2茬生物量为304.3g/m2,比2龄141.4g/m2增加了115.3%。年生物量从2龄476.2g/m2上升到6龄995g/m2。6龄草地之后,草地生物量逐渐降低,显示出明显的退化趋势,演替至31年,植被生物量减至最低值76.3g/m2,退化原因主要是由于多年生杂草如长芒草等的不断入侵,以及土壤养分、水分供应不足。6年时间即达到生物量高峰也反映了苜蓿人工草地生产的不稳定性[11]。
2.2.2 地上生物量与群落数量特征之间的通径分析
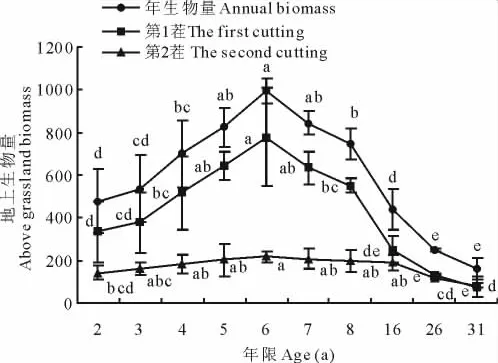
图2 苜蓿草地地上生物量的动态变化Fig.2 The dynamic changes of above vegetation biomass in alfalfa grassland
为了进一步分析苜蓿人工草地影响地上生物量的因素重要性大小,特选取了苜蓿种群重要值、生态优势度、多样性指数和长芒草种群重要值进行通径分析,结果(表3)显示:从相关系数来看,苜蓿种群重要值与地上生物量相关系数最大,其次是群落生态优势度和多样性指数,长芒草种群重要值与地上生物量呈负相关。从各因素对地上生物量的直接通径系数大小可以看出,多样性指数对地上生物量的影响最大,直接通径系数达0.536,众多学者研究也表明人工草地退化演替过程中多样性对生产力的影响比较显著[16,18-21]。
进一步对苜蓿草地植物群落多样性与生产力之间的相关性进行分析,结果表明(图3),物种多样性和生产力二者之间的相关性随草地生长年限延长呈“倒V”型变化趋势,6龄苜蓿人工草地二者相关性最高。建植苜蓿草地初期,苜蓿处于人为建植优势,群落物种多样性与生产力之间的变化趋势较为一致(图1,图3),故二者相关系数越来越大,相关性也越来越高。6龄之后,苜蓿草地群落多样性和生产力之间的相关性表现为由强变弱,31龄苜蓿草地二者的相关系数r仅为0.12。苜蓿人工草地植物群落生产力水平最高时多样性也达到最高,可见6龄苜蓿草地是生态和生产功能的转折点,是苜蓿草地管理的关键阶段。

表3 群落数量特征值与年地上生物量之间的通径系数及相关系数Table 3 Path coefficients and correlation coefficients between the population characteristic value and the annual above grassland biomass
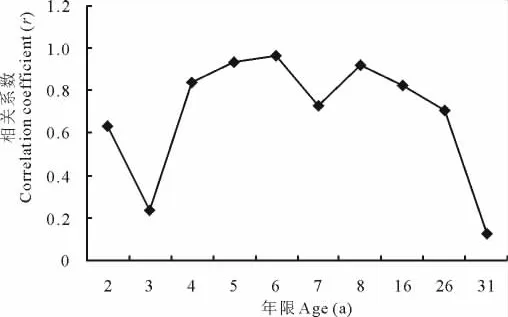
图3 苜蓿草地物种多样性与生产力之间相关系数动态变化Fig.3 The dynamics changes of the correlation coefficients between the diversity index and the annual biomass of the alfalfa grassland grassland
3 结果与讨论
人工草地是人为建植的植物群落,由单一外来物种构成,草地建植初期是一种“生产稳定性”群落,随草地年限的延长、土壤条件的变化以及乡土草种的侵入必然会增强群落的物种多样性,加速人工草地的退化,使得草地逐渐进入一种“生态稳定性”群落[6,11]。这2种功能对黄土高原地区都是有利的。种植人工草地旨在发展畜牧业,提高当地居民的经济收入,同时,退耕还草利于水土保持,又是西部地区生态环境建设的主要内容,因此需要将人工草地群落的“生产”和 “生态”功能有机结合,农民的经济利益和国家生态建设有机结合,实现生态效益和经济效益兼顾。
3.1 延缓人工草地的生态功能
从苜蓿草地群落的演替分析可以看出,6龄人工草地苜蓿重要值和密度均达到最大值,此阶段为紫花苜蓿人工草地演替阶段,是人工草地的生长旺期;之后重要值和密度均开始下降,苜蓿草地开始进入衰退阶段,优势种逐渐由苜蓿向天然草地优势种长芒草过渡;31年之后群落完全退化,完全演替为长芒草次生天然草地。
本研究中苜蓿草地具有的6年左右生长旺期与众多学者的研究结论一致,如李裕元等[11]对黄土高原北部森林草原气候区紫花苜蓿草地退化过程研究,另外还有其他学者对不同类型草地的研究也得到类似结论,如张耀生和赵新全[12]对青藏高寒牧区多年生人工草地持续利用的研究、周华坤等[6]对高山草甸垂穗披碱草(Elymusnutans)人工草地群落、王长庭等[22]、史惠兰等[23]对三江源区不同建植年代人工草地群落的研究等。从苜蓿人工草地的生态功能来看,经历短暂的生长盛期便开始向次生天然草地过渡,表明人工草地若无相应的田间管理和施肥、灌溉等措施,的确有助于加速其向天然草地过渡,从而更好地发挥其保护环境、防治水土流失的生态功能。
本研究与其他学者研究不同的是,李裕元等[11]的研究中,紫花苜蓿在第10年已完全退化为长芒草群落,而本研究则可以持续到31年才完全退化,这可能是研究区域差异和土质差异造成的,李裕元选择的是黄土高原北部中温带森林草原气候区坡耕地,水土流失严重,而本研究选择区域为平坦塬地,原本是较好的适耕地,区域环境和土质条件存在较大差异。对于紫花苜蓿人工草地长时间序列演替分析的研究不多,苜蓿草地向天然次生草地过渡的时间也不一致,但相同的是过渡阶段时期,优势种苜蓿的重要值逐渐降低,其他杂草开始争取其自身的优势,故种间竞争也最激烈。本研究中6龄草地之前主要的植物种类(按重要值大小分类)有10种,茵陈蒿、赖草、小花唐芥、灰绿藜、阿尔泰狗娃花、飞廉等在演替过程中基本全被竞争抑制,其他还有播娘蒿、葛缕子在演替过渡阶段也被竞争抑制,最后剩下的只有附地菜、五爪金龙、绢毛委陵菜、长芒草等竞争能力较强的植物,故过渡阶段群落的物种多样性指数较高,从生态角度来看,种群竞争是演替的一部分,且有利于草地生态系统的稳定。
群落内主要物种的数量特征和重要值的时间动态变化可在很大程度上反映该种群在群落内地位、功能、作用、竞争力的变化以及群落的演替特征。在苜蓿人工草地群落经过31年演替之后,苜蓿种群退化极为显著,密度、高度、盖度、重要值都降为最低,而长芒草的各数量特征值开始超过苜蓿,最终向次生天然长芒草草地演替,群落生态功能趋于稳定。
从上述分析可以看出,6龄草地生态功能稳定而强大,但这种稳定只是一种“暂稳态”,如果仅从苜蓿水土保持发挥角度来讲,要延缓这种暂稳态则需在6~31龄草地适当施加人为扰动和影响;如果从植被恢复与重建角度来讲,也可任其自然演替,愈来愈趋向“人工草地自然化”,在一定程度上这种“暂稳态”将有利于加速人工草地天然化。
3.2 延长人工草地的生产功能
从生产功能角度来看,苜蓿草地生长旺盛期短不利于发挥其经济价值。可能原因是由于人工大面积建植草地导致土壤水分、养分、酶活性等的供应能力和变化有关[24]。程积民等[25]从土壤水分变化等方面研究表明因水分限制,在黄土高原半干旱区苜蓿适宜种植年限应为8~10年,其中第4~8年为苜蓿生长的高峰期;其他类型人工草地也有此变化,如王长庭等[22]对三江源区不同建植年代人工草地群落土壤养分分析表明,7龄和9龄草地土壤的含水量、容重、土壤中有机质、氮素和磷素在演替过程中逐渐降低。人工草地的退化是必然的,本研究中的人工草地均无相应的田间管理和施肥措施,因此,为了提高人工草地的生产力,必须采取一定的措施。
1)灭杂。这是最直接、最有效的手段。在人工草地建植前即采取适当的灭杂除草措施,以尽量减少早期杂草的入侵数量和速度。通过人工草地生态功能中对演替中期杂草入侵的分析可以看出,6龄之后的草地群落主要是种群数量的发育与优势度的变化,因此,田间管理关键期主要在播种之后的6年左右[11],需要及时进行田间管理,清除那些竞争优势比较大的物种,如附地菜、夏至草、长芒草等,减缓它们对优势种苜蓿的抑制,从而延缓苜蓿人工草地的生产期限,同时也可通过田间杂草危害性评价来及时清除杂草[26]。如果这一时期能够及时抑制和延缓杂草对苜蓿人工草地群落的侵入速度,就有利于延长人工草地的生产功能,更好地发挥其经济效益。
2)施肥。苜蓿草地在实行施肥、中耕等措施后,苜蓿盛产期可延长至10~15年[27-30]。由此可见,施肥也是比较好的措施,能明显提高和维持苜蓿生物量,在一定程度上可以延迟苜蓿种群衰退发生的时间,进而适当延长苜蓿草地盛产期年限,以更好地维持和实现苜蓿人工草地生态和生产功能的平衡和可持续利用。
3)有效利用水分。有很多学者发现制约苜蓿生产力的不仅是杂草和养分,还有水分,如程积民等[25]、李军等[31]的研究,本研究是旱地农业,水分来源缺乏,如何有效地提高水肥效应还需要进一步研究。水分条件适可区域,可以酌情考虑灌溉等措施[32],如孟季蒙和李卫军[33]对地下滴灌对苜蓿的生长发育与种子产量的影响表明地下滴灌能够起到增产效果。
4)选择适宜的耕作栽培制度。本研究是为了研究不受人为干扰自然演替下苜蓿人工草地的变化情况,故没有考虑栽培管理措施,刘沛松等[34]的研究结果显示不同草粮轮作方式可以恢复苜蓿草地土壤水分条件,韩丽娜等[35]、折凤霞等[36]、王学春等[37-38]、谢开云等[39]也研究了轮作、混播等对苜蓿草地可持续利用的影响。
[1]胡自治.人工草地在我国21世纪草业发展和环境治理中的重要意义[J].草原与草坪,2000,(1):12-15.
[2]朱显谟.黄土地区植被因素对于水土流失的影响[J].土壤学报,1960,8(2):110-121.
[3]张新时.草地的生态经济功能及其范式[J].科技导报,2000,8:3-7.
[4]Wang G H,Zhang X S.Supporting of potential forage production to the herbivore-based pastoral farming industry on the Loess Plateau[J].Acta Botanica Sinica,2003,45(10):1186-1194.
[5]Tang H P,Zhang X S.Establishment of optimized eco-productive paradigm in the farming-pastoral zone of northern China[J].Acta Botanica Sinica,2003,45(10):1166-1173.
[6]周华坤,赵兴全,赵亮,等.高山草甸垂穗披碱人工草地群落特征及稳定性研究[J].中国草地学报,2007,29(2):13-25.
[7]姜恕.关于开发中国西部地区退耕还林还草的建议[J].草地学报,2003,12(1):10-14.
[8]蔡志风,郝明德,吴振海,等.长期施肥对黄土区连作苜蓿草地植被特征的影响[J].水土保持通报,2011,31(1):46-52.
[9]牛翠娟,娄安如,孙儒泳,等.基础生态学(第2版)[M].北京:高等教育出版社,2011.
[10]余世孝.数量生态学导论[M].北京:科学技术文献出版社,1995.
[11]李裕元,邵明安,上官周平,等.黄土高原北部紫花苜蓿草地退化过程与植被演替研究[J].草业学报,2006,15(2):85-92.
[12]张耀生,赵新全.高寒牧区中华羊茅人工草地退化演替的数量特征研究[J].应用生态学报,2002,13(3):285-289.
[13]Peet R K.The measurement of species of diversity[J].Annual Review of Ecology and Systematic,1974,(5):285-307.
[14]王长庭,龙瑞军,王启基,等.不同类型高寒草地群落物种特征和均匀度的重要性[J].草业科学,2005,13(4):320-323.
[15]王鹤龄,牛俊义,郑华平,等.玛曲高寒沙化草地生态位特征及其施肥改良研究[J].草业学报,2008,12(3):12-24.
[16]周志宇,付华,陈亚明,等.阿拉善荒漠草地恢复演替过程中物种多样性与生产力的变化[J].草业学报,2003,12(1):34-40.
[17]侯扶江,南志标,肖金玉.重牧退化草地的植被、土壤及其耦合特征[J].应用生态学报,2002,13(8):915-922.
[18]Gary G.What is the observed relation between species richness and productivity[J].Ecology,2001,82(9):2381-2396.
[19]杜国祯,覃光莲,李自珍,等.高寒草甸植物群落中物种丰富度和生产力的关系研究[J].植物生态学报,2003,27(1):125-132.
[20]王长庭,龙瑞军,丁路明,等.草地生态系统中物种多样性、群落稳定性和生态系统功能的关系[J].草业科学,2005,22(6):1-7.
[21]杨利民,周广胜,李建东.松嫩平原草地群落物种多样性与生产力关系的研究[J].植物生态学报,2002,26(5):589-593.
[22]王长庭,龙瑞军,王启兰,等.三江源区不同建植年代人工草地群落演替与土壤养分变化[J].应用与环境生物学报,2009,15(6):737-744.
[23]史惠兰,王启基,景增春,等.江河源区人工草地群落特征、多样性及其稳定性分析[J].草业学报,2005,14(3):23-30.
[24]吴旭东,张晓娟,谢应忠,等.不同种植年限紫花苜蓿人工草地土壤有机碳及土壤酶活性垂直分布特征[J].草业学报,2013,22(1):245-251.
[25]程积民,程杰,高阳,等.半干旱区退耕地紫花苜蓿生长特性与土壤水分生态效应[J].草地学报,2011,19(4):565-570.
[26]林建海,王硕,路文杰,等.紫花苜蓿田间杂草危害评价[J].草业科学,2013,30(9):1412-1419.
[27]郝明德,张春霞,魏孝荣,等.黄土高原地区施肥对苜蓿生产力的影响[J].草地学报,2004,12(3):195-198.
[28]Raun W R,Johnson G V,Phillips S B.Alfalfa yield response to nitrogen applied after each cutting[J].Soil Science Society of America Journal,1999,63(5):1237-1243.
[29]马霞,王丽丽,李卫军,等.不同施氮水平下接种根瘤菌对苜蓿固氮效能及种子生产的影响[J].草业学报,2013,22(1):95-102.
[30]Hannaway D B,Shuler P E.Nitrogen fertilization in alfalfa production[J].Journal of Production Agriculture,1993,6:180-185.
[31]李军,陈兵,李小芳,等.黄土高原不同干旱类型区苜蓿草地深层土壤干燥化效应[J].生态学报,2007,27(1):75-89.
[32]王庆锁,孙东宝,侯向阳.晋北半干旱区苜蓿灌水次数与磷肥施用水平优化[J].农业工程学报,2012,28(14):112-117.
[33]孟季蒙,李卫军.地下滴灌对苜蓿的生长发育与种子产量的影响[J].草业学报,2012,21(1):291-295.
[34]刘沛松,贾志宽,李军,等.不同草粮轮作方式对退化苜蓿草地水分恢复的影响[J].农业工程学报,2010,26(2):95-103.
[35]韩丽娜,丁静,韩清芳,等.黄土高原区草粮(油)翻耕轮作的土壤水分及作物产量效应[J].农业工程学报,2012,28(24):129-137.
[36]折凤霞,郝明德,臧逸飞.黄土高原沟壑区苜蓿生产力及养分特性的研究[J].草业学报,2013,22(2):313-317.
[37]王学春,李军,方新宇,等.半干旱区草粮轮作田土壤水分恢复效应[J].农业工程学报,2011,27(1):81-87.
[38]王学春,李军,郝明德.长武旱塬草粮轮作田土壤水分可持续利用模式模拟[J].农业工程学报,2011,27(S1):257-266.
[39]谢开云,赵云,李向林,等.豆-禾混播草地种间关系研究进展[J].草业学报,2013,22(3):284-296.
