水稻产量和抽穗期性状的配合力效应分析
2013-12-24谢放鸣陈立云刘永柱
黄 明,谢放鸣,陈立云,刘永柱
(1.华南农业大学国家植物航天育种工程技术研究中心,广东广州510642;2.国际水稻研究所,菲律宾马尼拉DAPO Box 7777;3.湖南农业大学水稻研究所,湖南长沙410128)
配合力在亲本选育和配组选择中起着重要作用[1],亲本配合力决定杂种优势,只有选育出配合力高的亲本,才可能配组出具有强优势的杂交组合[2].近年来,关于两系法杂交水稻配合力和杂种优势的研究较多[3-6].在生产上,随着两系法杂交水稻的大面积推广应用,水稻两系法杂种优势利用显示出广阔的应用前景.两系法杂交稻能够实现持续稳步发展,关键在于培育出育性稳定、配合力强的实用型光温敏核不育系,而杂种优势由亲本的一般配合力和组合的特殊配合力决定.
水稻产量是重要育种目标之一,用光温敏核不育系水稻与广亲和性品种配制的两系杂交水稻组合,可以显著提高杂交水稻的产量.抽穗期也是重要性状,选育出生育期适宜的品种显得尤为重要,使之能够避开高温干旱或低温寒潮、雨水等恶性天气,保证稳产.抽穗期适宜的杂交品种选育,主要取决于亲本材料的选择.杂种一代抽穗期性状主要受亲本一般配合力的影响;在生育期方面,品种的一般配合力还可能与品种的生态类型和亲缘有关系,一般携带东南亚亲缘的籼型品种作为亚种间杂种的亲本之一,后代出现生育期延迟的可能性较小[7,8].
本研究选用国际水稻研究所(IRRI)育成的60个籼型或者具有粳稻血缘的偏籼型常规水稻品种,分别与两套不育系测交,对水稻产量性状及抽穗期性状的配合力效应进行研究,为根据配合力进行亲本的选择,以及在特定环境下进行生态类型育种等提供参考.
1 材料与方法
1.1 供试亲本材料
父本材料为60份常规水稻品种(表1),其选择过程为:根据国际水稻研究所2007年旱季、雨季以及2008年旱季的6组常规水稻产量比较试验,3组为Indica,另外3组为新株型(NPT),共688份;分别从每季产量比较试验中选出比对照极端增产(增产幅度前5名,划为“H”组)和极端减产(减产幅度前5名,划为“L”组)各5个品种,共60份.其中,Indica(A)和NPT(B)各占50%;高产和低产各占50%.5个温敏型核不育系分别为 S1(IR75589-31-27-8-33S)、S2(LP8S)、S3(LP10S)、S4(IR73827-23-26-15-7S)和 S5(IR77271-42-5-4-36S)和三系不育系25A(IR50825A).所有材料均由国际水稻所(IRRI)提供.本实验室前期的SSR(Simple Sequence Repeat)分子标记聚类分析结果表明,6个不育系被聚类在亲本之外的两个独立组,故将其分为两套:S1、S2、S3为第一套;其余为第二套.
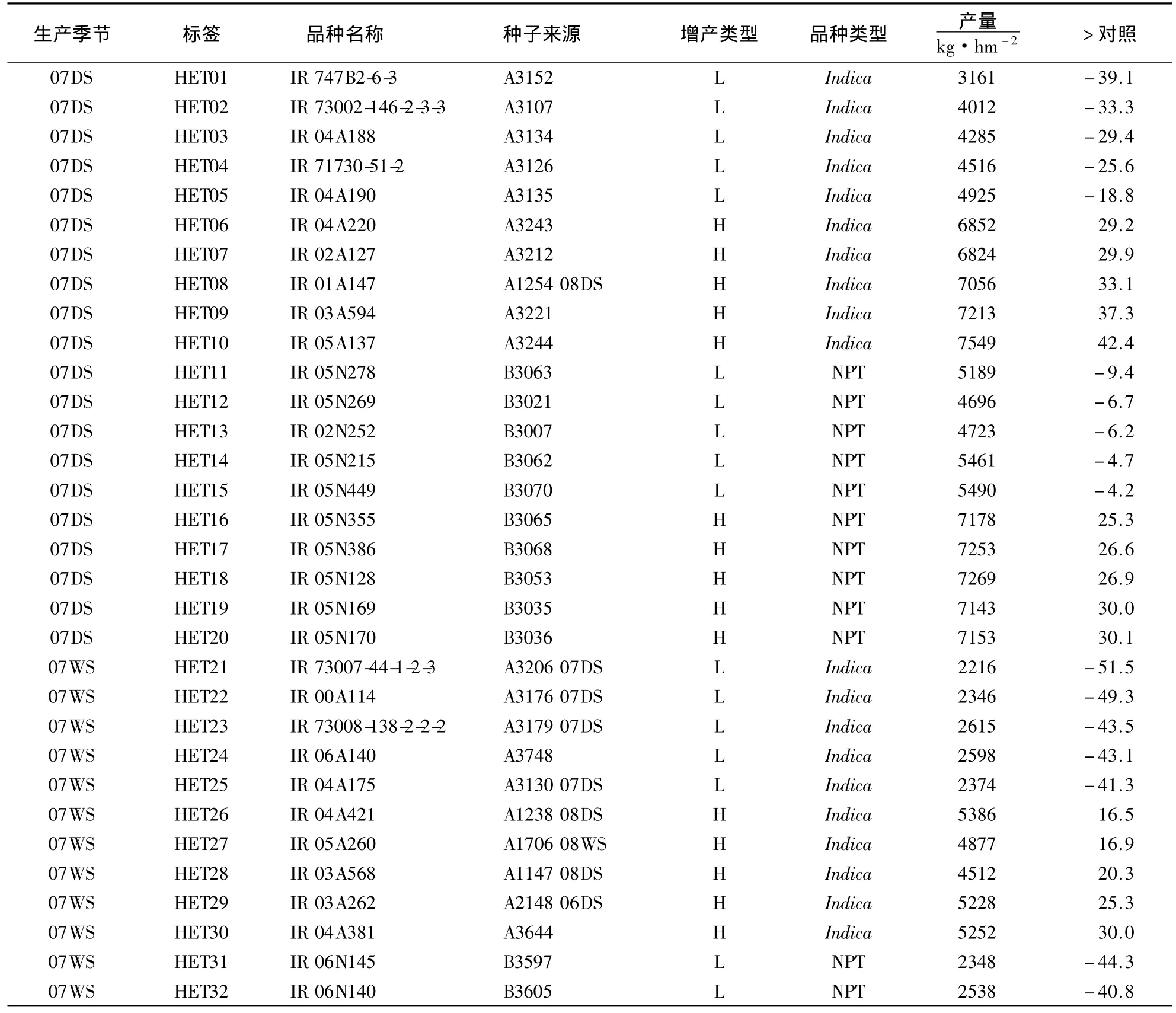
表1 60份常规水稻的信息1)Table 1 Information of the 60 accessions used in this study

续表
1.2 播种安排与田间管理
第一季(2009年旱季,09DS)安排两套测交制种.第一套为Set1 WS和Set1 DS,材料均为60个亲本与S1、S2和S3的测交;第二套为Set2 WS,即60个亲本与不育系S4、S5和25A的测交.
第二季(2009年雨季,09WS)Set1 WS的测交组合及其亲本和对照材料于2009年6月16日播种,7月6日移栽.三次重复,按随机区组实验设计(RCBD).每小区种植3行,每行12株.按照大田常规水肥管理.Set2 WS的材料及亲本和对照材料种植于另外的田块,方法同上.
第三季(2010年旱季,10DS)Set1 DS测交组合及亲本和对照于2009年11月23日播种,12月16日移栽,其试验设计、田间管理同上.
1.3 测交方法
Set1测交组合采用人工杂交方法获得杂交种子,Set2测交组合采用免隔离制种[9].
1.4 数据处理与分析
田间调查播始历期(从播种到5%抽穗的天数,DTH)和单株产量(GWP,20个单株的平均值).一般配合力和特殊配合力按照莫惠栋[10]的方法计算,利用Micro Excel 2007进行其它数据分析.
2 结果与分析
2.1 亲本材料的一般配合力
2.1.1 父本一般配合力的比较分析 (1)单季比较:分别对两套测交组合的3个随机区组试验结果进行方差分析.结果表明,60个父本(Set1 WS中没有父本HET29)在产量性状和播始历期的一般配合力之间差异极显著(P<0.01).由表 2可以看出,单株产量的一般配合力在 HET60、HET24、HET44、HET54、HET19和HET37等父本中表现比较稳定,在3个随机区组试验中,都表现为较高值或最高值;而HET04、HET05、HET32、HET35和HET31等父本都表现为较低或者最低的一般配合力负效应.播始历期一般配合力较高的亲本为HET32、HET33、HET40、HET15和HET16等,表现为对抽穗期延迟的效应,这些亲本所配组合在田间表现抽穗延迟;在亲本HET48、HET04、HET45、HET03和HET28等材料中,都为较低的负值,表现为生育期缩短,促进早熟的效应.如HET48播始历期的一般配合力分别为-7.87、-4.0和-5.96 d,具有很强的缩短生育期效应;其所配组合在田间明显表现为较早的抽穗扬花.
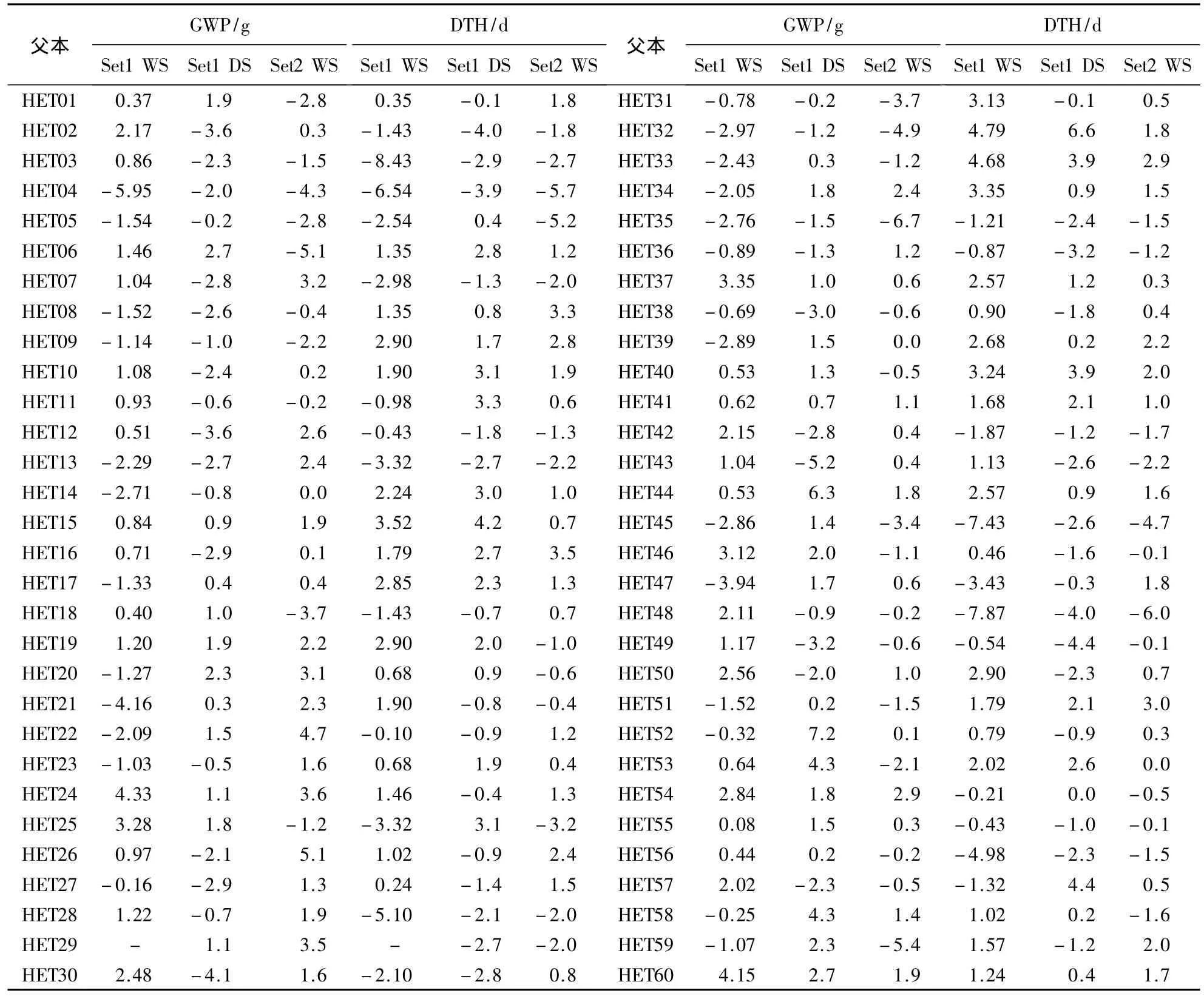
表2 父本材料在两套测交组合的3个随机区组试验中单株产量性状和播始历期的一般配合力1)Table 2 GCA for GWP and DTH of male parents in two sets of combinations with three RCBD trials
(2)父本类型比较:按照父本材料的选择标准,可以分成A(Indica)、B(NPT)两类,或者H(极端增产型)、L(极端减产型)两类.对不同父本类型的一般配合力进行方差分析.结果表明,在雨季(Set1 WS和Set2 WS),A组父本的单株产量一般配合力极显著大于B组父本(图1),且前者表现为正效应,后者表现为负效应;在2010旱季(Set1 DS),Set1表现相反.单株产量性状的一般配合力在H组和L组之间的差异与其在A组和B组之间的差异相似,且在H、L组之间差异更明显.
在雨季(WS)和旱季(DS),在A组父本中,播始历期的一般配合力均表现为负效应,而在B组父本中,均表现为正效应(图1),方差分析的结果表明,两者差异显著.H组和L组之间播始历期一般配合力的差异随季节而变,即在雨季(WS)时,H组播始历期一般配合力表现为负效应;在旱季(DS)时,H组播始历期一般配合力表现为正效应.L组则相反.
(3)季节比较:对同套测交材料Set1进行两季种植(Set1 WS,2009;Set1 DS,2010).方差分析表明(表3),父本单株产量及播始历期的一般配合力效应与环境的互作极显著,说明一般配合力效应受环境影响较大.另外,在雨季和旱季,父本产量性状一般配合力的相关系数为-0.01,未达到显著水平;而播始历期一般配合力的相关系数为0.618,达到极显著水平(P<0.01).

图1 不同类型父本在雨季(Set1 WS和Set2 WS)和旱季(Set1 DS)单株产量和播始历期一般配合力的比较Fig.1 Comparisons of GCA for GWP and DTH among different types of male parents under wet season(Set1 WS and Set2 WS)and dry season(Set1 DS)

表3 单株产量性状和播始历期的一般配合力和特殊配合力效应与环境(雨季、旱季)互作的方差分析结果Table 3 ANOVA analysis of GCA and SCA×environment(WS,DS)for GWP and DTH
在不同环境或不同测交组合中,单株产量的一般配合力效应均表现不同.播始历期的一般配合力效应与环境存在互作效应,但在不同的环境下或者不同测交系之间,播始历期的变化趋势比较接近,表现相对稳定.
2.1.2 母本一般配合力比较 在Set1测交组合中,母本S3单株产量的一般配合力最高(分别为0.545和0.905 g),在Set1 WS中,S3显著高于另2个不育系,但在Set1 DS中,差异不显著;在Set2 WS中,S4的一般配合力最高(2.305 g),三系不育系25A最低(-3.158 g).
Set1测交组合的两季数据都表明,在播始历期方面,S1的一般配合力最高,田间表现为生育期延迟,其所配组合的抽穗期一般比其他组合迟数天;S2和S3,S4和25A都表现为具有负向效应的一般配合力,能够使其杂种后代提早抽穗杨花.
Set1测交组合在WS和DS两个季节中,母本S1和S3的单株产量和播始历期的一般配合力表现比较稳定.在雨季时,S2的单株产量一般配合力为负;而在旱季时,S2的单株产量一般配合力为正,对季节的变化比较敏感.在Set 2中,只有三系不育系25A(IR50825A)的单株产量一般配合力表现为负向效应,而其他2个两系不育系均表现为正向效应,说明两系不育系在IRRI的杂交水稻育种上存在一定增产潜力和发展空间.总之,此批不育系在产量和抽穗期方面的一般配合力差异都较大,特别是播始历期的差异最大,能够为杂交水稻育种提供不同的选择,以选育出具有适当抽穗期的高产组合.
2.2 杂交组合的特殊配合力分析
在两套测交组合之间,单株产量和播始历期的特殊配合力效应均差异显著(表4).在Set1 WS、Set1 DS和 Set2 WS中,单株产量的特殊配合力变化范围分别为-7.83-6.25、-4.79-9.12 和-8.51-5.56.在Set1测交组合中,特殊配合力表现最高的组合S2/HET58(WS)和S3/HET52(DS),其对应的产量也为最高;这套测交组合在雨季和夏季种植的表现存在差异,在10个最高特殊配合力的组合中,未出现重复的组合;在两个季节中,所有组合的单株产量特殊配合力效应之间没有相关性.播始历期特殊配合力表现为相同差异.单株产量和抽穗期的特殊配合力效应受环境影响较大,互作效应极显著(表4).

表4 两套测交组合单株产量(GWP)和播始历期(DTH)特殊配合力方差分析表1)Table 4 ANOVA analysis results of SCA for GWP and DTH of the two sets of combinations
2.3 亲本一般配合力和特殊配合力效应方差
组合间方差是由父母本的一般配合力方差和特殊配合力方差组成,这2个方差分量的差异都显著,所以根据方差分析模式[11]进一步估算亲本的一般配合力及组合的特殊配合力的相对效应.结果表明,父本和母本的一般配合力差异均达到极显著水平,说明在单株产量和抽穗期方面,亲本对杂交后代有显著影响.
对亲本一般配合力相对效应值作进一步估算(表5).在Set1测交组合中,两季结果都表明,父本的单株产量一般配合力方差显著大于母本;特殊配合力方差大于亲本的一般配合力方差,尤其在雨季表现最为突出,说明测交组合的单株产量受特殊配合力的影响较大,特殊配合力的相对效应(方差)值占总基因型方差的77.88%,而一般配合力相对效应值仅为22.12%.在Set2 WS的180个测交组合中,单株产量一般配合力和特殊配合力方差的关系相反,母本表现出较强的一般配合力方差,成为影响F1代产量的主要因素,说明Set2 WS的3个不育系对后代产量的贡献较大,且大于Set1中的3个母本.

表5 亲本一般配合力方差和特殊配合力方差及其相对效应值1)Table 5 The variances of parents'GCA and SCA and its relative contribution
在雨季时,父母本的播始历期一般配合力方差之间差异不显著;在旱季时,母本表现为较强的优势;但是3组试验的结果表明,不论是旱季还是雨季,特殊配合力的相对效应值都占主导地位,且显著高于亲本一般配合力.
3 小结与讨论
本研究采用同一套IRRI常规水稻品种作为父本,分别与两套不同的不育系进行测交,其中一套分两季种植,研究了亲本材料单株产量和播始历期的一般配合力与特殊配合力效应.结果表明,单株产量和播始历期的一般配合力和特殊配合力方差达到显著或者极显著水平,说明这2个性状受加性基因和非加性基因控制.
产量性状是最为重要的一个性状,在杂交水稻育种中,常用亲本的一般配合力选择亲本材料,尤其是对父本的选择,显得比较重要.本研究表明,在两套测交系中,父母本产量性状的一般配合力方差表现不一致,在Set1测交系中,父本一般配合力方差大于母本,在另一套测交系(Set2)中,母本一般配合力方差大于父本.因此,一般配合力受亲本材料尤其是测交系(母本)的影响较大,这与翟虎渠等[6]研究一致.刘小川等[12]研究表明,母本对F1稻米品质性状的影响大于父本.抽穗期的一般配合力方差对F1的贡献大于特殊配合力方差,亲本的一般配合力对F1抽穗期的影响更为重要,其中,母本的一般配合力方差效应大于父本[7,8].以往研究主要是在一套测交或者双列杂交基础上,本研究采用不同测交系和不同季节比较.结果表明,在旱季时,母本的抽穗期一般配合力方差显著大于父本;在雨季时,表现相反;而特殊配合力的相对效应值都大于一般配合力的相对效应值.
关于亲本一般配合力方差和组合特殊配合力方差(相对效应)对杂交后代影响程度的研究,存在不同结果:前者效应大于后者[3,5]或后者效应大于前者[13];两者之间是相对对立的[4,14];两者效应基本一致[15-17];两者性质随地点而变,变异程度因性状和亲本类型而不同[18].综合本研究的2套不同测交组合(测交系不同)以及同套材料的两季种植的比较结果,产量性状受亲本一般配合力或者双亲特殊配合力的影响程度与环境(雨季和旱季)有较大关系,甚至表现出完全相反的结果.抽穗期受环境影响相对较小,但仍表现出差异.因此,有必要进行生态型水稻育种,特别是在明显旱、雨季之分的热带和亚热带地区,需要专门选育适应旱季或雨季的品种,以提高杂交水稻的产量.
[1]赵庆勇,朱镇,张亚文,等.12个粳稻新不育系的配合力及利用价值评价[J].中国水稻科学,2008,22(1):57-64.
[2]陆作楣.论杂交稻育种的配合力选择[J].中国水稻科学,1999,13(1):1-5.
[3]陈光辉,官春云,陈立云,等.两系亚种间杂交稻籽粒充实度的遗传研究[J].作物学报,2001,27(6):811-816.
[4]龚光明,周国锋,尹楚球,等.籼型两用核不育系主要农艺性状的配合力分析[J].中国水稻科学,1993,7(3):137-142.
[5]伍箴勇,李春海,牟同敏.两系杂交籼稻配合力分析[J].中国水稻科学,2007,21(6):612-618.
[6]翟虎渠,曹树青,唐运来,等.籼型杂交水稻光合性状的配合力及遗传力分析[J].作物学报,2002,28(2):154-160.
[7]袁龙江,邢祖颐.籼粳稻杂交育种的研究:Ⅲ籼粳交主要性状的配合力及遗传力[J].作物学报,1989,15(2):182-188.
[8]孙义伟,李继庆,刘义柏.两系亚种间杂交水稻抽穗日数的杂种优势和配合力分析[J].中国水稻科学,1993,7(2):105-108.
[9]VIRMANI S S,VIRAKTAMATH B C,CASAL C L,et al.Hybrid Rice Breeding Manual[M].Laguna:International Rice Research Institute,1997:89-91.
[10]莫惠栋.P×q交配模式的配合力分析[J].江苏农学院学报,1982,3(3):51-57.
[11]GRIFFING B.A generalized treatment of the use of diallel crosses in quantitative inheritance[J].Heredity,1956(10):31-50.
[12]刘小川,王渭霞,陈深广,等.杂交稻米质性状的亲本配合力分子标记鉴定[J].中国水稻科学,2005,19(1):25-28.
[13]GUIMARAES E P.Combining ability of upland rice progenitors[J].IRRN,1989,14(1):4-5.
[14]齐绍武,盛孝邦.籼型两系杂交水稻主要农艺性状配合力及遗传力分析[J].杂交水稻,2000,15(3):38-41.
[15]BOBBY T P M,NADARAJAN N.Genetic analysis of yields components in rice involving CMS lines[J].IRRN,1993,18(1):8-9.
[16]廖佩言.水稻主要性状配合力的分析[J].遗传学报,1980,2(5):22-24.
[17]王才林,汤玉庚.杂交粳稻若干米质性状的配合力[J].江苏农业学报,1989,5(1):11-17.
[18]彭俊华.杂交水稻数量性状配合力、遗传力及其地点变异性分析[J].西南农业学报,1993,6(2):12-20.
